
The Effect of DNA from Escherichia Coli at High and Low CO2 Concentrations on the Shape and Form of Crystal-line Silica-Carbonates of Barium (II)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of the Genomic DNA
2.2. Electrophoretic Analysis
2.3. Spectrophotometric Analysis
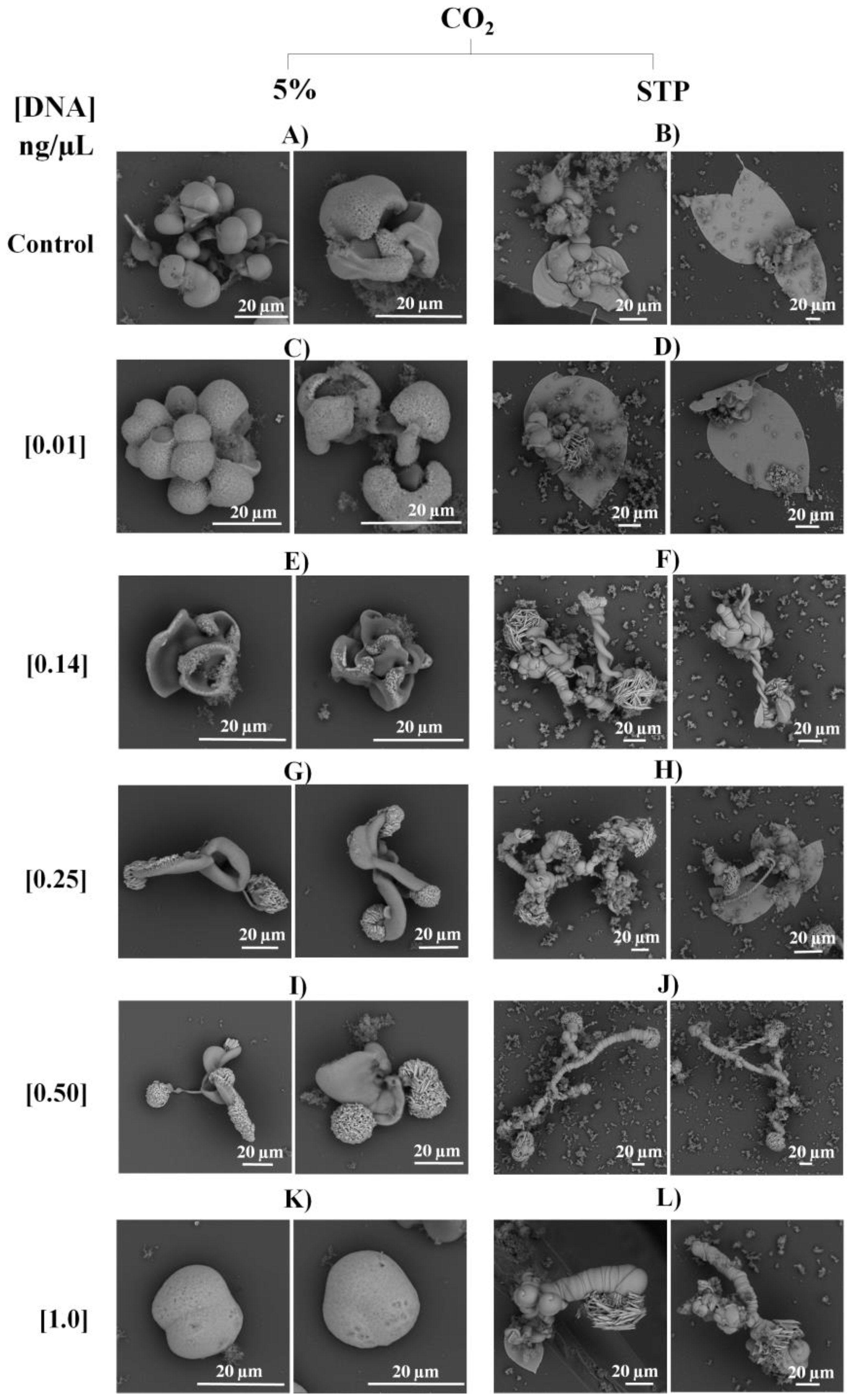
2.4. Biomorphs Formation
2.5. Characterization of Biomorphs
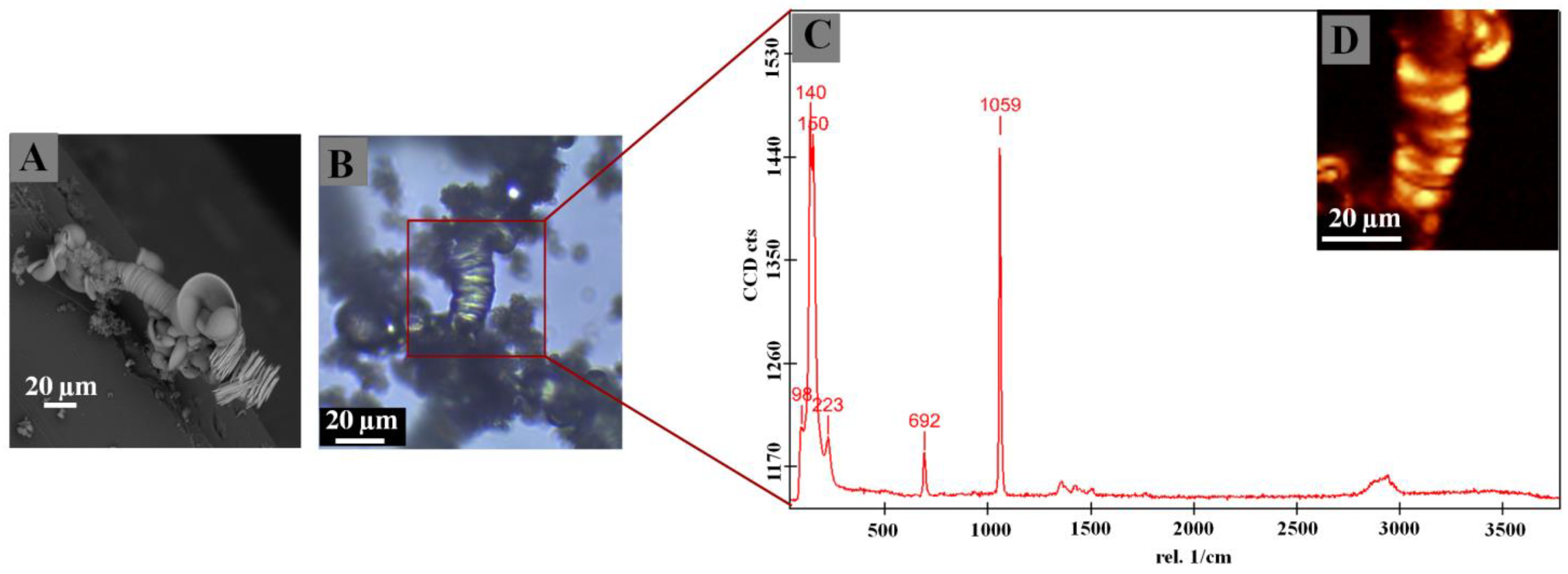
2.5.1. Scanning Electron Microscopy (SEM)
2.5.2. Raman Microspectroscopy
2.5.3. Fourier-Transform Infrared Spectroscopy (FTIR)
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oro, J. Chemical evolution and the origin of life. Adv. Space Res. 1983, 9, 77–94. [Google Scholar] [CrossRef]
- Powner, M.; Gerland, B.; Sutherland, J. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.; et al. History of the ribosome and the origin of translation. Proc. Natl. Acad. Sci. USA 2015, 112, 15396–15401. [Google Scholar] [CrossRef]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef]
- Fedor, M.J.; Williamson, J.R. The catalytic diversity of RNAs. Nat. Rev. Mol. Cell Biol. 2005, 6, 399–412. [Google Scholar] [CrossRef]
- Brunk, C.F.; Marshall, C.R. Whole Organism, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life. Life 2021, 11, 690. [Google Scholar] [CrossRef]
- Yadav, M.; Kumar, R.; Krishnamurthy, R. Chemistry of Abiotic Nucleotide Synthesis. Chem. Rev. 2020, 120, 4766–4805. [Google Scholar] [CrossRef]
- Orgel, L.E.; Lohrmann, R. Prebiotic Chemistry and Nucleic Acid Replication. Acc. Chem. Res. 1974, 7, 368–377. [Google Scholar] [CrossRef]
- Lazcano, A.; Miller, S.L. The Origin and Early Evolution of Life: Prebiotic Chemistry, the Pre-RNA World, and Time. Cell 1996, 85, 793–798. [Google Scholar] [CrossRef]
- Oro, J.; Stephen-Sherwood, E. The Prebiotic Synthesis of Oligonucleotides. In Cosmochemical Evolution and the Origins of Life: Proceedings of the Fourth International Conference on the Origin of Life and the First Meeting of the International Society for the Study of the Origin of Life, Barcelona, June 25−28, 1973, Volume I: Invited Papers and Volume II: Contributed Papers; Oró, J., Miller, S.L., Ponnamperuma, C., Young, R.S., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 1974; pp. 159–172. [Google Scholar]
- Powner, M.W.; Zheng, S.L.; Szostak, J.W. Multicomponent Assembly of Proposed DNA Precursors in Water. J. Am. Chem. Soc. 2012, 134, 13889–13895. [Google Scholar] [CrossRef]
- Follmann, H. Deoxyribonucleotides: The Unusual Chemistry and Biochemistry of DNA Precursors. Chem. Soc. Rev. 2004, 33, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Wachtershauser, G. The Place of RNA in the Origin and Early Evolution of the Genetic Machinery. Life 2014, 4, 1050–1091. [Google Scholar] [CrossRef] [PubMed]
- Dortch, Q.; Roberts, T.; Clayton, J.; Ahmed, S. RNA/DNA ratios and DNA concentrations as indicators of growth rate and biomass in planktonic marine organisms. Mar. Ecol. Prog. Ser. 1983, 13, 61–71. [Google Scholar] [CrossRef]
- Caldwell, P.C.; Hinshelwood, C. The nucleic acid content of Bact. lactis aerogenes. J. Chem. Soc. 1950, 1415–1418. [Google Scholar] [CrossRef]
- Dennis, P.P.; Bremer, H. Macromolecular composition during steady-state growth of Escherichia coli B-r. J Bacteriol. 1974, 119, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Chícharo, M.A.; Chícharo, L. RNA:DNA ratio and other nucleic acid derived indices in marine ecology. Int. J. Mol. Sci. 2008, 9, 1453–1471. [Google Scholar] [CrossRef]
- Leick, V. Ratios between contents of DNA, RNA and protein in different micro-organisms as a function of maximal growth rate. Nature 1968, 217, 1153–1155. [Google Scholar] [CrossRef]
- Mirsky, A.; Ris, H. Variable and Constant Components of Chromosomes. Nature 1949, 163, 666–667. [Google Scholar] [CrossRef]
- Wächtershäuser, G. From volcanic origins of chemoautotrophic life to Bacteria, Archaea and Eukarya. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1787–1808. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Moreno, A. Synthesis of crystalline silica-carbonate biomorphs of Ba (II) under the presence of RNA and positively and negatively charged ITO electrodes: Obtainment of graphite via bioreduction of CO2 and its implications to the chemical origin of life on primitive Earth. ACS Omega 2020, 5, 5460–5469. [Google Scholar]
- Cuéllar-Cruz, M.; Islas, S.R.; González, G.; Moreno, A. Influence of nucleic acids on the synthesis of crystalline Ca (II), Ba (II), and Sr (II) silica-carbonate biomorphs: Implications for the chemical origin of life on primitive Earth. Cryst. Growth Des. 2019, 19, 4667–4682. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Moreno, A. The role of calcium and strontium as the most dominant elements during combinations of different alkaline Earth metals in the synthesis of crystalline silica-carbonate biomorphs. Crystals. 2019, 9, 381. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Scheneider, D.K.; Stojanoff, V.; Islas, S.R.; Sánchez-Puig, N.; Arreguín-Espinosa, R.; Delgado, J.M.; Moreno, A. Formation of crystalline silica-carbonate biomorphs of alkaline Earth metals (Ca, Ba, Sr) from ambient to low temperatures: Chemical implications during the primitive Earth’s life. Cryst. Growth Des. 2020, 20, 1186–1195. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M. Influence of abiotic factors in the chemical origin of life: Biomorphs as a study model. ACS Omega 2021, 6, 8754–8763. [Google Scholar] [CrossRef]
- Zhang, G.; Morales, J.; Garcia-Ruiz, J.M. Growth behaviour of silica/carbonate nanocrystalline composites of calcite and aragonite. J. Mater. Chem. B 2017, 5, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, J.M.; Hyde, S.T.; Carnerup, A.M.; Christy, A.G.; Kranendonk, V.M.J.; Welham, N.J. Self-Assembled Silica-Carbons structures and detection of ancient microfossils. Science 2003, 302, 1194–1197. [Google Scholar] [CrossRef] [PubMed]
- Opel, J.; Wimmer, F.P.; Kellermeier, M.; Colfen, H. Functionalisation of silica-carbonate biomorphs. Nanoscale Horiz. 2016, 1, 144–149. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Islas, S.R.; Ramírez-Ramírez, N.; Pedraza-Reyes, M.; Moreno, A. Protection of the DNA from selected species of five kingdoms in Nature by Ba(II), Sr(II), and Ca(II) silica-carbonates: Implications about biogenicity and evolving from the prebiotic chemistry to biological chemistry. ACS Omega 2022. submitted at August 2022. [Google Scholar]
- Sánchez-Puig, N.; Cuéllar-Cruz, M.; Islas, S.R.; Tapia-Vieyra, J.V.; Arreguín-Espinosa, R.A.; Moreno, A. The influence of silicateins on the shape and crystalline habit of silica carbonate biomorphs of alkaline Earth metals (Ca, Ba, Sr). Cyrstals 2021, 11, 438. [Google Scholar] [CrossRef]
- Noorduin, W.L.; Grinthal, A.; Mahadevan, L.; Aiznberg, J. Rationally Designed Complex, Hierarchical Microarchitectures. Science 2013, 340, 832–837. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; Melero-García, E.; Hyde, S.T. Morphogenesis of self-assembled nanocrystalline materials of barium carbonate and silica. Science 2009, 362, 362–365. [Google Scholar] [CrossRef]
- Montalti, M.; Zhang, G.; Genovese, D.; Morales, J.; Kellermeier, M.; Garcia-Ruiz, J.M. Local pH oscillations witness autocatalytic self organization of biomorphic nanostructures. Nat. Commun. 2017, 8, 14427. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Liu, L.G. High-pressure Raman spectroscopic study of post-aragonite phase transition in witherite (BaCO3). Eur. J. Miner. 1997, 9, 785–792. [Google Scholar] [CrossRef]
- Strauss, H.; Moore, T.B. Abundances and isotopic compositions of carbon and sulfur species in whole rock and kerogen samples. In The Proterozoic Biosphere, A Multidisciplinary Study; Schopf, J.W., Klein, C., Eds.; Cambridge University Press: New York, NY, USA, 1992; pp. 709–798. [Google Scholar]
- Schopf, J.W.; Kudryavtsev, A.B. Biogenicity of Earth´s earliest fossils: A resolution of the controversy. Gondwana Res. 2012, 22, 761–771. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Wdowiak, T.J.; Czaja, A.D. Laser-Raman imagery of Earth’s earliest fossils. Nature 2002, 416, 73–76. [Google Scholar] [CrossRef]
- Marshall, W.F.; Young, K.D.; Swaffer, M.; Wood, E.; Nurse, P.; Kimura, A.; Frankel, J.; Wallingford, J.; Walbot, V.; Qu, X.; et al. What determines cell size? BMC Biol. 2012, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Ganz, H.H.; Kalkreuth, W. IR classification of kerogen type, thermal maturation, hydrocarbon potential and lithological characteristics. J. Southeast Asian Earth Sci. 1991, 5, 19–28. [Google Scholar] [CrossRef]
- Shuter, B.J.; Thomas, J.E.; Taylor, W.D.; Zimmerman, A.M. Phenotypic correlates of genomic DNA content in unicellular eukaryotes and other cells. Am. Nat. 1983, 122, 26–44. [Google Scholar] [CrossRef]
- Gunge, N.; Nakatomi, Y. Genetic mechanisms of rare matings of the yeast Saccharomyces cerevisiae heterozygous for mating type. Genetics 1972, 70, 41–58. [Google Scholar] [CrossRef]
- Pedersen, R.A. DNA content, ribosomal gene multiplicity and cell size in fish. J. Exp. Zool. 1971, 177, 65–78. [Google Scholar] [CrossRef]
- Zhang, K.; Feng, Q.L. Early Cambrian radiolarians and sponge spicules from the Niujiaohe Formation in South China. Palaeoworld 2019, 28, 234–242. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Hein, A.; Damiani, R. Nuclear DNA Content Varies with Cell Size across Human Cell Types. Cold Spring Harb. Perspect. Biol. 2015, 7, a019091. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gong, F.C.; Lambert, G.M.; Galbraith, D. Cell type-specific characterization of nuclear DNA contents within complex tissues and organs. Plant Methods 2005, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Rauwolf, U.; Golczyk, H.; Greiner, S.; Herrmann, R.G. Variable amounts of DNA related to the size of chloroplasts III. Biochemical determinations of DNA amounts per organelle. Mol. Genet. Genom. 2010, 283, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Jovtchev, G.; Schubert, V.; Meister, A.; Barow, M.; Schubert, I. Nuclear DNA content and nuclear and cell volume are positively correlated in angiosperms. Cytogenet. Genome Res. 2006, 114, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, B.; Stewart, J.M. Estimation of the nuclear DNA content of gossypium species. Ann. Bot. 2005, 95, 789–797. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.D.; Lewis-Smith, R.I. DNA amounts of angiosperms from the Antartic and South Georgia. Environ. Exp. Bot. 1982, 22, 307–318. [Google Scholar] [CrossRef]
- Caceres, M.E.; Pace, C.D.; Mugnozza, G.T.S.; Kotsonis, P.; Ceccarelli, M.; Cionini, P.G. Genomes size variations within Dasypyrum villosum: Correlations with chromosomal traits, environmental factors and plant phenotypic characteristics and behavior in reproduction. Theoret. Appl. Genet. 1998, 96, 559–567. [Google Scholar] [CrossRef]
- Rayburn, A.L. Genome size variation in Southwestern United States Indian maize adapted to various altitudes. Evol. Trends Plants 1990, 4, 53–57. [Google Scholar]
- Wakamiya, I.; Newton, R.J.; Johnston, S.J.; Price, J.H. Genome size and environmental factors in the genus Pinus. Am. J. Bot. 1993, 80, 1235–1241. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample/DNA Concentration [ng/µL] | Synthesis Condition | Raman (cm−1) | IR (cm−1) | Composition |
|---|---|---|---|---|
| Control (-) | 5% CO2 | 93, 138, 223, 690, 1058 | 629, 787, 855, 937, 1059, 1417, 1732 | Witherite /abiotic |
| STP | 140, 698, 1059, 2893 | 692, 796, 855, 958, 1099, 1417 | Witherite/abiotic | |
| [0.01] | 5% CO2 | 106, 147, 151, 700, 1067, 2915 | 692, 789, 855, 1059, 1417, 1999 | Witherite/abiotic |
| STP | 93, 138, 691, 1058 | 692, 787, 796, 855, 946, 1050, 1072, 1416, 1577 | Witherite/abiotic | |
| [0.14] | 5% CO2 | 112, 499, 1456, 2918 | 649, 787, 937, 1417, 2000 | Witherite/abiotic |
| STP | 141, 155, 693, 1061 | 592, 692, 767, 899, 1069, 1417, 1577, 2851 | Witherite/abiotic | |
| [0.25] | 5% CO2 | 93, 138, 155, 222, 690, 1058, 2908 | 582, 692, 789, 855, 1059, 1415, 1732, 1969 | Witherite/abiotic |
| STP | 141, 152, 223, 494693, 1061, 2909, 2967 | 603, 796, 882, 997, 1075, 1415, 1590, 1749, 1974 | Witherite/abiotic | |
| [0.50] | 5% CO2 | 99, 485, 1280, 1460, 2411, 2910, 2970 | 693, 787, 856, 891, 937, 1057, 1423, 1732, 1970, 2400, 2850, 2940 | Witherite/biotic |
| STP | 96, 156, 692, 1060, 1360, 1415, 2917 | 583, 692, 788, 796, 854, 947, 1071, 1416, 1560, 1770, 1979, 2160, 2480, 2820 | Witherite/biotic | |
| [1.00] | 5% CO2 | 100, 498, 893, 1451, 1600, 2911, 2962 | 629, 761, 787, 891, 1033, 1417, 1732, 1969, 2860, 2929 | Witherite/biotic |
| STP | 96, 150, 224, 691, 1059, 1362, 1421, 2942 | 608, 692, 796, 854, 968, 999, 1413, 1780, 2470, 2880, 2920 | Witherite/biotic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Aguilar, C.D.; Islas, S.R.; Moreno, A.; Cuéllar-Cruz, M. The Effect of DNA from Escherichia Coli at High and Low CO2 Concentrations on the Shape and Form of Crystal-line Silica-Carbonates of Barium (II). Crystals 2022, 12, 1147. https://doi.org/10.3390/cryst12081147
Pérez-Aguilar CD, Islas SR, Moreno A, Cuéllar-Cruz M. The Effect of DNA from Escherichia Coli at High and Low CO2 Concentrations on the Shape and Form of Crystal-line Silica-Carbonates of Barium (II). Crystals. 2022; 12(8):1147. https://doi.org/10.3390/cryst12081147
Chicago/Turabian StylePérez-Aguilar, Cesia D., Selene R. Islas, Abel Moreno, and Mayra Cuéllar-Cruz. 2022. "The Effect of DNA from Escherichia Coli at High and Low CO2 Concentrations on the Shape and Form of Crystal-line Silica-Carbonates of Barium (II)" Crystals 12, no. 8: 1147. https://doi.org/10.3390/cryst12081147