Irrigation Combined with Aeration Promoted Soil Respiration through Increasing Soil Microbes, Enzymes, and Crop Growth in Tomato Fields

Abstract
:1. Introduction
2. Results and Discussion
2.1. Environmental Variables
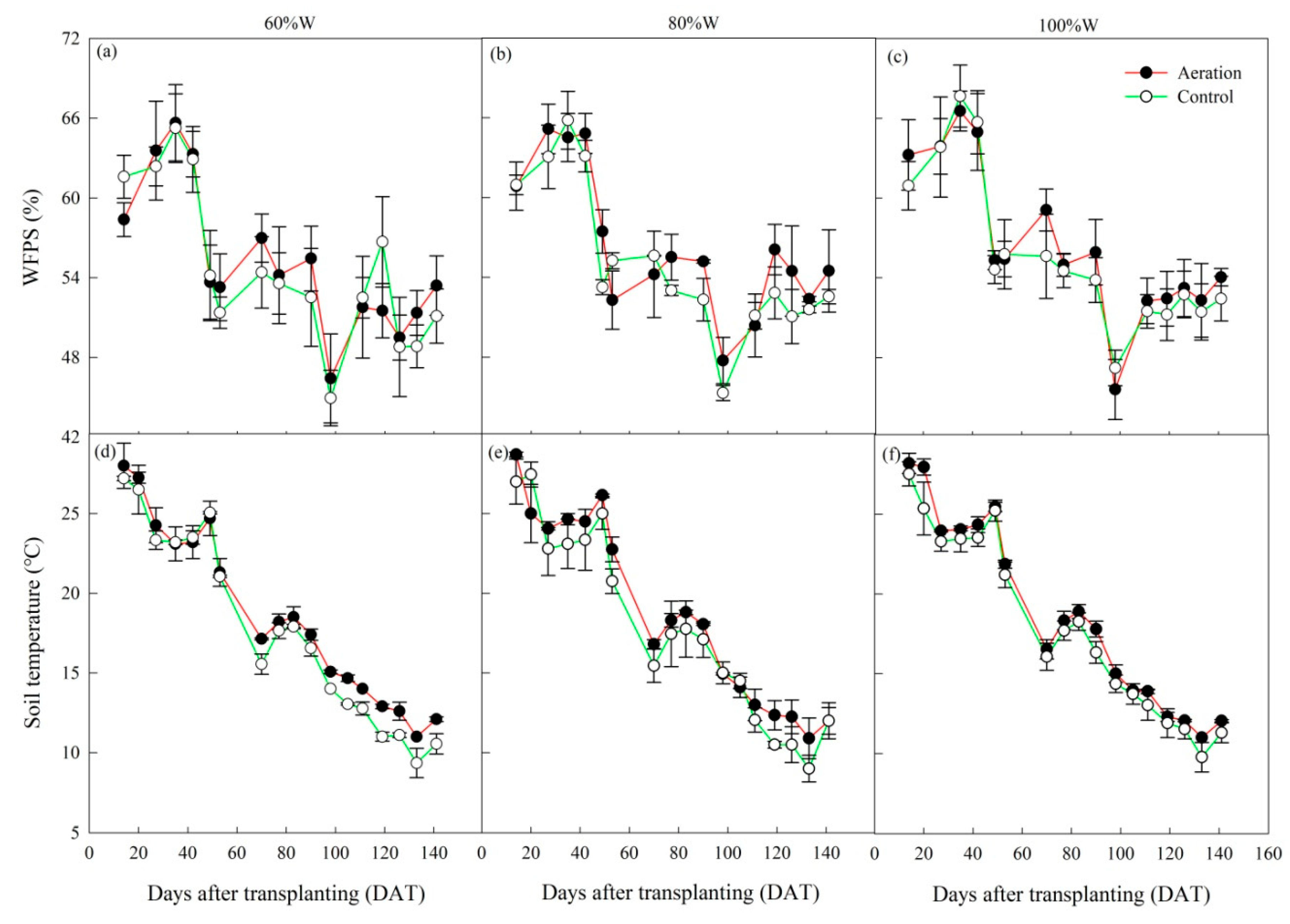
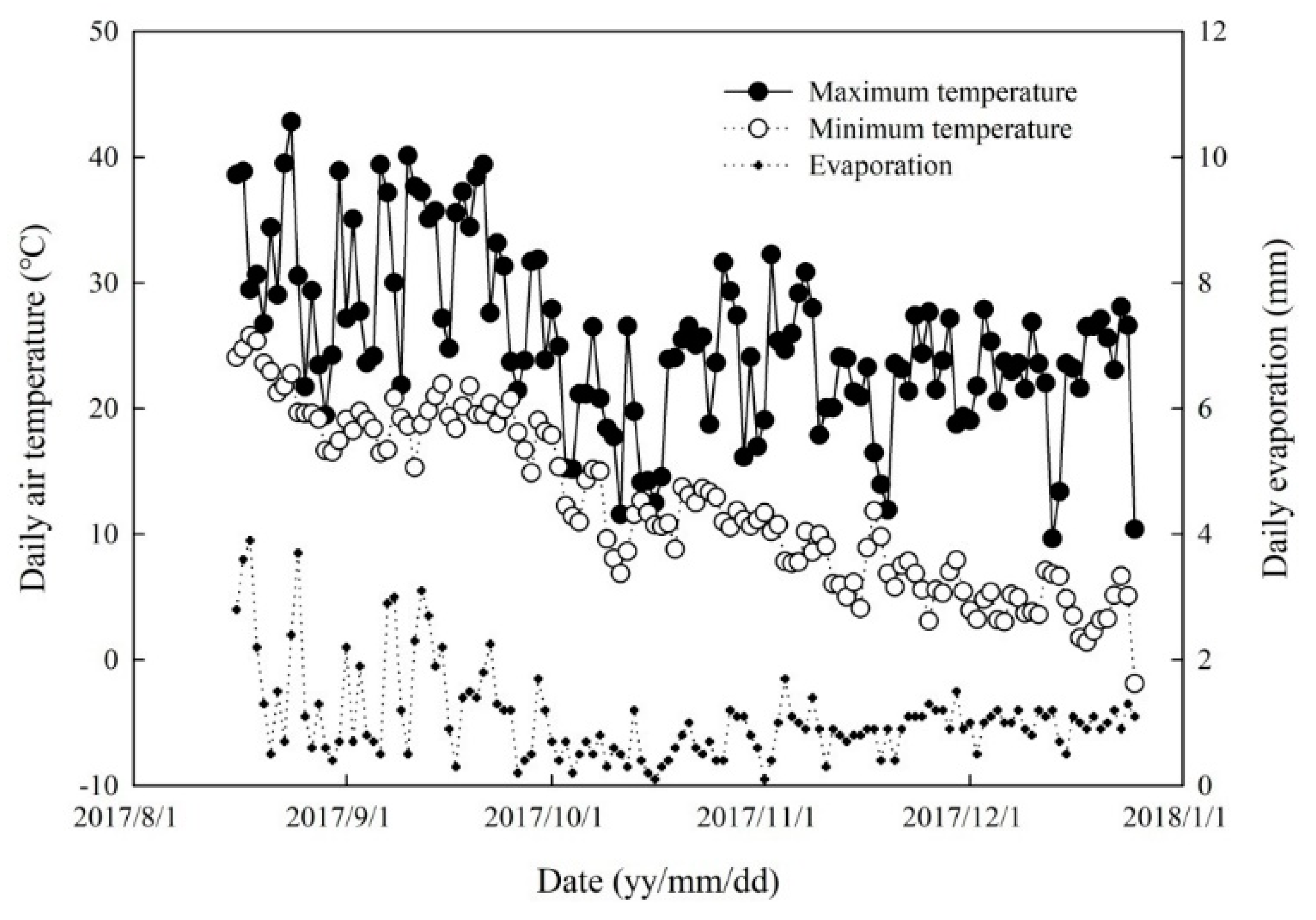
2.1.1. Soil Water-Filled Pore Space and Temperature
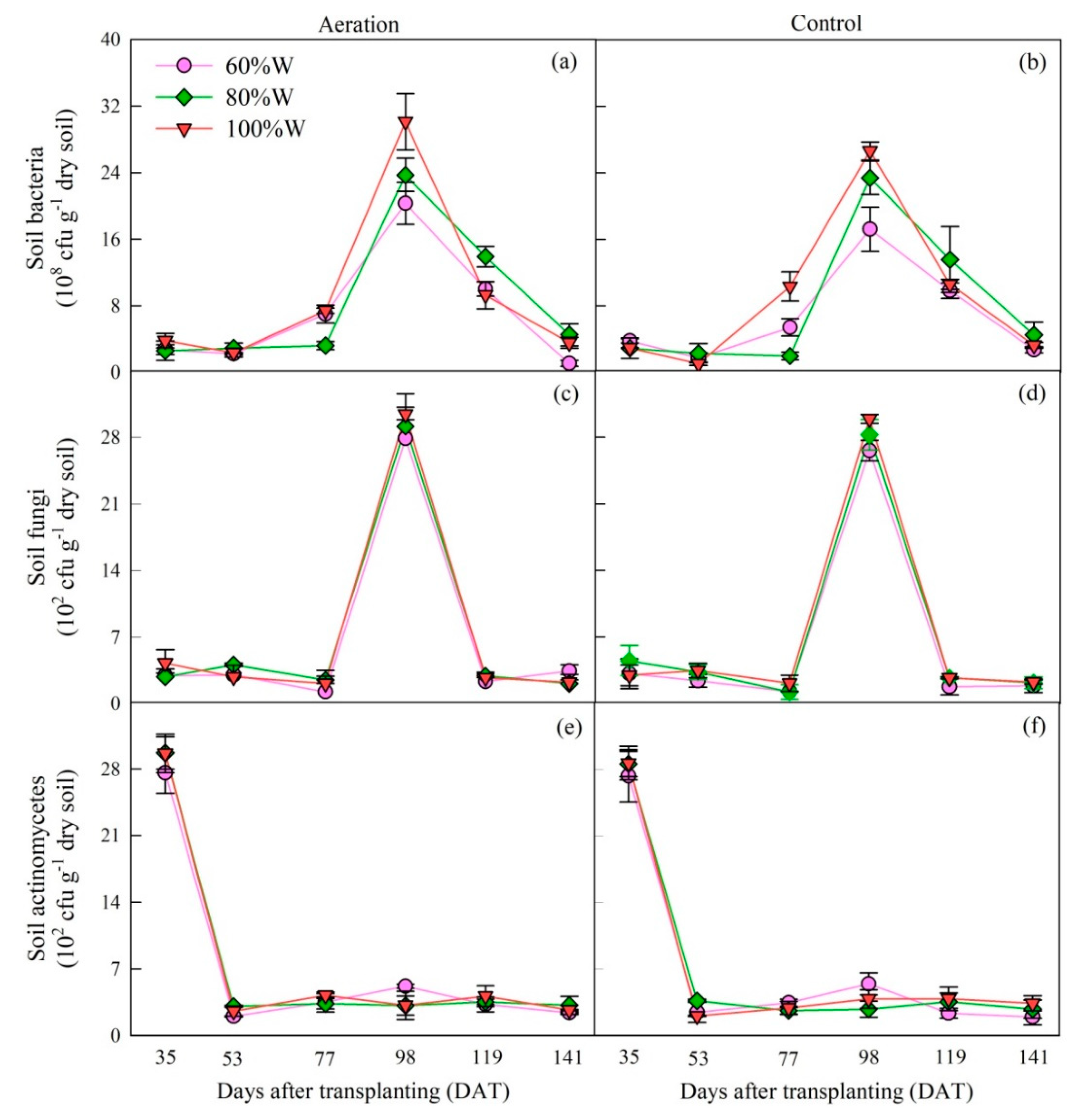
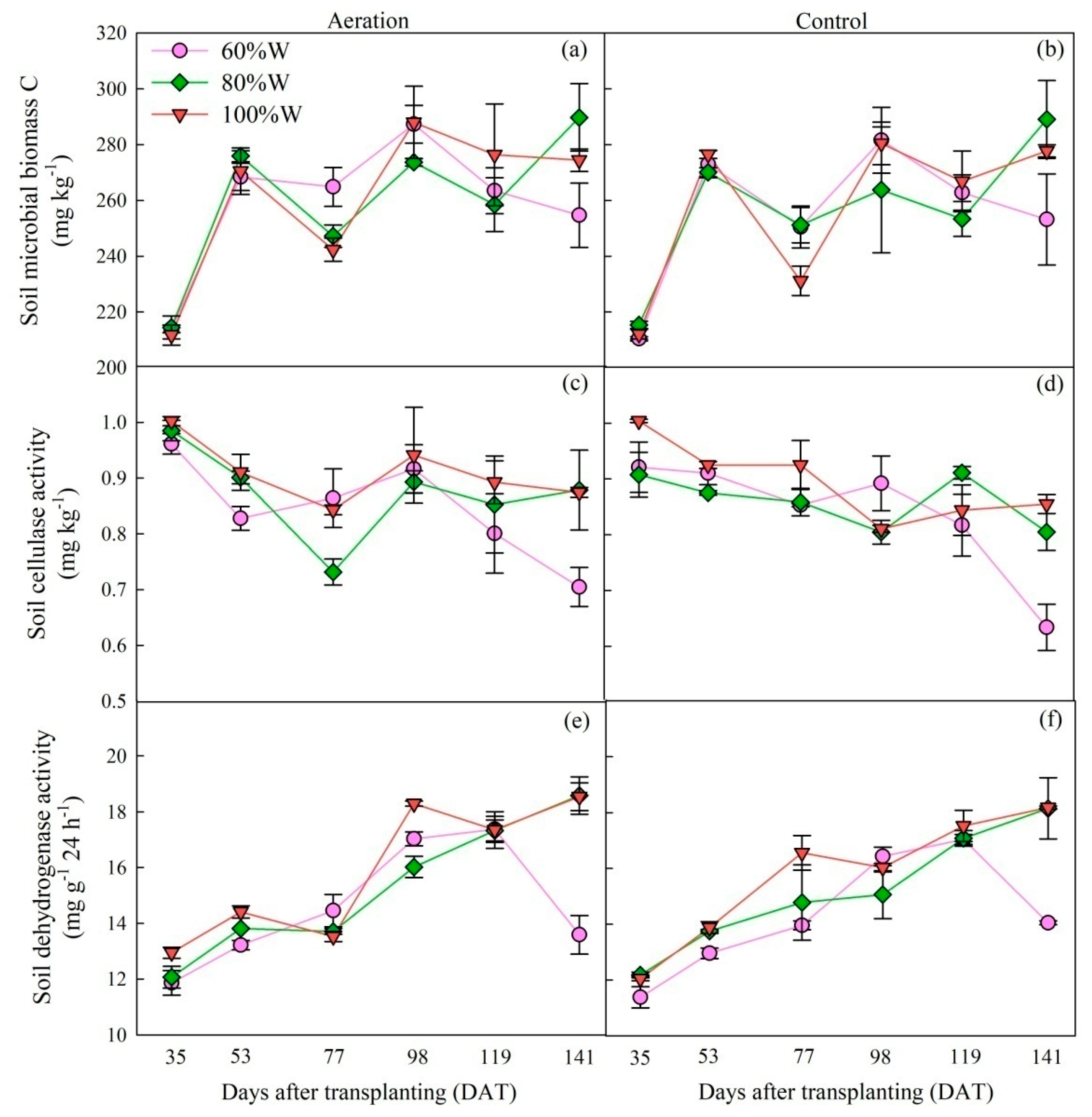
2.1.2. Soil Microbe and Enzyme Activity
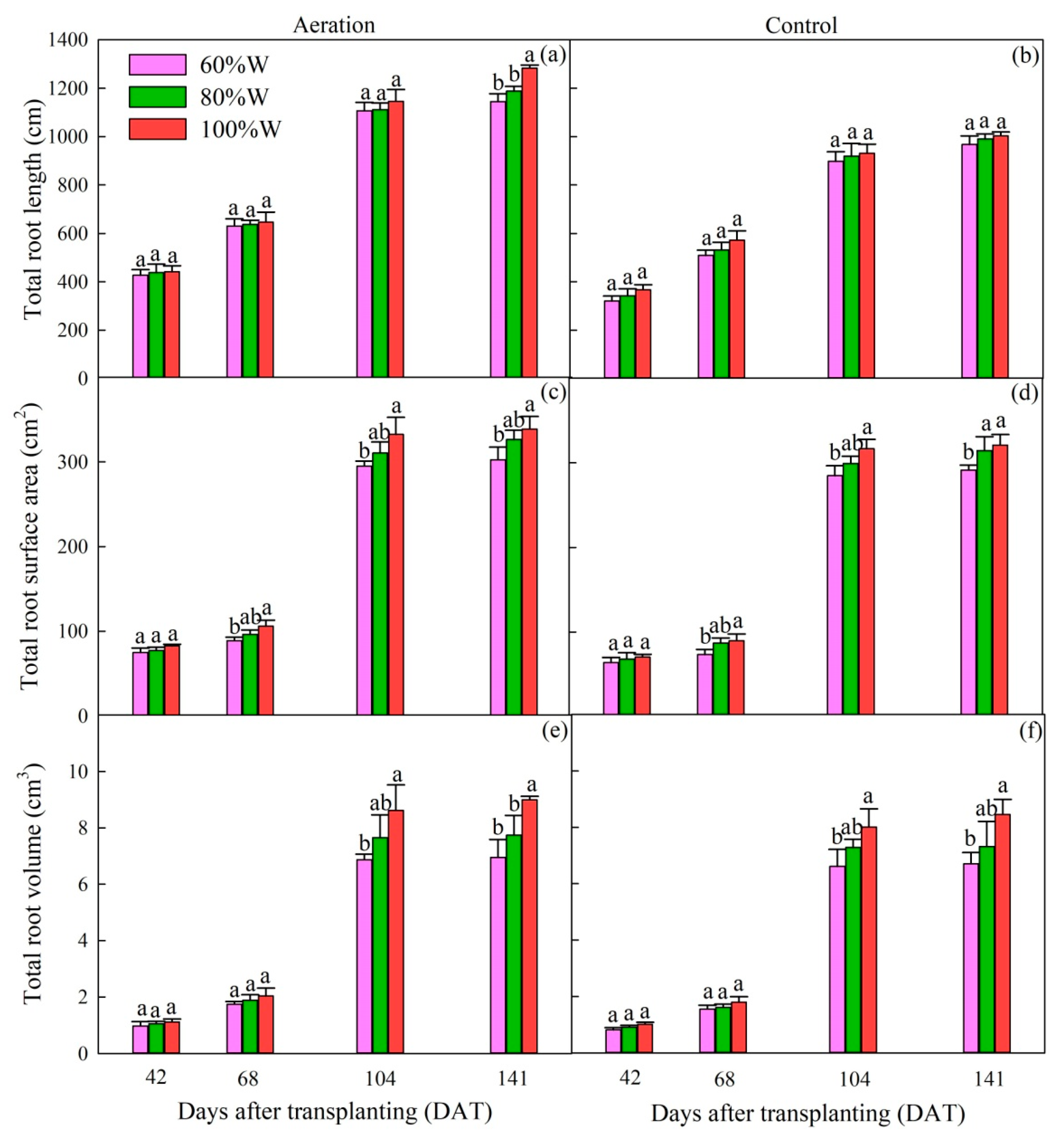
2.1.3. Root Morphology
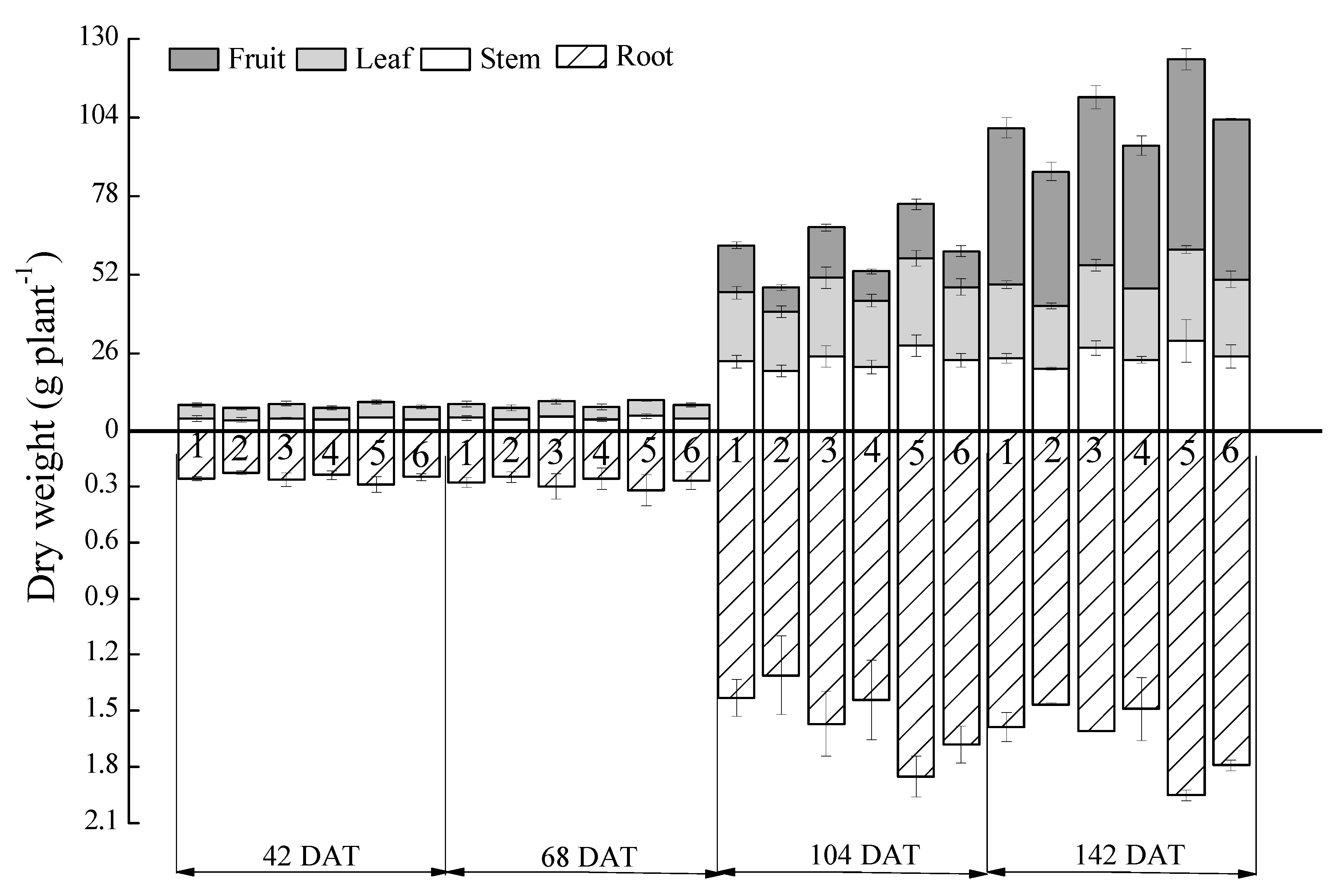
2.1.4. Dry Biomass
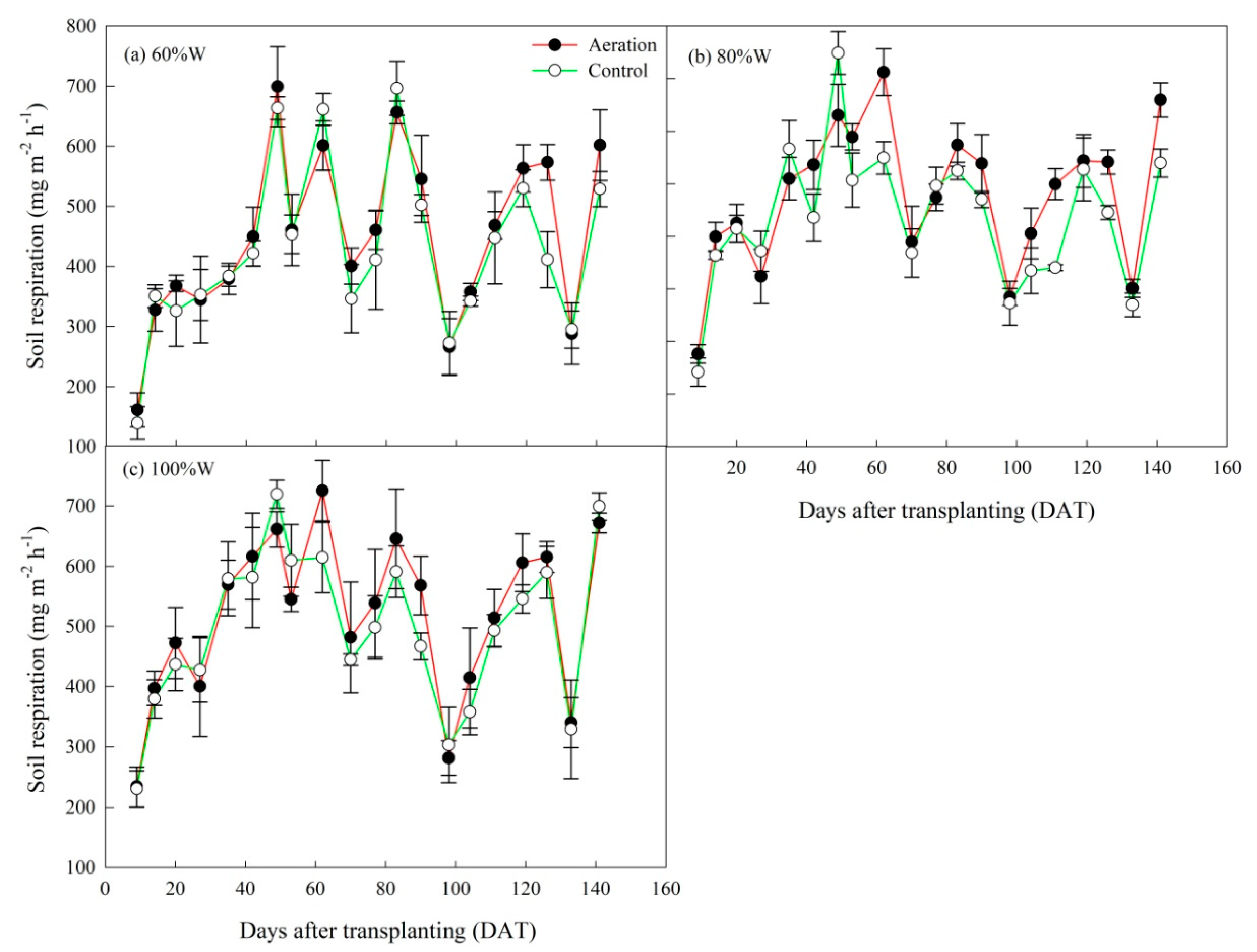
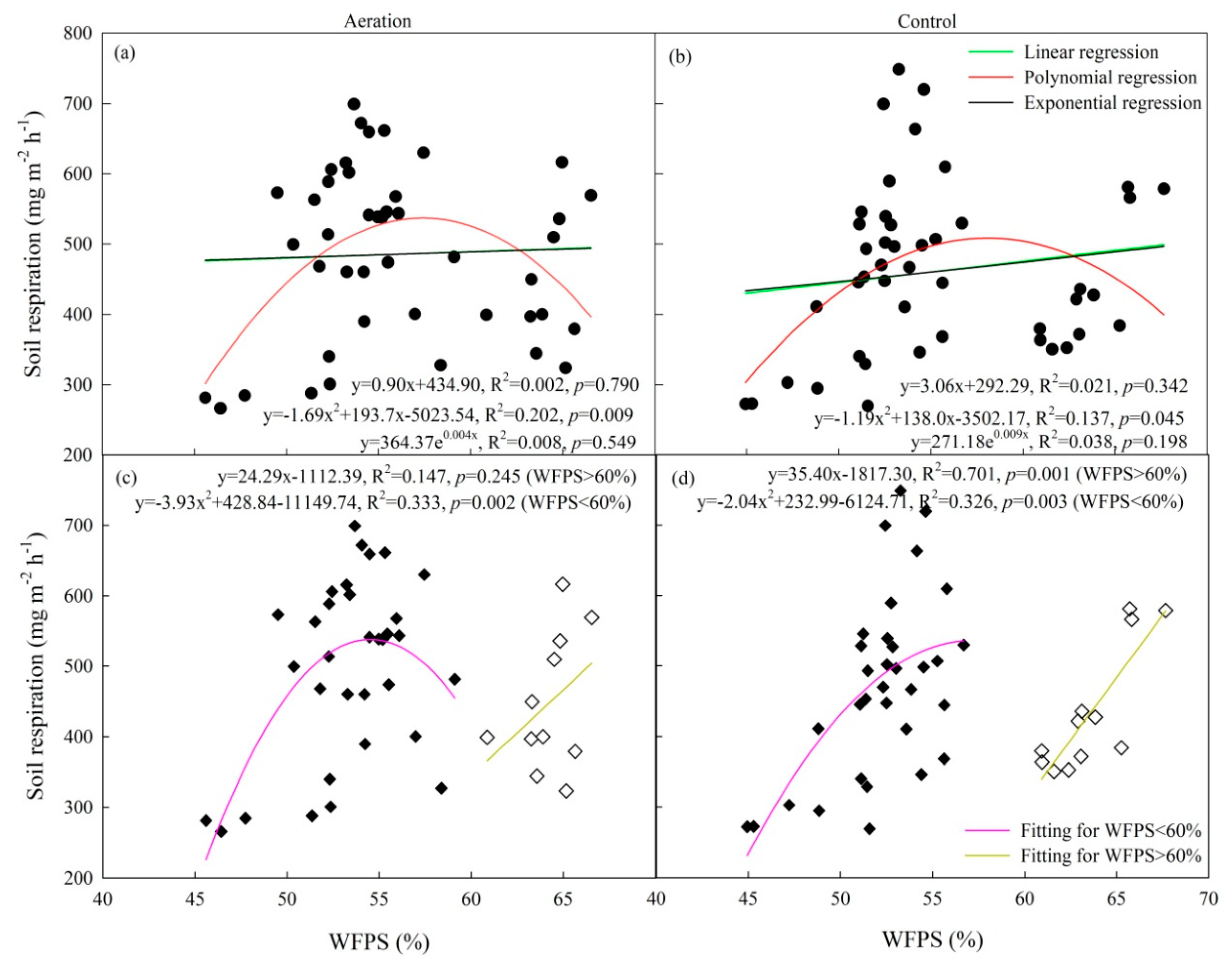
2.2. Soil Respiration
3. Materials and Methods
3.1. Experimental Site
3.2. Experimental Design
3.3. Measurement Index and Methods
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bhattarai, S.P.; Midmore, D.J.; Pendergast, L. Yield, water-use efficiencies and root distribution of soybean, chickpea and pumpkin under different subsurface drip irrigation depths and oxygation treatments in vertisols. Irrig. Sci. 2008, 26, 439–450. [Google Scholar] [CrossRef]
- Mo, Y.; Li, G.; Wang, D. A sowing method for subsurface drip irrigation that increases the emergence rate, yield, and water use efficiency in spring corn. Agric. Water Manag. 2017, 179, 288–295. [Google Scholar] [CrossRef]
- Růžička, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene Regulates Root Growth through Effects on Auxin Biosynthesis and Transport-Dependent Auxin Distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Cai, H.J.; Song, L.B.; Chen, H. Impacts of oxygation on plant growth, yield and fruit quality of tomato. Trans. Chin. Soc. Agric. Mach. 2017, 48, 199–211. (In Chinese) [Google Scholar]
- Bhattarai, S.P.; Pendergast, L.; Midmore, D.J. Root aeration improves yield and water use efficiency of tomato in heavy clay and saline soils. Sci. Hortic. 2006, 108, 278–288. [Google Scholar] [CrossRef]
- Dhungel, J.; Torabi, M.; Pendergast, L.; Chen, X.; Bhattarai, S.P.; Midmore, D.J. Impact of oxygation on soil respiration, yield and water use efficiency of three crop species. J. Plant Ecol. 2010, 4, 236–248. [Google Scholar]
- Hanson, P.J.; Edwards, N.; Garten, C.; Andrews, J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Boil. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Chen, H.; Hou, H.-J.; Wang, X.-Y.; Zhu, Y.; Saddique, Q.; Wang, Y.-F.; Cai, H. The effects of aeration and irrigation regimes on soil CO2 and N2O emissions in a greenhouse tomato production system. J. Integr. Agric. 2018, 17, 449–460. [Google Scholar] [CrossRef]
- Hou, H.; Chen, H.; Cai, H.; Yang, F.; Li, D.; Wang, F. CO2 and N2O emissions from Lou soils of greenhouse tomato fields under aerated irrigation. Atmos. Environ. 2016, 132, 69–76. [Google Scholar] [CrossRef]
- Qi, Y.; Xu, M. Separating the effects of moisture and temperature on soil CO2 efflux in a coniferous forest in the Sierra Nevada mountains. Plant Soil 2001, 237, 15–23. [Google Scholar] [CrossRef]
- Shi, W.-Y.; Du, S.; Morina, J.C.; Guan, J.-H.; Wang, K.-B.; Ma, M.-G.; Yamanaka, N.; Tateno, R. Physical and biogeochemical controls on soil respiration along a topographical gradient in a semiarid forest. Agric. For. Meteorol. 2017, 247, 1–11. [Google Scholar] [CrossRef]
- Jia, S.; Wang, Z.; Li, X.; Sun, Y.; Zhang, X.; Liang, A. N fertilization affects on soil respiration, microbial biomass and root respiration in Larix gmelinii and Fraxinus mandshurica plantations in China. Plant Soil 2010, 333, 325–336. [Google Scholar] [CrossRef]
- Yu, C.-L.; Hui, D.; Deng, Q.; Dzantor, E.K.; Fay, P.A.; Shen, W.; Luo, Y. Responses of switchgrass soil respiration and its components to precipitation gradient in a mesocosm study. Plant Soil 2017, 420, 105–117. [Google Scholar] [CrossRef]
- Makoi, J.; Ndakidemi, P. Selected soil enzymes: Examples of their potential roles in the ecosystem. Afr. J. Biotechnol. 2008, 7, 181–191. [Google Scholar]
- Li, Y.; Niu, W.; Wang, J.; Liu, L.; Zhang, M.; Xu, J. Effects of Artificial Soil Aeration Volume and Frequency on Soil Enzyme Activity and Microbial Abundance when Cultivating Greenhouse Tomato. Soil Sci. Soc. Am. J. 2016, 80, 1208. [Google Scholar] [CrossRef]
- Li, J.; Sun, J.; Yang, Y.; Guo, S.; Glick, B.R. Identification of hypoxic-responsive proteins in cucumber roots using a proteomic approach. Plant Physiol. Biochem. 2012, 51, 74–80. [Google Scholar] [CrossRef]
- Li, Y.; Niu, W.; Xu, J.; Wang, J.; Zhang, M.; Lv, W. Root morphology of greenhouse produced muskmelon under sub-surface drip irrigation with supplemental soil aeration. Sci. Hortic. 2016, 201, 287–294. [Google Scholar] [CrossRef]
- Xu, C.-M.; Wang, D.-Y.; Chen, S.; Chen, L.-P.; Zhang, X.-F. Effects of Aeration on Root Physiology and Nitrogen Metabolism in Rice. Rice Sci. 2013, 20, 148–153. [Google Scholar] [CrossRef]
- Jia, B.; Zhou, G.; Wang, F.; Wang, Y.; Yuan, W.; Zhou, L. Partitioning root and microbial contributions to soil respiration in Leymus chinensis populations. Soil Boil. Biochem. 2006, 38, 653–660. [Google Scholar] [CrossRef]
- Iqbal, J.; Hu, R.; Feng, M.; Lin, S.; Malghani, S.; Ali, I.M. Microbial biomass, and dissolved organic carbon and nitrogen strongly affect soil respiration in different land uses: A case study at Three Gorges Reservoir Area, South China. Agric. Ecosyst. Environ. 2010, 137, 294–307. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Niu, W.; Zhang, Q.; Cui, B.; Gu, X.; Guo, L.; Liang, B. Effects of Nitrogen on Soil Microbial Abundance, Enzyme Activity, and Nitrogen Use Efficiency in Greenhouse Celery under Aerated Irrigation. Soil Sci. Soc. Am. J. 2018, 82, 606. [Google Scholar] [CrossRef]
- Manzoni, S.; Schimel, J.P.; Porporato, A. Responses of soil microbial communities to water stress: Results from a meta-analysis. Ecology 2012, 93, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Cai, H.; Song, L.; Chen, H. Aerated Irrigation Promotes Soil Respiration and Microorganism Abundance around Tomato Rhizosphere. Soil Sci. Soc. Am. J. 2019, 83, 1343. [Google Scholar] [CrossRef]
- Chen, S.; Liu, A.; He, C.; Zou, Z. Microbial and enzyme activities in tomato rhizosphere with organic soil cultivation in solar greenhouse. Chin. J. Soil Sci. 2010, 41, 815–818. (In Chinese) [Google Scholar]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Boil. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Drenovsky, R.; Vo, D.; Graham, K.; Scow, K. Soil Water Content and Organic Carbon Availability Are Major Determinants of Soil Microbial Community Composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef]
- Ben-Noah, I.; Friedman, S. Aeration of clayey soils by injecting air through subsurface drippers: Lysimetric and field experiments. Agric. Water Manag. 2016, 176, 222–233. [Google Scholar] [CrossRef]
- Xu, G.-W.; Lu, D.-K.; Wang, H.-Z.; Li, Y. Morphological and physiological traits of rice roots and their relationships to yield and nitrogen utilization as influenced by irrigation regime and nitrogen rate. Agric. Water Manag. 2018, 203, 385–394. [Google Scholar] [CrossRef]
- Chen, H.; Shang, Z.-H.; Cai, H.-J.; Zhu, Y. An Optimum Irrigation Schedule with Aeration for Greenhouse Tomato Cultivations Based on Entropy Evaluation Method. Sustainability 2019, 11, 4490. [Google Scholar] [CrossRef]
- Bai, T.; Li, C.; Li, C.; Liang, D.; Ma, F. Contrasting hypoxia tolerance and adaptation in Malus species is linked to differences in stomatal behavior and photosynthesis. Physiol. Plantarum. 2013, 147, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Intrigliolo, D.S.; Bonet, L.; Nortes, P.A.; Puerto, H.; Nicolas, E.; Bartual, J. Pomegranate trees performance under sustained and regulated deficit irrigation. Irrig. Sci. 2013, 31, 959–970. [Google Scholar] [CrossRef]
- Zhu, Y.; Cai, H.; Song, L.; Chen, H. Oxygation improving soil aeration around tomato root zone in greenhouse. Trans. Chin. Soc. Agric. Eng. 2017, 33, 163–172. (In Chinese) [Google Scholar]
- Borkhuu, B.; Peckham, S.; Ewers, B.; Norton, U.; Pendall, E.; Ewers, B. Does soil respiration decline following bark beetle induced forest mortality? Evidence from a lodgepole pine forest. Agric. For. Meteorol. 2015, 214, 201–207. [Google Scholar] [CrossRef]
- Dieleman, W.I.J.; Luyssaert, S.; Rey, A.; de Angelis, P.; Barton, C.V.M.; Broadmeadow, M.S.J.; Broadmeadow, S.B.; Chigwerewe, K.S.; Crookshanks, M.; Dufrêne, E.; et al. Soil [N] modulates soil C cycling in CO2-fumigated tree stands: A meta-analysis. Plant Cell Environ. 2010, 33, 2001–2011. [Google Scholar] [CrossRef]
- Cheng, W. Rhizosphere priming effect: Its functional relationships with microbial turnover, evapotranspiration, and C–N budgets. Soil Boil. Biochem. 2009, 41, 1795–1801. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Cheng, W. Interactions between soil and tree roots accelerate long-term soil carbon decomposition. Ecol. Lett. 2007, 10, 1046–1053. [Google Scholar] [CrossRef]
- Zak, D.R.; Pregitzer, K.S.; King, J.S.; Holmes, W.E. Elevated atmospheric CO2, fine roots and the response of soil microorganisms: A review and hypothesis. New Phytol. 2000, 147, 201–222. [Google Scholar] [CrossRef]
- Scheer, C.; Grace, P.R.; Rowlings, D.W.; Payero, J. Soil N2O and CO2 emissions from cotton in Australia under varying irrigation management. Nutr. Cycl. Agroecosys. 2013, 95, 43–56. [Google Scholar] [CrossRef]
- Reth, S.; Göckede, M.; Falge, E. CO2 efflux from agricultural soils in Eastern Germany-comparison of a closed chamber system with eddy covariance measurements. Theor. Appl. Climatol. 2005, 80, 105–120. [Google Scholar] [CrossRef]
- Chen, H.; Hou, H.; Hu, H.; Shang, Z.; Zhu, Y.; Cai, H.; Qaisar, S. Aeration of different irrigation levels affects net global warming potential and carbon footprint for greenhouse tomato systems. Sci. Hortic. 2018, 242, 10–19. [Google Scholar] [CrossRef]
- Хазиев, Φ.Х. Soil Enzymes Activities; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Analysis of Variance (p-Value) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| WFPS | Temperature | cfub | cfuf | cfua | MBC | CA | DHA | Rs | |
| Irrigation | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Aeration | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Irrigation × Aeration | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Factor | Analysis of Variance (p-Value) | ||||||
|---|---|---|---|---|---|---|---|
| Root Morphology | Dry Biomass | ||||||
| Length | Surface Area | Volume | Leaf | Stem | Fruit | Root | |
| Irrigation | ns | ns | ns | * | ns | ** | ** |
| Aeration | * | ns | ns | * | * | * | ns |
| Irrigation × Aeration | ns | ns | ns | ns | ns | ns | ns |
| Treatment | Model | R2 | p |
|---|---|---|---|
| Aeration | Rs = −10.579cfub + 588.685 | 0.503 | 0.001 |
| Rs = −9.997cfuf + 571.845 | 0.660 | 0.000 | |
| Rs = 0.257WFPS2 − 36.187WFPS − 0.943cfub − 12.586cfuf + 1799 | 0.702 | 0.001 | |
| Control | Rs = −10.576cfub + 569.316 | 0.491 | 0.001 |
| Rs = −9.142cfuf + 546.848 | 0.608 | 0.000 | |
| Rs = −0.351WFPS2 + 41.536WFPS − 2.142cfub − 5.858cfuf − 664.275 | 0.616 | 0.003 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Shang, Z.; Cai, H.; Zhu, Y. Irrigation Combined with Aeration Promoted Soil Respiration through Increasing Soil Microbes, Enzymes, and Crop Growth in Tomato Fields. Catalysts 2019, 9, 945. https://doi.org/10.3390/catal9110945
Chen H, Shang Z, Cai H, Zhu Y. Irrigation Combined with Aeration Promoted Soil Respiration through Increasing Soil Microbes, Enzymes, and Crop Growth in Tomato Fields. Catalysts. 2019; 9(11):945. https://doi.org/10.3390/catal9110945
Chicago/Turabian StyleChen, Hui, Zihui Shang, Huanjie Cai, and Yan Zhu. 2019. "Irrigation Combined with Aeration Promoted Soil Respiration through Increasing Soil Microbes, Enzymes, and Crop Growth in Tomato Fields" Catalysts 9, no. 11: 945. https://doi.org/10.3390/catal9110945