Exploring Structure–Property Relationships of GAGs to Tailor ECM-Mimicking Hydrogels


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Charge, Structure, Ion Binding, and Morphogen Transport in GAG Hydrogels—Case Studies
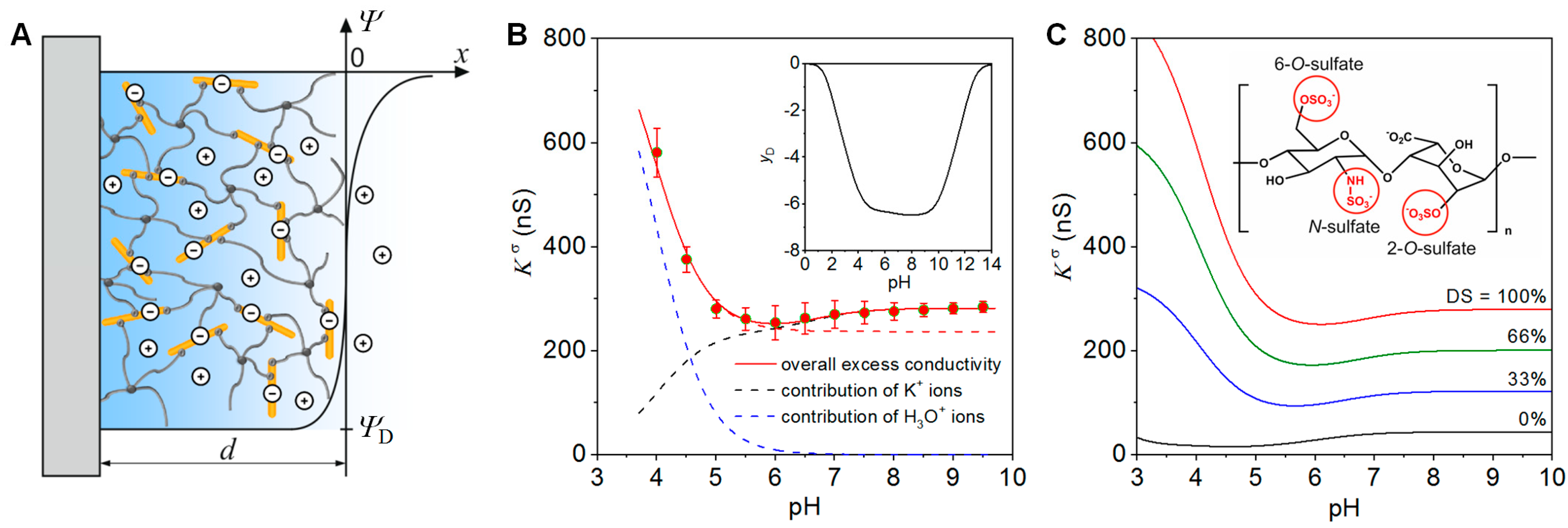
2.1. Charge and Structure of StarPEG-heparin Hydrogels
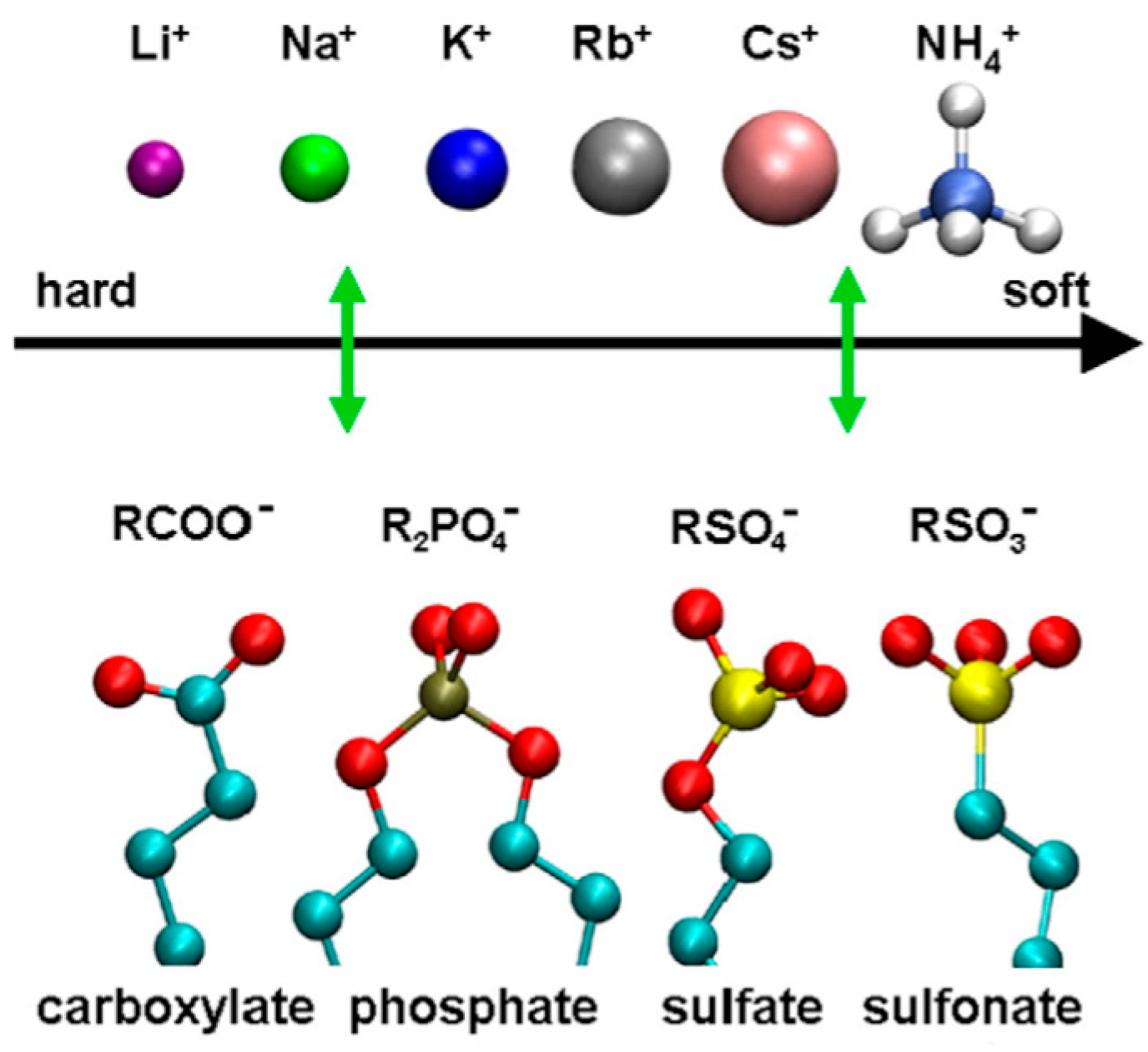
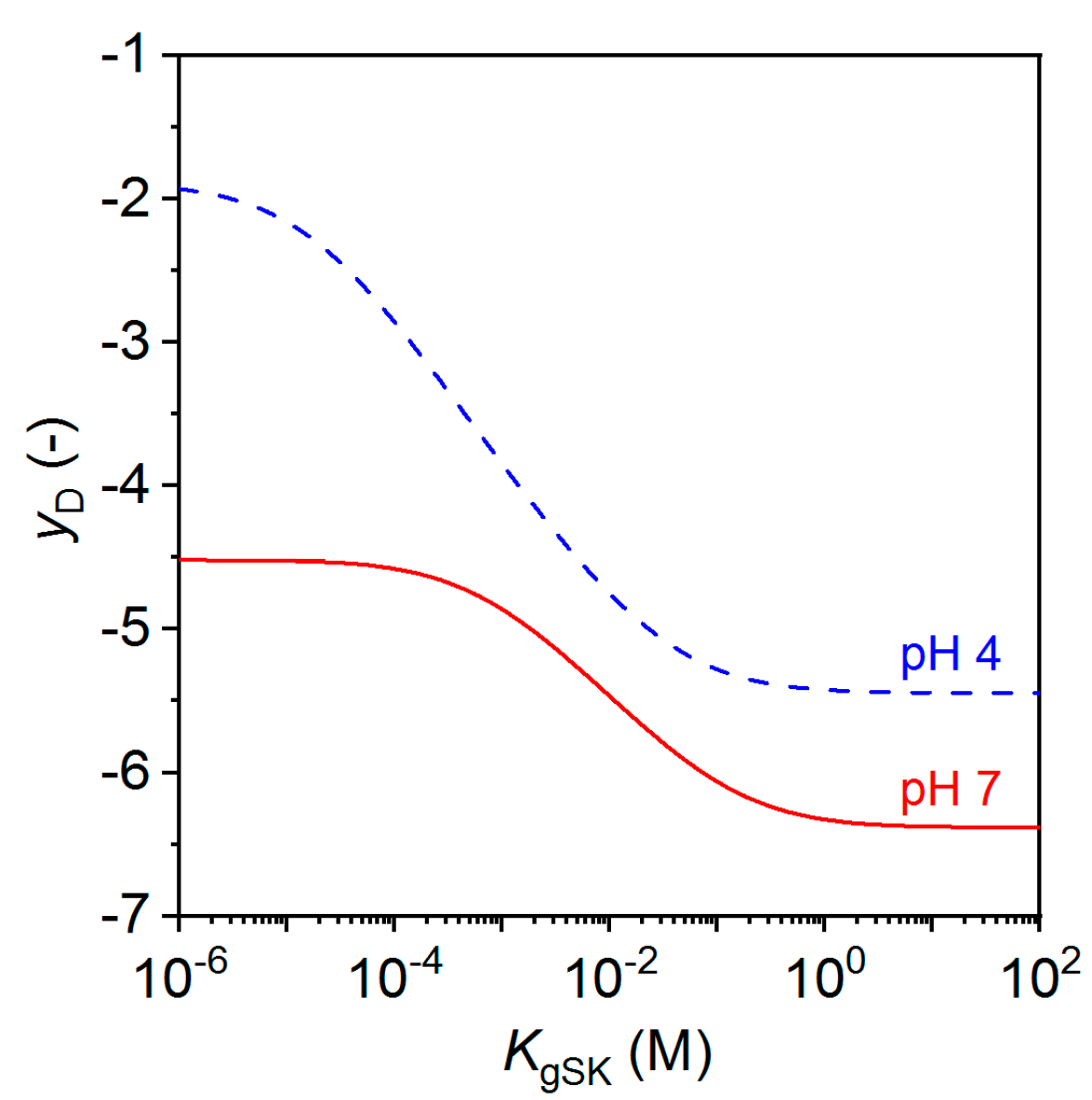
2.2. Mucosal Model with Glycan-Ion Pairing
2.3. Modulation of Morphogen Administration in StarPEG-heparin Hydrogels
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hattori, T.; Kimura, K.; Seyrek, E.; Dubin, P.L. Binding of Bovine Serum Albumin to Heparin Determined by Turbidimetric Titration and Frontal Analysis Continuous Capillary Electrophoresis. Anal. Biochem. 2001, 295, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Seyrek, E.; Dubin, P. Glycosaminoglycans as polyelectrolytes. Adv. Colloid Interface Sci. 2010, 158, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Minsky, B.B.; Atmuri, A.; Kaltashov, I.A.; Dubin, P.L. Counterion condensation on heparin oligomers. Biomacromolecules 2013, 14, 1113–1121. [Google Scholar] [CrossRef]
- Minsky, B.B.; Nguyen, T.V.; Peyton, S.R.; Kaltashov, I.A.; Dubin, P.L. Heparin Decamer Bridges a Growth Factor and an Oligolysine by Different Charge-Driven Interactions. Biomacromolecules 2013, 14, 4091–4098. [Google Scholar] [CrossRef] [PubMed]
- Comert, F.; Malanowski, A.J.; Azarikia, F.; Dubin, P.L. Coacervation and precipitation in polysaccharide–protein systems. Soft Matter 2016, 12, 4154–4161. [Google Scholar] [CrossRef]
- Ghiselle, G. Drug-Mediated Regulation of Glycosaminoglycan Biosynthesis. Med. Res. Rev. 2017, 37, 1051–1094. [Google Scholar] [CrossRef]
- Yamada, S.; Sugahara, K.; Özbek, S. Evolution of glycosaminoglycans, Comparative biochemical study. Commun. Integr. Biol. 2011, 4, 150–158. [Google Scholar] [CrossRef]
- Kamhi, E.; Joo, E.J.; Dordick, J.S.; Linhardt, R.J. Glycosaminoglycans in infectious disease. Biol. Rev. 2013, 88, 928–943. [Google Scholar] [CrossRef]
- Almond, A. Multiscale modeling of glycosaminoglycan structure and dynamics: Current methods and challenges. Curr. Opin. Struct. Biol. 2018, 50, 58–64. [Google Scholar] [CrossRef]
- Richter, R.P.; Baranova, N.S.; Day, A.J.; Kwok, J.C.F. Glycosaminoglycans in extracellular matrix organisation: Are concepts from soft matter physics key to understanding the formation of perineuronal nets? Curr. Opin. Struct. Biol. 2018, 50, 65–74. [Google Scholar] [CrossRef]
- Freudenberg, U.; Liang, Y.; Kiick, K.L.; Werner, C. Glycosaminoglycan-Based Biohybrid Hydrogels: A Sweet and Smart Choice for Multifunctional Biomaterials. Adv. Mater. 2016, 28, 8861–8891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atallah, P.; Schirmer, L.; Tsurkan, M.; Limasale, Y.D.P.; Zimmermann, R.; Werner, C.; Freudenberg, U. In situ-forming, cell-instructive hydrogels based on glycosaminoglycans with varied sulfation patterns. Biomaterials 2018, 181, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.; Goude, M.C.; McDevitt, T.C.; Temenoff, J.S. Molecular engineering of glycosaminoglycan chemistry for biomolecule delivery. Acta Biomater. 2014, 10, 1705–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belair, D.G.; Le, N.N.; Murphy, W.L. Design of Growth Factor Sequestering Biomaterials. Chem. Commun. 2014, 50, 15651–15668. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, N.; Schirmer, L.; Atallah, P.; Wandel, E.; Ferrer, R.A.; Werner, C.; Simon, J.C.; Franz, S.; Freudenberg, U. Glycosaminoglycan-based hydrogels capture chemokines and rescue wound healing deficiency. Sci. Transl. Med. 2017, 9, eaai9044. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.E.; Mooney, D.J.; Zandstra, P.W. Growth factors, matrices, and forces combine and control stem cells. Science 2009, 324, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Duval, J.F.L.; Werner, C. Probing Biointerfaces—Electrokinetics. In Biointerfaces: Where Material Meets Biology, 1st ed.; Hutmacher, D.W., Chrzanowski, W., Eds.; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 55–73. ISBN 978-1-84973-876-7. [Google Scholar]
- Barbati, A.C.; Kirby, B.J. Soft diffuse interfaces in electrokinetics—Theory and experiment for transport in charged diffuse layers. Soft Matter 2012, 8, 10598–10613. [Google Scholar] [CrossRef]
- Zimmermann, R.; Dukhin, S.S.; Werner, C.; Duval, J.F.L. On the use of electrokinetics for unraveling charging and structure of soft planar polymer films. Curr. Opin. Colloid Interface Sci. 2013, 18, 83–92. [Google Scholar] [CrossRef]
- Duboue-Dijon, E.; Delcroix, P.; Martinez-Seara, H.; Hladílkovaá, J.; Coufal, P.; Křížek, T.; Jungwirth, P. Binding of Divalent Cations to Insulin: Capillary Electrophoresis and Molecular Simulations. J. Phys. Chem. B 2018, 122, 5640–5648. [Google Scholar] [CrossRef]
- Sankaranarayanan, N.V.; Nagarajan, B.; Desai, U.R. So you think computational approaches to understanding glycosaminoglycan–protein interactions are too dry and too rigid? Think again! Curr. Opin. Struct. Biol. 2018, 50, 91–100. [Google Scholar] [CrossRef]
- Zimmermann, R.; Bartsch, S.; Freudenberg, U.; Werner, C. Electrokinetic Analysis to Reveal Composition and Structure of Biohybrid Hydrogels. Anal. Chem. 2012, 84, 9592–9595. [Google Scholar] [CrossRef] [PubMed]
- Yezek, L.P.; Duval, J.F.L.; van Leeuwen, H.P. Electrokinetics of Diffuse Soft Interfaces. III. Interpretation of Data on the Polyacrylamide/Water Interface. Langmuir 2005, 21, 6220–6227. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, U.; Hermann, A.; Welzel, P.B.; Stirl, K.; Schwarz, S.C.; Grimmer, M.; Zieris, A.; Panyanuwat, W.; Zschoche, S.; Meinhold, D.; et al. A star-PEG-heparin hydrogel platform to aid cell replacement therapies for neurodegenerative diseases. Biomaterials 2009, 30, 5049–5060. [Google Scholar] [CrossRef] [PubMed]
- Lide, D.R.; Frederikse, H.P.R. CRC Handbook of Chemistry and Physics, 78th ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 5–93. [Google Scholar]
- Freudenberg, U.; Sommer, J.-U.; Levental, K.R.; Welzel, P.B.; Zieris, A.; Chwalek, K.; Scheider, K.; Prokoph, S.; Prewitz, M.; Dockhorn, R.; et al. Using Mean Field Theory to Guide Biofunctional Materials Design. Adv. Funct. Mater. 2012, 22, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Takano, R. Desulfation of Sulfated Carbohydrates. Trends Glycosci. Glycotechnol. 2002, 14, 343–354. [Google Scholar] [CrossRef]
- Cohen, M.; Varki, A. The sialome–far more than the sum of its parts. OMICS 2010, 14, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.; Best, M.D.; Hanson, S.R.; Wong, C.-H. Sulfotransferases: Structure, Mechanism, Biological Activity, Inhibition, and Synthetic Utility. Angew. Chem. Int. Ed. 2004, 43, 3526–3548. [Google Scholar] [CrossRef] [PubMed]
- Faller, C.E.; Guvench, O. Sulfation and Cation Effects on the Conformational Properties of the Glycan Backbone of Chondroitin Sulfate Disaccharides. J. Phys. Chem. B 2015, 119, 6063–6073. [Google Scholar] [CrossRef]
- Marcus, Y.; Hefter, G. Ion Pairing. Chem. Rev. 2006, 106, 4585–4621. [Google Scholar] [CrossRef]
- Kunz, E. Specific Ion Effects; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2010; ISBN 978-981-4271-57-8. [Google Scholar]
- Vlachy, N.; Jagoda-Cwiklik, B.; Vácha, R.; Tourauda, D.; Jungwirth, P.; Kunz, W. Hofmeister series and specific interactions of charged headgroups with aqueous ions. Adv. Colloid Interface Sci. 2009, 146, 42–47. [Google Scholar] [CrossRef]
- Sakai, N.; Matile, S. Anion-Mediated Transfer of Polyarginine across Liquid and Bilayer Membranes. J. Am. Chem. Soc. 2003, 125, 14348–14356. [Google Scholar] [CrossRef]
- Fromm, J.R.; Hileman, R.E.; Caldwell, E.E.O.; Weiler, J.M.; Linhardt, R.J. Differences in the Interaction of Heparin with Arginine and Lysine and the Importance of these Basic Amino Acids in the Binding of Heparin to Acidic Fibroblast Growth Factor. Arch. Biochem. Biophys. 1995, 323, 279–287. [Google Scholar] [CrossRef]
- Schneider, C.P.; Shukla, D.; Trout, B.L. Arginine and the Hofmeister Series: The Role of Ion–Ion Interactions in Protein Aggregation Suppression. J. Phys. Chem. B 2011, 115, 7447–7458. [Google Scholar] [CrossRef] [Green Version]
- Sterling, J.D.; Baker, S.M. A Continuum Model of Mucosa with Glycan-Ion Pairing. Macromol. Theory Simul. 2018, 27, 1700079. [Google Scholar] [CrossRef]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell. Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Peysselon, F.; Ricard-Blum, S. Heparin-protein interactions: From affinity and kinetics to biological roles. Application to an interaction network regulating angiogenesis. Matrix Biol. 2014, 35, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Meneghetti, M.C.Z.; Hughes, A.J.; Rudd, T.R.; Nader, H.B.; Powell, A.K.; Yates, E.A.; Lima, M.A. Heparan sulfate and heparin interactions with proteins. J. R. Soc. Interface 2015, 12, 20150589. [Google Scholar] [CrossRef] [Green Version]
- Copeland, R.A.; Pompliano, D.L.; Meek, T.D. Drug–target residence time and its implications for lead optimization. Nat. Rev. Drug Discov. 2006, 5, 730–739. [Google Scholar] [CrossRef]
- Jecklin, M.C.; Schauer, S.; Dumelin, C.E.; Zenobi, R. Label-free determination of protein-ligand binding constants using mass spectrometry and validation using surface plasmon resonance and isothermal titration calorimetry. J. Mol. Recognit. 2009, 22, 319–329. [Google Scholar] [CrossRef]
- Murthy, B.N.; Sinha, S.; Surolia, A.; Indi, S.S.; Jayaraman, N. SPR and ITC determination of the kinetics and the thermodynamics of bivalent versus monovalent sugar ligand-lectin interactions. Glycoconj J. 2008, 25, 313–321. [Google Scholar] [CrossRef]
- Chwalek, K.; Tsurkan, M.V.; Freudenberg, U.; Werner, C. Glycosaminoglycan-based hydrogels to modulate heterocellular communication in in vitro angiogenesis models. Sci. Rep. 2014, 4, 4414. [Google Scholar] [CrossRef] [PubMed]
- Bray, L.J.; Binner, M.; Holzheu, A.; Friedrichs, J.; Freudenberg, U.; Hutmacher, D.W.; Werner, C. Multi-parametric hydrogels support 3D in vitro bioengineered microenvironment models of tumour angiogenesis. Biomaterials 2015, 53, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.M.; Tsurkan, M.V.; Magno, V.; Freudenberg, U.; Werner, C. Heparin-based hydrogels induce human renal tubulogenesis in vitro. Acta Biomater. 2017, 57, 59–69. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zimmermann, R.; Werner, C.; Sterling, J. Exploring Structure–Property Relationships of GAGs to Tailor ECM-Mimicking Hydrogels. Polymers 2018, 10, 1376. https://doi.org/10.3390/polym10121376
Zimmermann R, Werner C, Sterling J. Exploring Structure–Property Relationships of GAGs to Tailor ECM-Mimicking Hydrogels. Polymers. 2018; 10(12):1376. https://doi.org/10.3390/polym10121376
Chicago/Turabian StyleZimmermann, Ralf, Carsten Werner, and James Sterling. 2018. "Exploring Structure–Property Relationships of GAGs to Tailor ECM-Mimicking Hydrogels" Polymers 10, no. 12: 1376. https://doi.org/10.3390/polym10121376