Microbial Kinetic Resolution of Aroma Compounds Using Solid-State Fermentation


Abstract
:1. Introduction
2. Results and Discussion
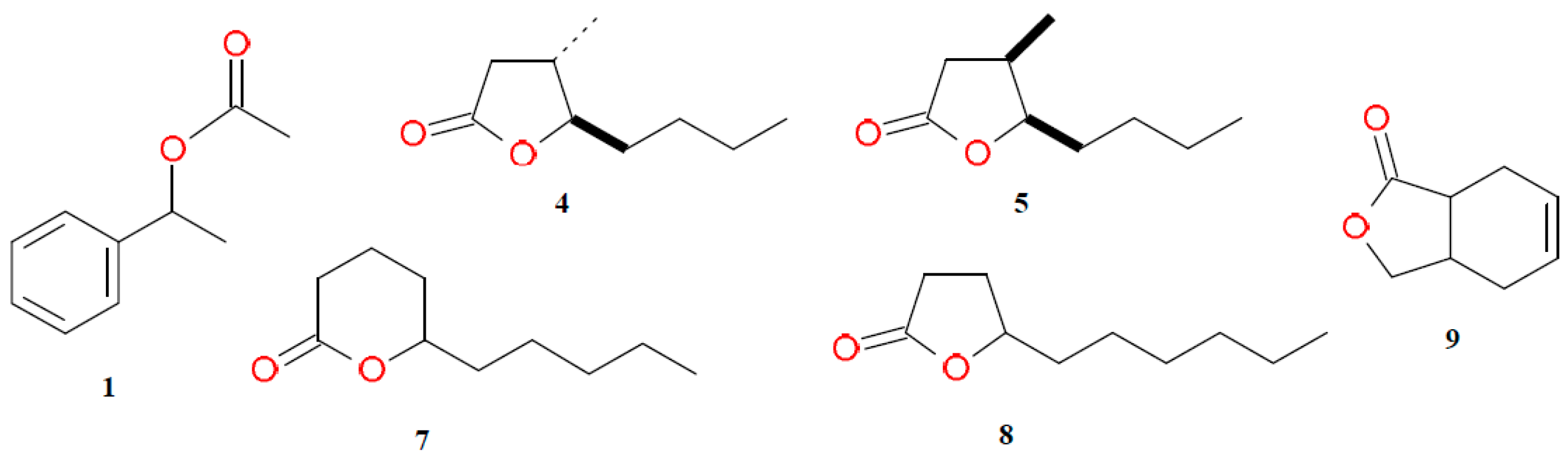
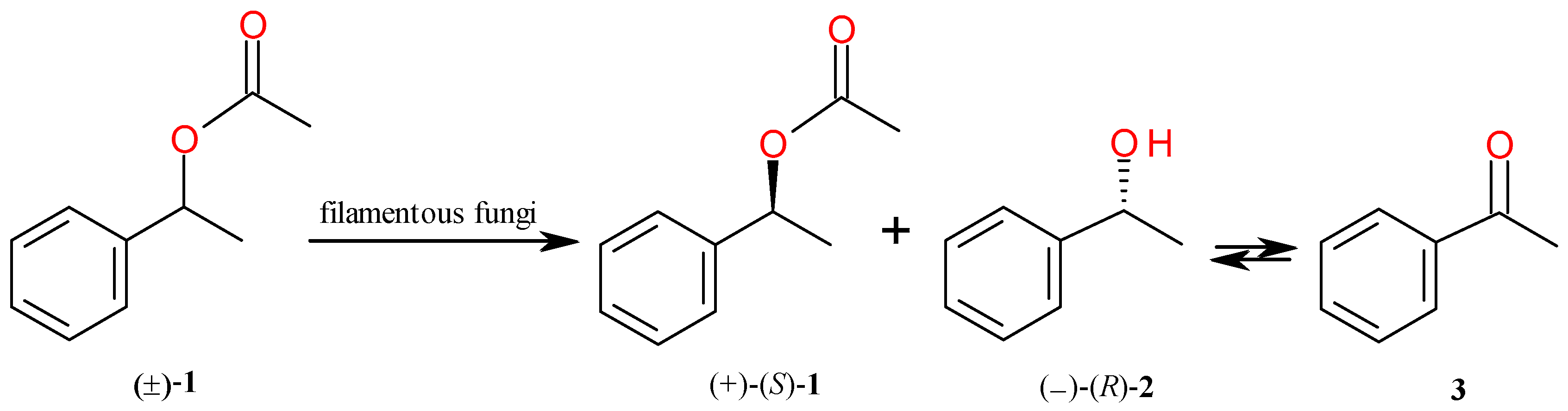
2.1. Kinetic Resolution of 1-Phenylethyl Acetate (1)
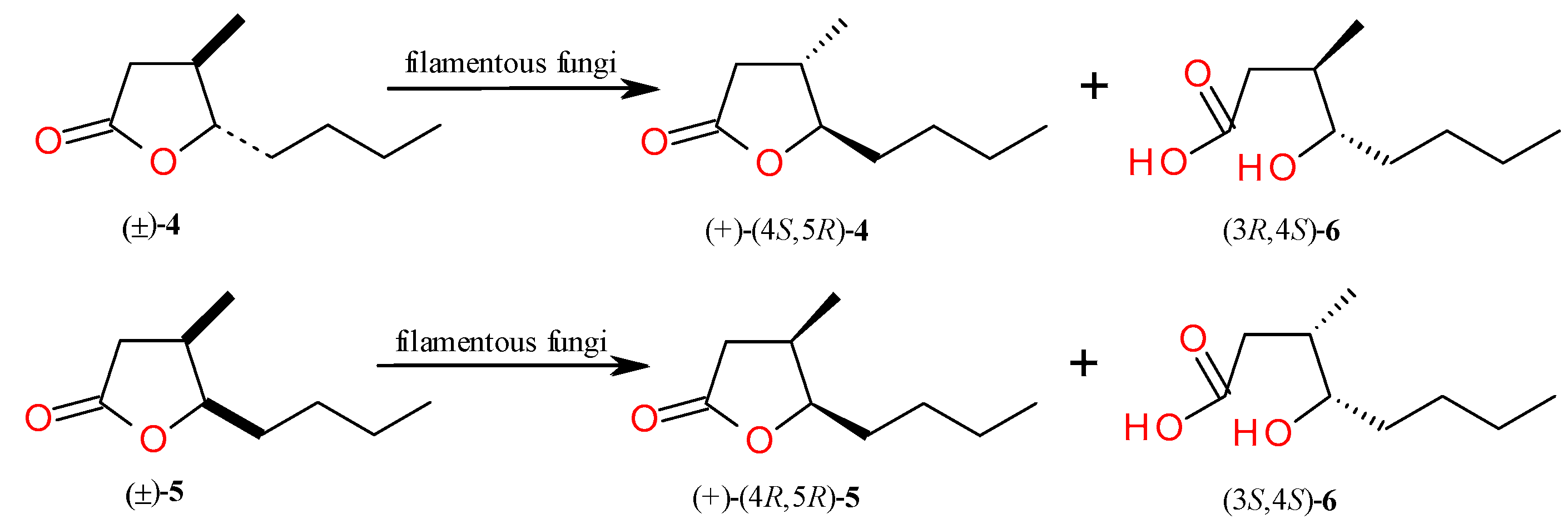
2.2. Kinetic Resolution of trans and cis Whisky Lactones (4, 5)
2.3. Kinetic Resolution of γ-Decalactone (7) and δ-Decalactone (8)
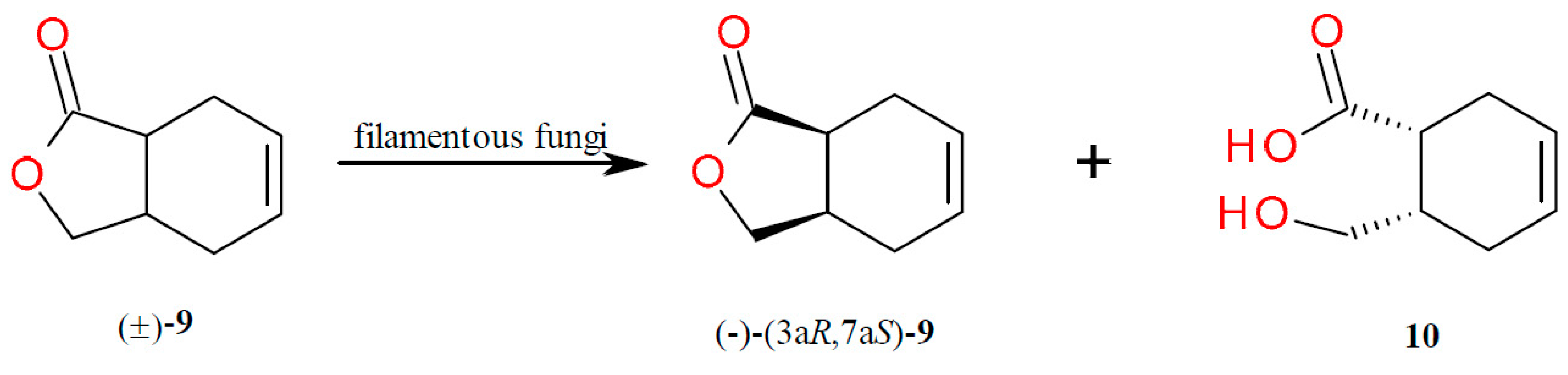
2.4. Kinetic Resolution of cis-3a,4,7,7a-Tetrahydro-1(3H)-isobenzofuranone (9)
3. Materials and Methods
3.1. Materials
3.2. Microorganisms
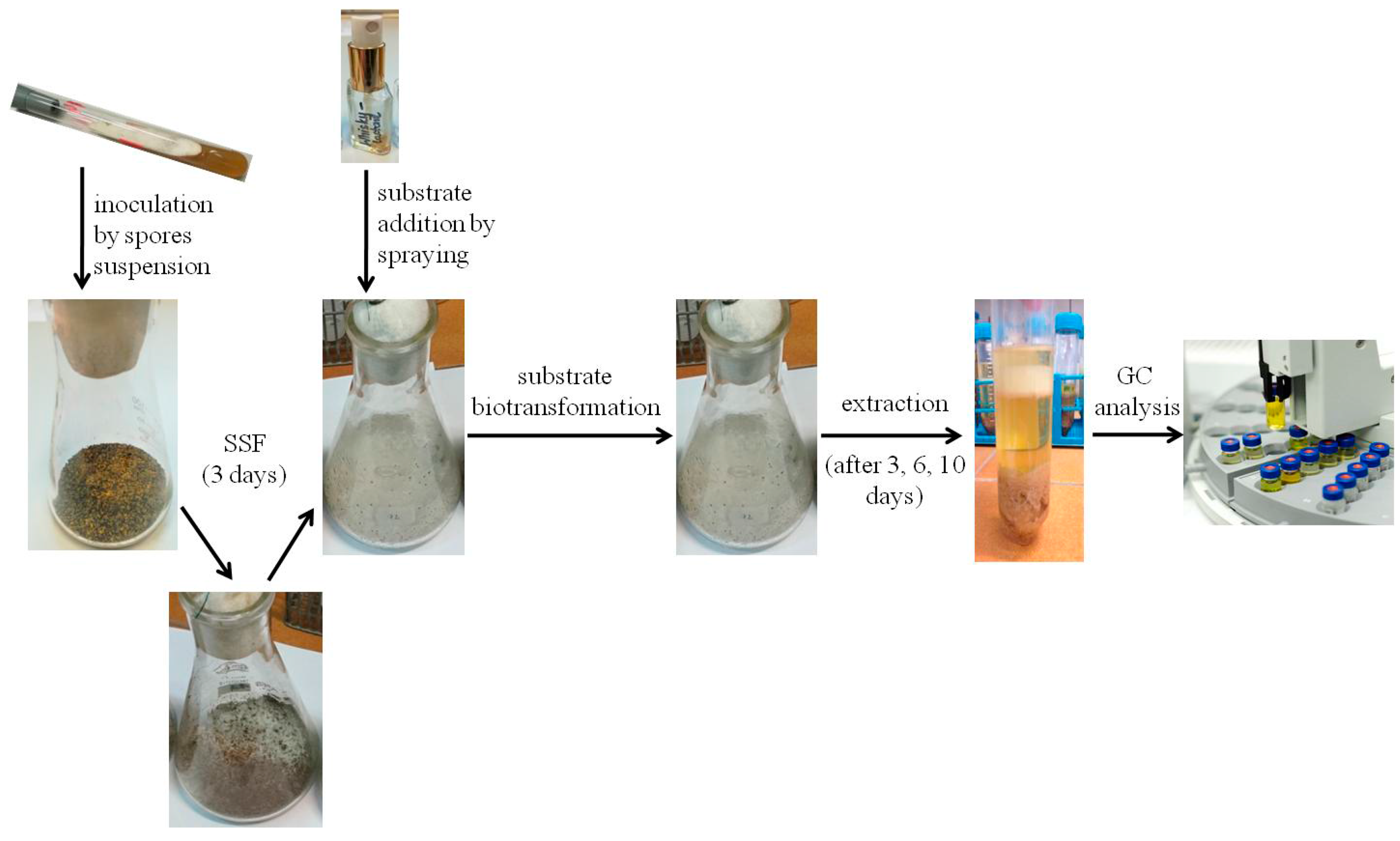
3.3. Solid-State Fermentation
3.4. Enzyme Extraction and Activity Assay
3.5. Biotransformation Process
3.6. Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aggelopoulos, T.; Katsieris, K.; Bekatorou, A.; Pandey, A.; Banat, I.M.; Koutinas, A.A. Solid state fermentation of food waste mixtures for single cell protein, aroma volatiles and fat production. Food Chem. 2014, 145, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Kapilan, R. Solid state fermentation for microbial products: A review. Appl. Sci. Res. 2015, 7, 21–25. [Google Scholar]
- Castilho, L.R.; Polato, C.M.S.; Baruque, E.A.; Sant’Anna, G.L.; Freire, D.M.G. Economic analysis of lipase production by Penicillium restrictum in solid-state and submerged fermentations. Biochem. Eng. J. 2000, 4, 239–247. [Google Scholar] [CrossRef]
- Joo, H.S.; Kumar, C.G.; Park, G.C.; Paik, S.R.; Chang, C.S. Oxidant and SDS-stable alkaline protease from Bacillus clausii I-52: Production and some properties. J. Appl. Microbiol. 2003, 95, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Salihu, A.; Alam, Z.; Abdulkarim, M.I.; Salleh, H.M. Resources, conservation and recycling lipase production: An insight in the utilization of renewable agricultural residues. Resour. Conserv. Recycl. 2012, 58, 36–44. [Google Scholar] [CrossRef]
- De Castro, R.J.S.; Sato, H.H. Enzyme production by solid state fermentation: General aspects and an analysis of the physicochemical characteristics of substrates for agro-industrial wastes valorization. Waste Biomass Valoriz. 2015, 6, 1085–1093. [Google Scholar] [CrossRef]
- Muñoz Solano, D.; Hoyos, P.; Hernáiz, M.J.; Alcántara, A.R.; Sánchez-Montero, J.M. Industrial biotransformations in the synthesis of building blocks leading to enantiopure drugs. Bioresour. Technol. 2012, 115, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Fuhshuku, K.; Oda, S.; Sugai, T. Enzyme reactions as the key step in the synthesis of terpenoids, degraded cartoenoids, steroids, and related substances. Recent Res. Dev. Org. Chem. 2002, 6, 57–74. [Google Scholar]
- Etschmann, M.; Bluemke, W.; Sell, D.; Schrader, J. Biotechnological production of 2-phenylethanol. Appl. Microbiol. Biotechnol. 2002, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; Fuganti, C.; Brenna, E. Biocatalytic preparation of natural flavours and fragrances. Trends Biotechnol. 2005, 23, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Pisani, L.; Superchi, S.; D’Elia, A.; Scafato, P.; Rosini, C. Synthetic approach toward cis-disubstituted γ- and δ-lactones through enantioselective dialkylzinc addition to aldehydes: Application to the synthesis of optically active flavors and fragrances. Tetrahedron 2012, 68, 5779–5784. [Google Scholar] [CrossRef]
- Brenna, E.; Fuganti, C.; Gatti, F.G.; Serra, S. Biocatalytic methods for the synthesis of enantioenriched odor active compounds. Chem. Rev. 2011, 111, 4036–4072. [Google Scholar] [CrossRef] [PubMed]
- Krings, U.; Berger, R.G. Biotechnological production of flavours and fragrances. Appl. Microbiol. Biotechnol. 1998, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Okui, S.; Uchiyama, M.; Mizugaki, M. Metabolism of hydroxy fatty acids II. Intermediates of the oxidative breakdown acid by genus Candida. J. Biochem. 1963, 54, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Van der Schaft, P.H.; ter Burg, N.; van den Bosch, S.; Cohen, A.M. Microbial production of natural δ-decalactone and δ-dodecalactone from the corresponding α,β-unsaturated lactones in Massoi bark oil. Appl. Microbiol. Biotechnol. 1992, 36, 712–716. [Google Scholar] [CrossRef]
- Beck, J.J.; Chou, S.C. The structural diversity of phthalides from the Apiaceae. J. Nat. Prod. 2007, 70, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Gassenmeier, K.; Schwager, H.; Houben, E.C.R. Unequivocal identification of 1-phenylethyl acetate in clove buds (Syzygium aromaticum L.). Foods 2017, 6, 46. [Google Scholar] [CrossRef]
- Alves Macedo, G.; Soberón Lozano, M.M.; Pastore, G.M. Enzymatic synthesis of short chain citronellyl esters by a new lipase from Rhizopus sp. Electron. J. Biotechnol. 2003, 6, 69–72. [Google Scholar] [CrossRef]
- Nagy, V.; Toke, E.R.; Keong, L.C.; Szatzker, G.; Ibrahim, D.; Omar, I.C.; Szakács, G.; Poppe, L. Kinetic resolutions with novel, highly enantioselective fungal lipases produced by solid state fermentation. J. Mol. Catal. B Enzym. 2006, 39, 141–148. [Google Scholar] [CrossRef]
- Kamble, M.P.; Chaudhari, S.A.; Singhal, R.S.; Yadav, G.D. Synergism of microwave irradiation and enzyme catalysis in kinetic resolution of (R,S)-1-phenylethanol by cutinase from novel isolate Fusarium ICT SAC1. Biochem. Eng. J. 2017, 117, 121–128. [Google Scholar] [CrossRef]
- Amin, M.; Bhatti, H.N. Effect of physicochemical parameters on lipase production by Penicillium fellutanum using canola seed oil cake as substrate. Int. J. Agric. Biol. 2014, 16, 118–124. [Google Scholar]
- Freitas, A.C.; Castro, R.J.S.; Fontenele, M.A.; Egito, A.S.; Farinas, C.S.; Pinto, G.A.S. Canola cake as a potential substrate for proteolytic enzymes production by a selected strain of Aspergillus oryzae: Selection of process conditions and product characterization. ISRN Microbiol. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Castro, A.M.; De Andréa, T.V.; Carvalho, D.F.; Teixeira, M.M. P.; Dos Reis Castilho, L.; Freire, D.M.G. Valorization of residual agroindustrial cakes by fungal production of multienzyme complexes and their use in cold hydrolysis of raw starch. Waste Biomass Valoriz. 2011, 2, 291–302. [Google Scholar] [CrossRef]
- Rehman, S. Optimization of process parameters for enhanced production of lipase by Penicillium notatum using agricultural wastes. Afr. J. Biotechnol. 2011, 10, 19580–19589. [Google Scholar] [CrossRef]
- Tan, T.; Zhang, M.; Xu, J.; Zhang, J. Optimization of culture conditions and properties of lipase from Penicillium camembertii Thom PG-3. Process Biochem. 2004, 39, 1495–1502. [Google Scholar] [CrossRef]
- Ramachandran, S.; Singh, S.K.; Larroche, C.; Soccol, C.R.; Pandey, A. Oil cakes and their biotechnological applications—A review. Bioresour. Technol. 2007, 98, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Mueller, K.; Eisner, P.; Yoshie-Stark, Y.; Nakada, R.; Kirchhoff, E. Functional properties and chemical composition of fractionated brown and yellow linseed meal (Linum usitatissimum L.). J. Food Eng. 2010, 98, 453–460. [Google Scholar] [CrossRef]
- Sivaramakrishnan, S.; Gangadharan, D. Edible Oil Cakes. In Biotechnology for Agro-Industrial Residues Utilisation; Singh nee’ Nigam, P., Pandey, A., Eds.; Springer: New Delhi, India, 2009; pp. 253–271. ISBN 978-1-4020-9942-7. [Google Scholar]
- Orsavova, J.; Misurcova, L.; Ambrozova, J.V.; Vicha, R.; Mlcek, J. Fatty acids composition of vegetable oils and its contribution to dietary energy intake and dependence of cardiovascular mortality on dietary intake of fatty acids. Int. J. Mol. Sci. 2015, 16, 12871–12890. [Google Scholar] [CrossRef] [PubMed]
- Bayrak, A.; Kiralan, M.; Ipek, A.; Arslan, N.; Cosge, B.; Khawar, K.M. Fatty acid compositions of linseed (Linum usitatissimum L.) genotypes of different origin cultivated in Turkey. Biotechnol. Biotechnol. Equip. 2010, 24, 1836–1842. [Google Scholar] [CrossRef]
- Chen, H.; He, Q. Value-added bioconversion of biomass by solid-state fermentation. J. Chem. Technol. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Kumar, S.S.; Gupta, R. An extracellular lipase from Trichosporon asahii MSR 54: Medium optimization and enantioselective deacetylation of phenyl ethyl acetate. Process Biochem. 2008, 43, 1054–1060. [Google Scholar] [CrossRef]
- Jadhav, D.D.; Patil, H.S.; Chaya, P.S.; Thulasiram, H.V. Fungal mediated kinetic resolution of racemic acetates to (R)-alcohols using Fusarium proliferatum. Tetrahedron Lett. 2016, 57, 4563–4567. [Google Scholar] [CrossRef]
- Solarte, C.; Yara-Varón, E.; Eras, J.; Torres, M.; Balcells, M.; Canela-Garayoa, R. Lipase activity and enantioselectivity of whole cells from a wild-type Aspergillius flavus strain. J. Mol. Catal. B Enzym. 2014, 100, 78–83. [Google Scholar] [CrossRef]
- Fan, Y.; Xie, Z.; Zhang, H.; Qian, J. Kinetic resolution of both 1-phenylethanol enantiomers produced by hydrolysis of 1-phenylethyl acetate with Candida antarctica lipase B in different solvent systems. Kinet. Catal. 2011, 52, 686–690. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, Y.; Sun, A.; Deng, D.; Hu, Y. Enantioselective resolution of (±)-1-phenylethanol and (±)-1-phenylethyl acetate by a novel esterase from Bacillus sp. SCSIO 15121. Appl. Biochem. Biotechnol. 2016, 178, 558–575. [Google Scholar] [CrossRef] [PubMed]
- Vandenberghe, A.; Markó, I.E.; Lucaccioni, F.; Lutts, S. Enantioselective hydrolysis of racemic 1-phenylethyl acetate by an enzymatic system from fresh vegetables. Ind. Crops Prod. 2013, 42, 380–385. [Google Scholar] [CrossRef]
- Deng, D.; Zhang, Y.; Sun, A.; Hu, Y. Enantio-selective preparation of (S)-1-phenylethanol by a novel marine GDSL lipase MT6 with reverse stereo-selectivity. Chin. J. Catal. 2016, 37, 1966–1974. [Google Scholar] [CrossRef]
- Boratyński, F.; Smuga, M.; Wawrzeńczyk, C. Lactones 42. Stereoselective enzymatic/microbial synthesis of optically active isomers of whisky lactone. Food Chem. 2013, 141, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Boratyński, F.; Dancewicz, K.; Paprocka, M.; Gabrys̈, B.; Wawrzeńczyk, C. Chemo-enzymatic synthesis of optically active γ- and δ-decalactones and their effect on aphid probing, feeding and settling behavior. PLoS ONE 2016, 11, e0146160. [Google Scholar] [CrossRef] [PubMed]
- Boratyński, F.; Pannek, J.; Walczak, P.; Janik-Polanowicz, A.; Huszcza, E.; Szczepańska, E.; Martinez-Rojas, E.; Olejniczak, T. Microbial alcohol dehydrogenase screening for enantiopure lactone synthesis: Down-stream process from microtiter plate to bench bioreactor. Process Biochem. 2014, 49. [Google Scholar] [CrossRef]
- Walczak, P.; Pannek, J.; Boratyński, F.; Janik-Polanowicz, A.; Olejniczak, T. Synthesis and fungistatic activity of bicyclic lactones and lactams against Botrytis cinerea, Penicillium citrinum, and Aspergillus glaucus. J. Agric. Food Chem. 2014, 62, 8571–8578. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Time (Days) | Lipase Activity (U/g) | Conversion (%) | (S)-1 | (R)-2 | 3 | ||
|---|---|---|---|---|---|---|---|---|
| (%) | ee (%) | (%) | ee (%) | (%) | ||||
| Aspergillus candidus AM386 | 3 | 40.4 | 100 | 0 | - | 86 | >99 | 14 |
| Aspergillus nidulans AM243 | 3 | 106.6 | 100 | 0 | - | 100 | >99 | 0 |
| Aspergillus ochraceus AM370 | 3 | 21 | 100 | 0 | - | 0 | - | 100 |
| Aspergillus ochraceus AM456 | 3 | 90.3 | 88 | 12 | 74 | 88 | 22 | 0 |
| Aspergillus wenthi AM413 | 3 | 96.8 | 30 | 70 | 30 | 26 | 64 | 4 |
| Botrytis cirenea AM235 | 6 | 144.7 | 100 | 0 | - | 96 | 98 | 4 |
| Fusarium avenaceum AM11 | 3 | 46 | 100 | 0 | - | 0 | - | 100 |
| Fusarium oxysporum AM21 | 3 | 146.5 | 18 | 82 | >99 | 15 | 44 | 3 |
| Fusarium semitectum AM20 | 3 | 105.5 | 96 | 4 | >99 a | 56 | >99 | 40 |
| Fusarium tricinctum AM16 | 3 | 67 | 25 | 75 | 20 | 25 | 66 | 0 |
| Mucor spinosus AM398 | 3 | 54.1 | 94 | 6 | 66 a | 78 | >99 | 16 |
| Papularia rosea AM17 | 6 | 160.7 | 97 | 3 | 0 | 88 | 96 | 9 |
| Penicillum camemberti AM83 | 3 | 144.3 | 100 | 0 | - | 100 | >99 | 0 |
| Penicillium chrysogenum AM112 | 6 | 105.1 | 100 | 0 | - | 100 | 62 | 0 |
| Penicillium thomi AM91 | 6 | 155.5 | 98 | 2 | >99 a | 60 | 54 | 38 |
| Poria placenta AM38 | 10 | 22.1 | 98 | 2 | >99 a | 84 | 92 | 14 |
| Spicoria divaricata AM423 | 3 | 89.9 | 93 | 7 | >99 | 59 | 0 | 34 |
| Strain | Time (Days) | Lipase Activity (U/g) | Conversion (%) | (S)-1 | (R)-2 | 3 | ||
|---|---|---|---|---|---|---|---|---|
| (%) | ee (%) | (%) | ee (%) | (%) | ||||
| Aspergillus nidulans AM243 | 3 | 106.6 | 100 | 0 | - | 100 | >99 | 0 |
| Aspergillus ochraceus AM370 | 3 | 21 | 100 | 0 | - | 0 | - | 100 |
| Aspergillus ochraceus AM456 | 3 | 90.3 | 83 | 17 | 40 | 83 | 30 | 0 |
| Aspergillus wenthi AM413 | 10 | 237.5 | 90 | 10 | 24 | 78 | 0 | 12 |
| Botrytis cirenea AM235 | 3 | 14 | 10 | 90 | 80 | 0 | - | 10 |
| Fusarium avenaceum AM11 | 3 | 46 | 100 | 0 | - | 100 | >99 | 0 |
| Fusarium oxysporum AM21 | 10 | 26.6 | 75 | 25 | 62 | 59 | 0 | 16 |
| Fusarium semitectum AM20 | 3 | 105.5 | 70 | 30 | 72 | 34 | 38 | 36 |
| Fusarium tricinctum AM16 | 3 | 67 | 44 | 56 | 26 | 22 | 86 | 22 |
| Mucor spinosus AM398 | 3 | 54.1 | 100 | 0 | 0 | 95 | 90 | 5 |
| Papularia rosea AM17 | 6 | 160.7 | 100 | 0 | 0 | 46 | 86 | 54 |
| Penicillum camemberti AM83 | 3 | 144.3 | 30 | 70 | 30 | 30 | 58 | 0 |
| Penicillium chrysogenum AM112 | 6 | 105.1 | 92 | 8 | >99 | 67 | 50 | 25 |
| Penicillium notatum AM904 | 6 | 36.5 | 100 | 0 | 0 | 92 | 0 | 8 |
| Penicillium thomi AM91 | 3 | 39.4 | 100 | 0 | - | 0 | - | 100 |
| Sclerophoma pythiophila AR55 | 3 | 5.6 | 48 | 52 | 52 | 35 | 20 | 13 |
| Spicoria divaricata AM423 | 3 | 20.1 | 100 | 0 | - | 85 | 26 | 15 |
| Strain | Time (Days) | Lipase Activity (U/g) | trans/cis Ratio (%) | trans-(+)-(4S,5R)-4 ee (%) | cis-(+)-(4R,5R)-5 ee (%) |
|---|---|---|---|---|---|
| Aspergillus sp. AM31 | 6 | 18.1 | 59/41 | 38 | 28 |
| Fusarium culmorum AM9 | 3 | 3 | 48/52 | 12 | 14 |
| Fusarium equiseti AM15 | 3 | 5.8 | 50/50 | 24 a | 18 |
| Fusarium oxysporum AM13 | 6 | 25 | 56/44 | 56 | 60 |
| Papularia rosea AM17 | 6 | 160.7 | 33/67 | 70 | 42 |
| Penicillum camemberti AM83 | 3 | 144.3 | 77/23 | 32 | 0 |
| Penicillium chrysogenum AM112 | 10 | 49.2 | 77/23 | 42 | 14 |
| Penicillium notatum AM904 | 6 | 36.5 | 58/42 | 52 | 12 |
| Pycnidiella resinae AR50 | 6 | 19.8 | 27/73 | 40 | 0 |
| Strain | Time (Days) | Lipase Activity (U/g) | trans/cis Ratio (%) | trans-(+)-(4S,5R)-4 ee (%) | cis-(+)-(4R,5R)-5 ee (%) |
|---|---|---|---|---|---|
| Aspergillus nidulans AM243 | 6 | 47.2 | 46/54 | 90 | 0 |
| Aspergillus ochraceus AM456 | 6 | 15.4 | 65/35 | 0 | 14 |
| Fusarium avenaceum AM11 | 6 | 61.8 | 52/48 | 90 | 14 |
| Fusarium semitectum AM20 | 6 | 87 | 40/60 | 26 | 38 |
| Fusarium solani AM203 | 6 | 87.4 | 54/46 | 90 | 52 |
| Penicillum camembertii AM83 | 6 | 118 | 35/65 | 84 | 0 |
| Penicillium chrysogenum AM112 | 6 | 105.1 | 54/46 | 44 | 0 |
| Penicillium notatum AM904 | 6 | 36.5 | 65/35 | 50 | 0 |
| Penicillium vermiculatum AM30 | 10 | 3.2 | 55/45 | 40 | 10 |
| Sclerophoma pythiophila AM55 | 6 | 9.9 | 56/44 | 28 | 12 |
| Strain | Time (Days) | Lipase Activity (U/g) | (−)-(3aR,7aS)-9 ee (%) | |
|---|---|---|---|---|
| RC | LC | |||
| Aspergillus nidulans AM243 | 3 | 106.6 | 34 | 26 |
| Aspergillus wenthi AM413 | 3 | 96.8 | 50 | 0 |
| Botrytis cirenea AM235 | 6 | 144.7 | 0 | 80 |
| Fusarium avenaceum AM11 | 3 | 46 | 10 | 20 |
| Fusarium oxysporum AM21 | 3 | 146.5 | 74 | 16 |
| Fusarium semitectum AM20 | 3 | 105.5 | 66 | 44 |
| Fusarium tricinctum AM16 | 3 | 67 | 12 | 68 |
| Mucor spinosus AM398 | 3 | 54.1 | 38 | 40 |
| Papularia rosea AM17 | 6 | 160.7 | 26 | 10 |
| Penicillum camembertii AM83 | 3 | 144.3 | 34 | 34 |
| Penicillium chrysogenum AM112 | 6 | 105.1 | 36 | 50 |
| Penicillium notatum AM904 | 6 | 36.5 | 40 | 82 |
| Sclerophoma pythiophila AM55 | 3 | 5.6 | 20 | 24 |
| Spicoria divaricata AM423 | 3 | 20.1 | 26 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boratyński, F.; Szczepańska, E.; Grudniewska, A.; Olejniczak, T. Microbial Kinetic Resolution of Aroma Compounds Using Solid-State Fermentation. Catalysts 2018, 8, 28. https://doi.org/10.3390/catal8010028
Boratyński F, Szczepańska E, Grudniewska A, Olejniczak T. Microbial Kinetic Resolution of Aroma Compounds Using Solid-State Fermentation. Catalysts. 2018; 8(1):28. https://doi.org/10.3390/catal8010028
Chicago/Turabian StyleBoratyński, Filip, Ewa Szczepańska, Aleksandra Grudniewska, and Teresa Olejniczak. 2018. "Microbial Kinetic Resolution of Aroma Compounds Using Solid-State Fermentation" Catalysts 8, no. 1: 28. https://doi.org/10.3390/catal8010028