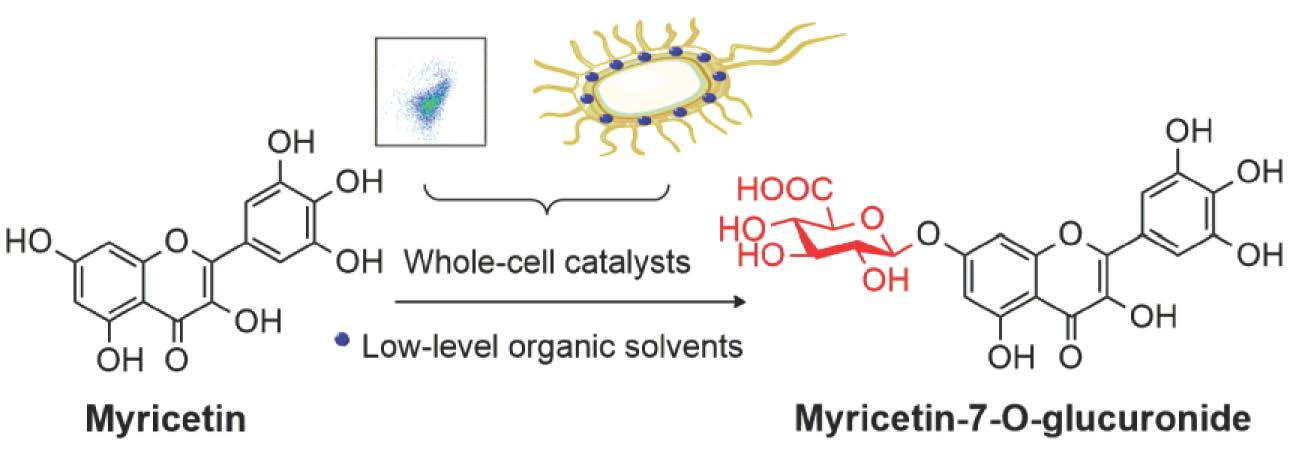
Low-Level Organic Solvents Improve Multienzyme Whole-Cell Catalytic Synthesis of Myricetin-7-O-Glucuronide

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
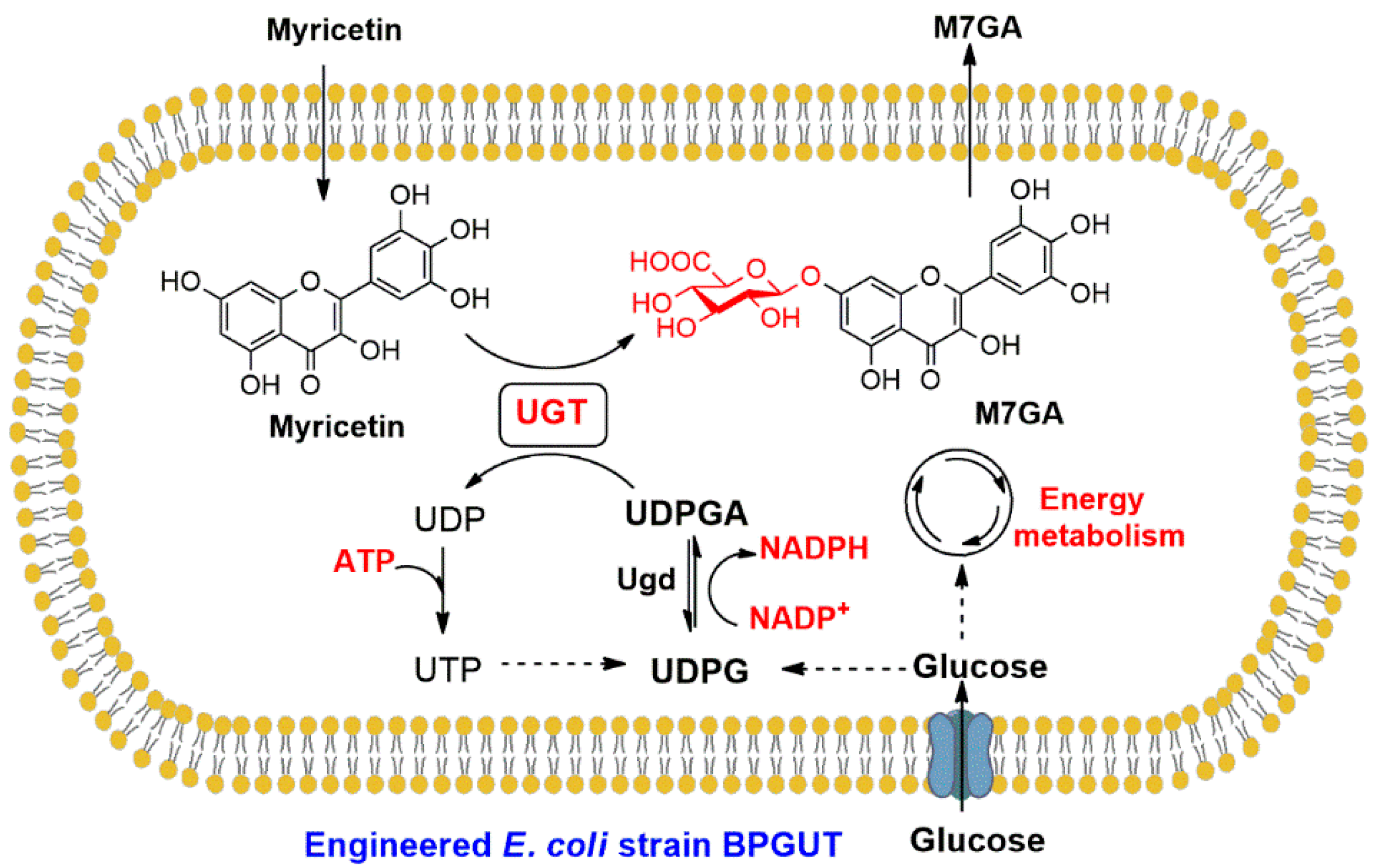
1. Introduction
2. Results and Discussion
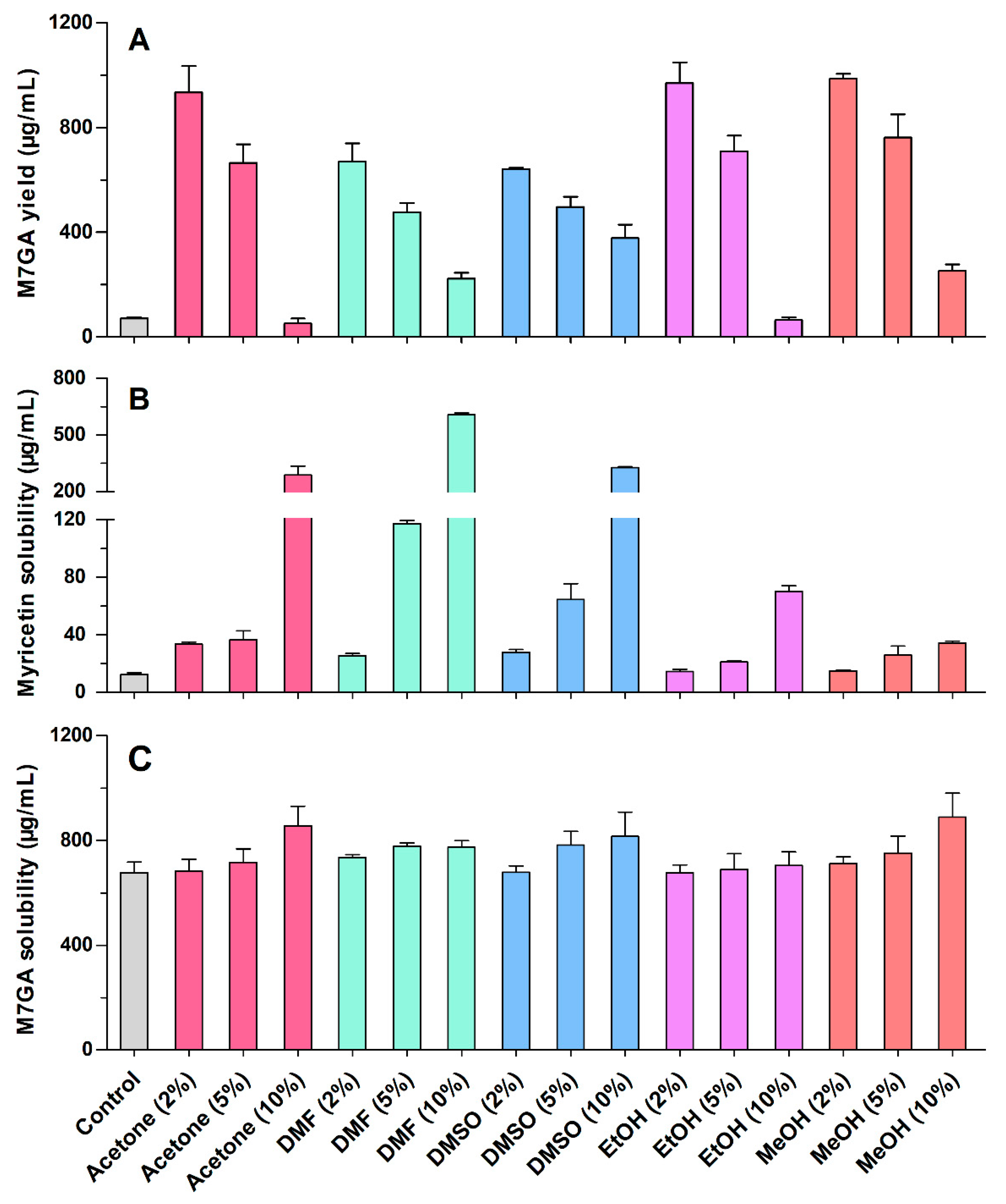
2.1. Effects of the Organic Solvents on Bioconversion
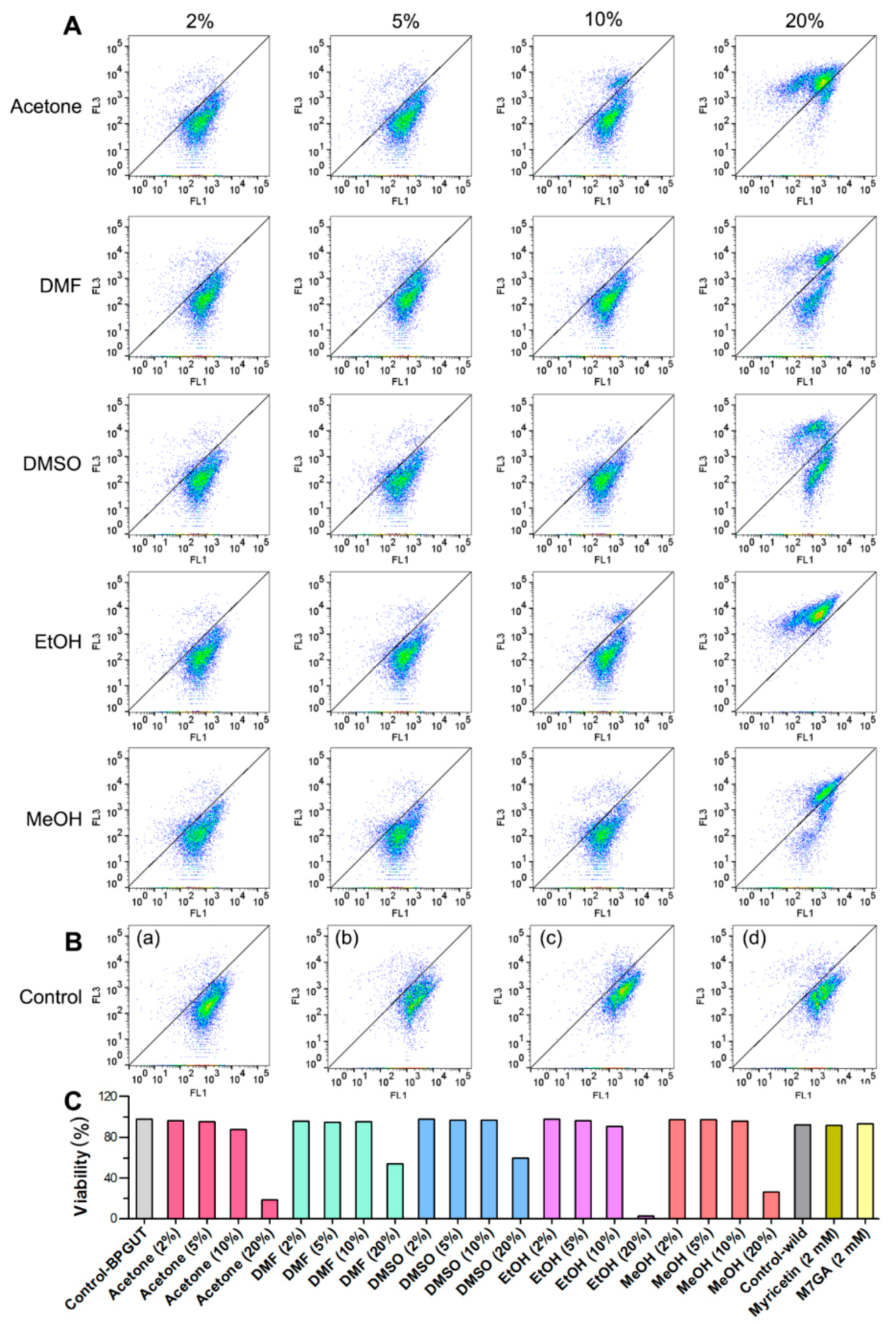
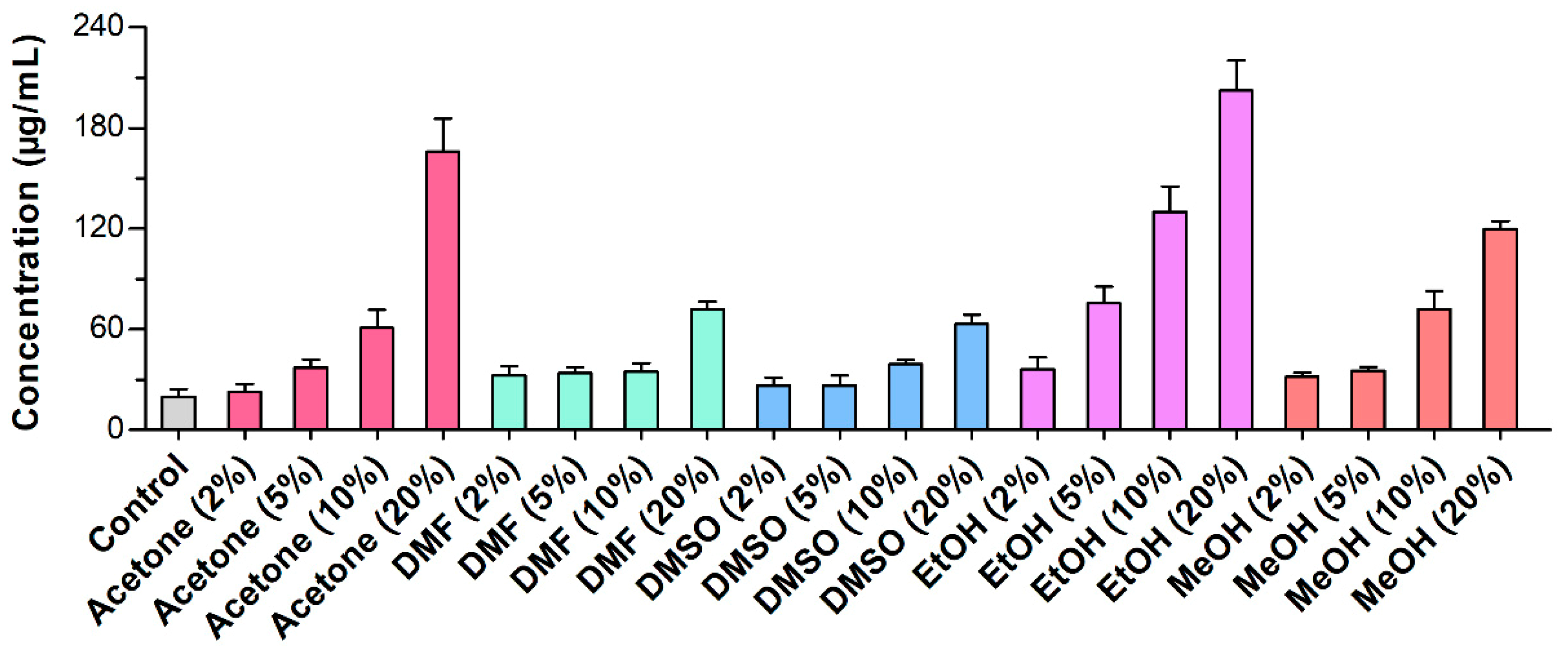
2.2. Cell Viability and Its Impacts on Bioconversion
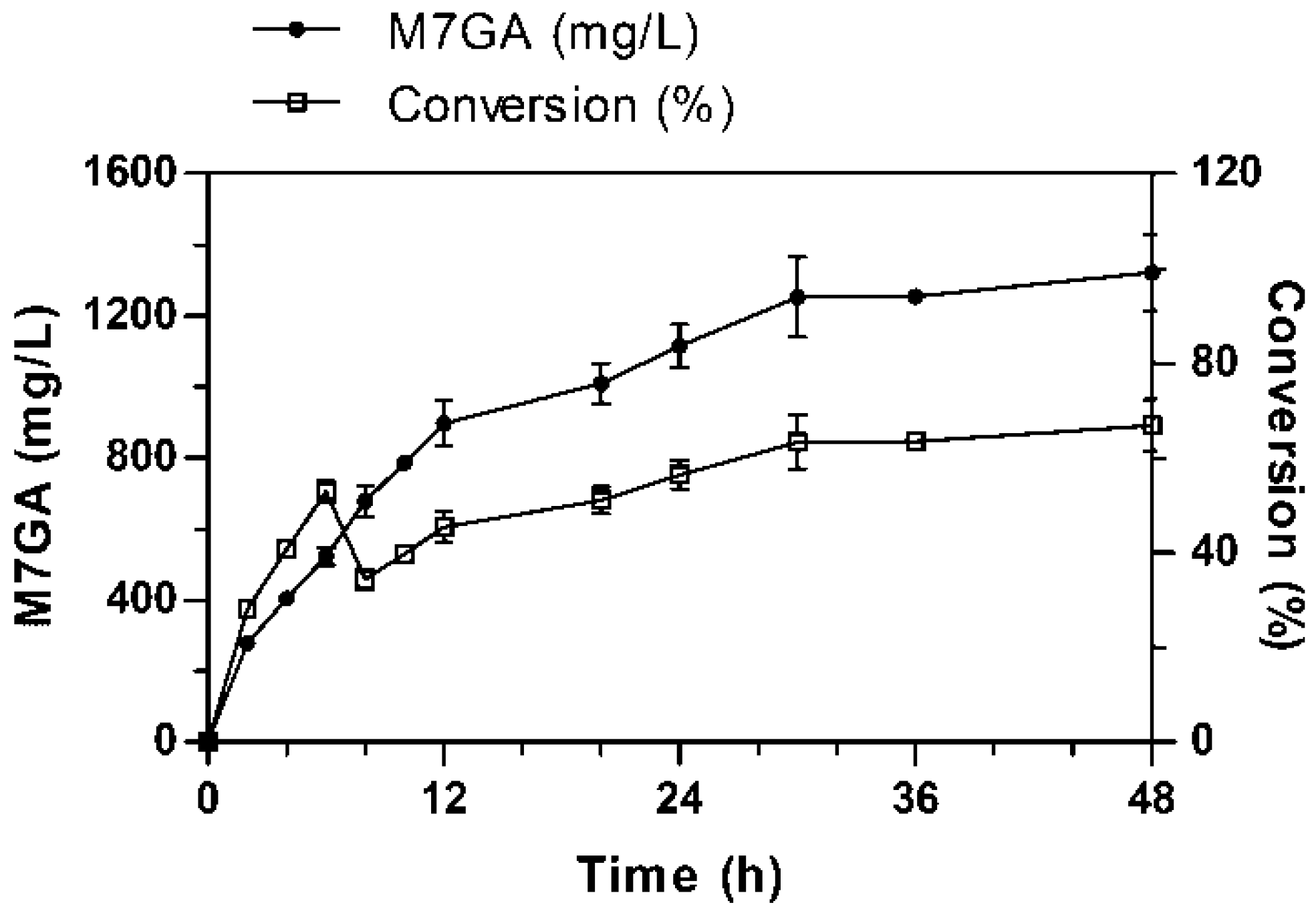
2.3. Scaled-Up Synthesis of M7GA in a Fermenter System
3. Materials and Methods
3.1. Materials
3.2. Preparation of a Certain Concentration of Organic Solvent
3.3. General Procedure for the Whole-Cell Biocatalytic Conversion of Myricetin into M7GA
3.4. Determination of Myricetin and M7GA Solubility
3.5. HPLC Analysis
3.6. Flow Cytometry (FCM) Tests
3.7. Extracellular Protein Leakage Assay
3.8. Scale-Up Synthesis of M7GA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DMF | N,N-dimethylformamide |
| DMSO | Dimethyl sulfoxide |
| EtOH | Ethanol |
| FCM | Flow cytometry |
| MeOH | Methanol |
| M7GA | Myricetin-7-O-glucuronide |
| PI | Propidium iodide |
References
- Lin, B.; Tao, Y. Whole-cell biocatalysts by design. Microb. Cell Fact. 2017, 16, 106. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Li, Z. Whole-cell cascade biotransformations for one-pot multistep organic synthesis. ChemCatChem 2018, 10, 2164–2178. [Google Scholar] [CrossRef]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-enzymatic cascade reactions: Overview and perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Shi, J.; Wu, Y.; Zhang, S.; Tian, Y.; Yang, D.; Jiang, Z. Bioinspired construction of multi-enzyme catalytic systems. Chem. Soc. Rev. 2018, 47, 4295–4313. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, C.C. Whole cell biocatalysts: Essential workers from nature to the industry. Microb. Biotechnol. 2017, 10, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Kadisch, M.; Willrodt, C.; Hillen, M.; Bühler, B.; Schmid, A. Maximizing the stability of metabolic engineering-derived whole-cell biocatalysts. Biotechnol. J. 2017, 12, 1600170. [Google Scholar] [CrossRef] [PubMed]
- Wachetmeister, J.; Rother, D. Recent advances in whole cell boicatalsysis. Curr. Opin. Biotech. 2016, 42, 169–177. [Google Scholar] [CrossRef]
- Rudroff, F. Whole-cell based synthetic enzyme cascades-light and shadow of a promising technology. Curr. Opin. Chem. Biol. 2019, 49, 84–90. [Google Scholar] [CrossRef]
- McAuliffe, J.C. Industrial enzymes and biocatalysis. In Handbook of Industrial Chemistry and Biotechnology; Kent, J.A., Ed.; Springer: Boston, MA, USA, 2012; pp. 1183–1227. ISBN 978-1-4614-4259-2. [Google Scholar]
- Krauser, S.; Weyler, C.; Blaß, L.K.; Heinzle, E. Directed multistep biocatalysis using tailored permeabilized cells. Adv. Biochem. Eng. Biotechnol. 2013, 137, 185–234. [Google Scholar] [CrossRef]
- Zheng, G.W.; Pan, J.; Yu, H.L.; Ngo-Thi, M.T.; Li, C.X.; Xu, J.H. An efficient bioprocess for enzymatic production of L-menthol with high ratio of substrate to catalyst using whole cells of recombinant E. coli. J. Biotechnol. 2010, 150, 108–114. [Google Scholar] [CrossRef]
- Li, X.F.; Yuan, T.; Xu, H.; Xin, X.; Zhao, G.; Wu, H.; Xiao, X. Whole-cell catalytic synthesis of puerarin monoesters and analysis of their antioxidant activities. J. Agric. Food Chem. 2019, 67, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Anwar, J. Modulating the structure and properties of cell membranes: The molecular mechanism of action of dimethyl sulfoxide. J. Phys. Chem. B 2007, 111, 10453–10460. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Seo, M.J.; Shin, K.C.; Oh, D.K. Production of 10S-hydroxy-8(E)-octadecenoic acid from oleic acid by whole recombinant Escherichia coli cells expressing 10S-dioxygenase from Nostoc punctiforme PCC 73102 with the aid of a chaperone. Biotechnol. Lett. 2017, 39, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Du, L.; Chen, H.; Sun, Y.; Huang, S.; Zheng, X.; Kim, E.S.; Li, S. Reconstitution of the in vitro activity of the cyclosporine-specific P450 hydroxylase from Sebekia benihana and development of a heterologous whole-cell biotransformation system. Appl. Environ. Microbiol. 2015, 81, 6268–6275. [Google Scholar] [CrossRef]
- Cui, J.D.; Jia, S.R.; Sun, A.Y. Influence of amino acids, organic solvents and surfactants for phenylalanine ammonia lyase activity in recombinant Escherichia coli. Lett. Appl. Microbiol. 2008, 46, 631–635. [Google Scholar] [CrossRef]
- Janardhan Garikipati, S.V.; Peeples, T.L. Solvent resistance pumps of Pseudomonas putida S12: Applications in 1-naphthol production and biocatalyst engineering. J. Biotechnol. 2015, 210, 91–99. [Google Scholar] [CrossRef]
- Honda, K.; Inoue, M.; Ono, T.; Okano, K.; Dekishima, Y.; Kawabata, H. Improvement of operational stability of Ogataea minuta carbonyl reductase for chiral alcohol production. J. Biosci. Bioeng. 2017, 123, 673–678. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, H.M.; Tong, Y.F.; Liu, M.Z.; Cheng, K.D.; Wu, S.; Wang, W. Systems metabolic engineering of Escherichia coli to enhance the production of flavonoid glucuronides. RSC Adv. 2016, 6, 33622–33630. [Google Scholar] [CrossRef]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A dietary molecule with diverse biological activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef]
- Guo, C.; Zhu, Y.; Weng, Y.; Wang, S.; Guan, Y.; Wei, G.; Yin, Y.; Xi, M.; Wen, A. Therapeutic time window and underlying therapeutic mechanism of breviscapine injection against cerebral ischemia/reperfusion injury in rats. J. Ethnopharmacol. 2014, 151, 660–666. [Google Scholar] [CrossRef]
- Ishisaka, A.; Kawabata, K.; Miki, S.; Shiba, Y.; Minekawa, S.; Nishikawa, T.; Mukai, R.; Terao, J.; Kawai, Y. Mitochondrial dysfunction leads to deconjugation of quercetin glucuronides in inflammatory macrophages. PLoS ONE 2013, 8, e80843. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P.; Abd-el-Mohsen, M.M.; Rice-Evans, C. Cellular uptake and metabolism of flavonoids and their metabolites: Implications for their bioactivity. Arch. Biochem. Biophys. 2004, 423, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Barron, D.; Orfila, C.; Dionisi, F.; Krajcsi, P.; Williamson, G. Interaction of hydroxycinnamic acids and their conjugates with organic anion transporters and ATP-binding cassette transporters. Mol. Nutr. Food Res. 2011, 55, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Notman, R.; Noro, M.; O´Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, N.R.; Papoutsakis, E.T. Engineering membrane and cell-wall programs for tolerance to toxic chemicals: Beyond solo genes. Curr. Opin. Microbiol. 2016, 33, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Marquês, J.T.; Viana, A.S.; De Almeida, R.F. Ethanol effects on binary and ternary supported lipid bilayers with gel/fluid domains and lipid rafts. Biochim. Biophys. Acta 2011, 1808, 405–414. [Google Scholar] [CrossRef]
- Weber, F.J.; de Bont, J.A. Adaptation mechanisms of microorganisms to the toxic effects of organic solvents on membranes. Biochim. Biophys. Acta 1996, 1286, 225–245. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Liu, M.-Z.; Cao, Y.-S.; Li, C.-K.; Wang, W. Low-Level Organic Solvents Improve Multienzyme Whole-Cell Catalytic Synthesis of Myricetin-7-O-Glucuronide. Catalysts 2019, 9, 970. https://doi.org/10.3390/catal9110970
Yang Y, Liu M-Z, Cao Y-S, Li C-K, Wang W. Low-Level Organic Solvents Improve Multienzyme Whole-Cell Catalytic Synthesis of Myricetin-7-O-Glucuronide. Catalysts. 2019; 9(11):970. https://doi.org/10.3390/catal9110970
Chicago/Turabian StyleYang, Yan, Min-Zhi Liu, Yun-Song Cao, Chang-Kun Li, and Wei Wang. 2019. "Low-Level Organic Solvents Improve Multienzyme Whole-Cell Catalytic Synthesis of Myricetin-7-O-Glucuronide" Catalysts 9, no. 11: 970. https://doi.org/10.3390/catal9110970