

The Effects of Buffer Nature on Immobilized Lipase Stability Depend on Enzyme Support Loading
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Immobilization of CALB and TLL on Octyl Agarose
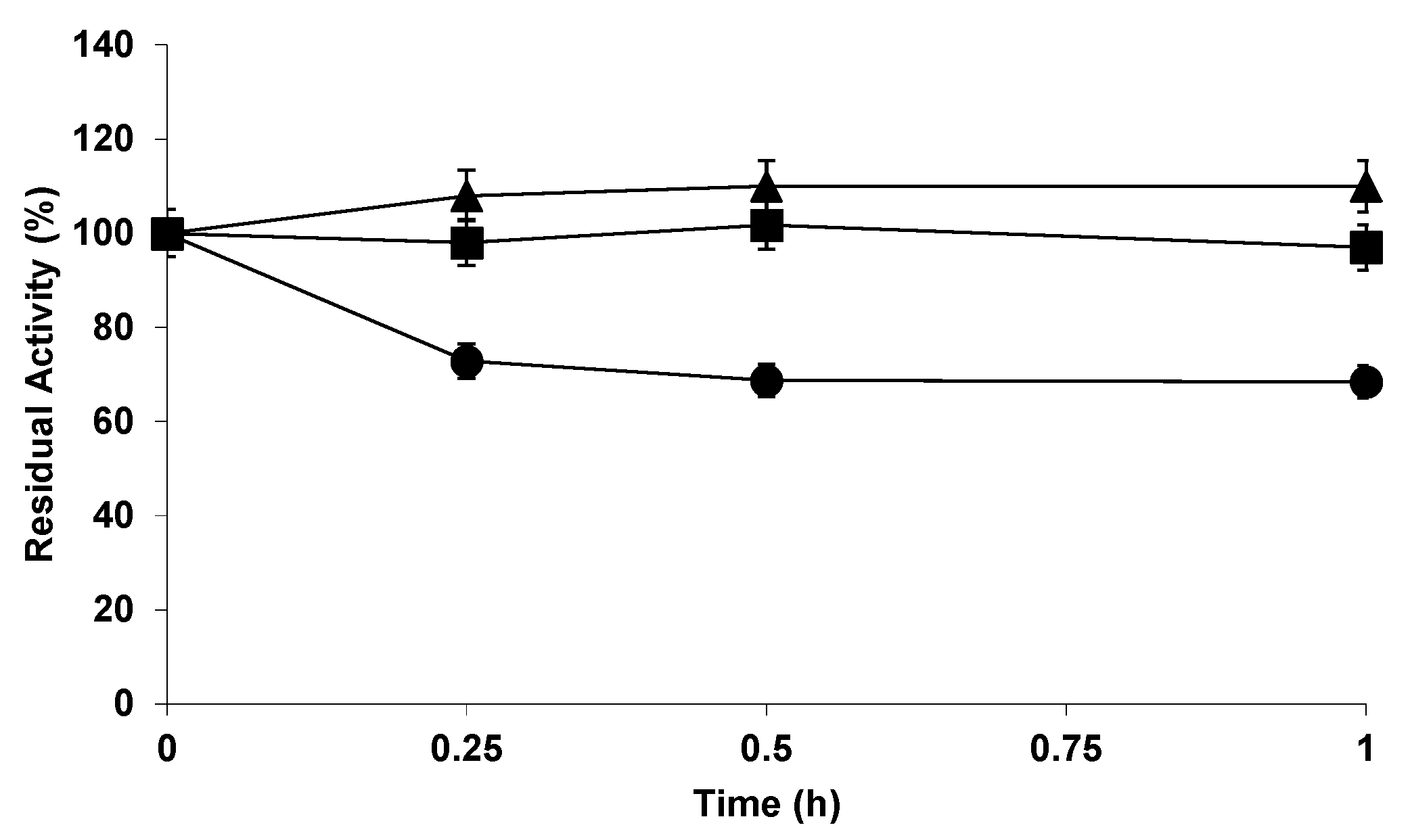
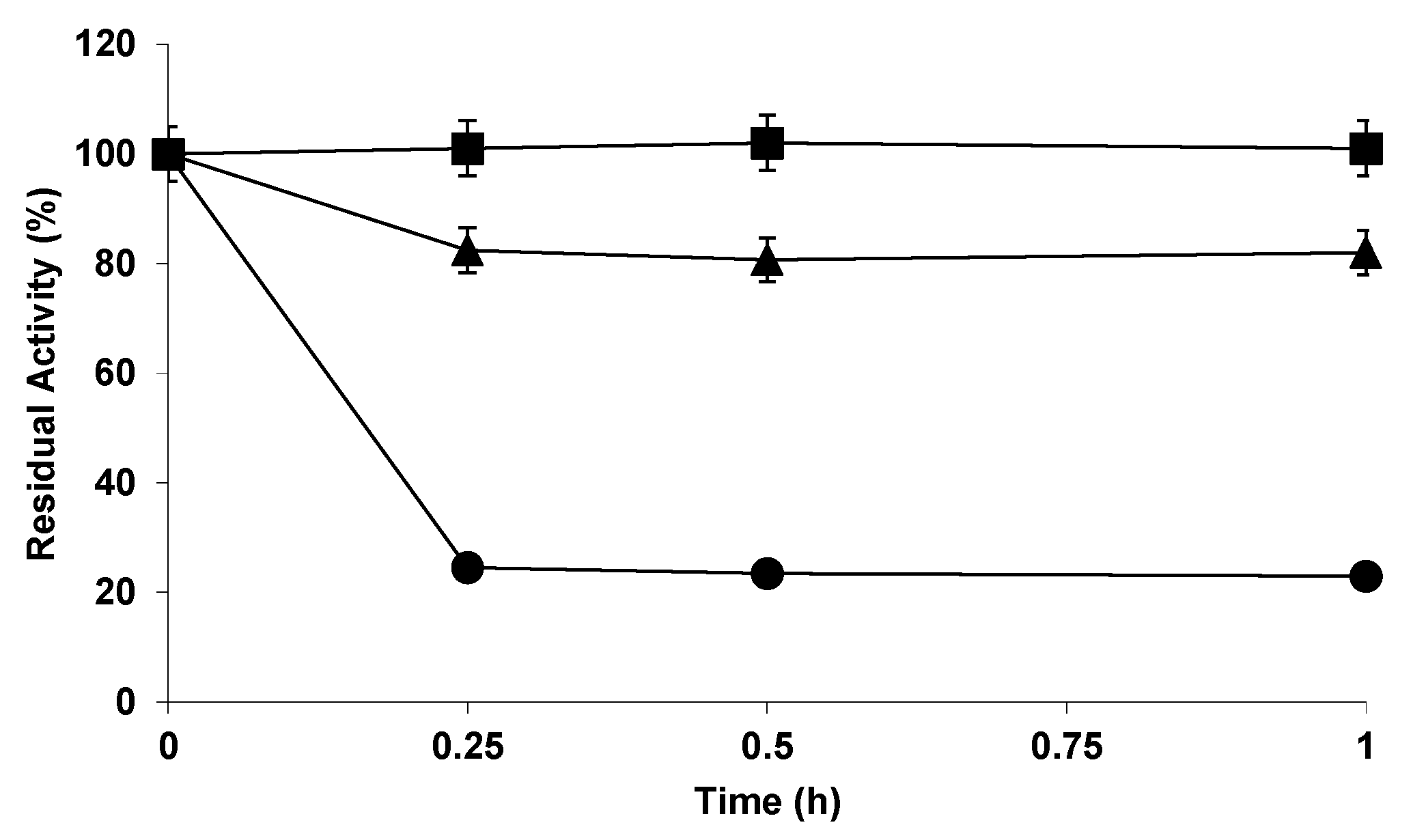
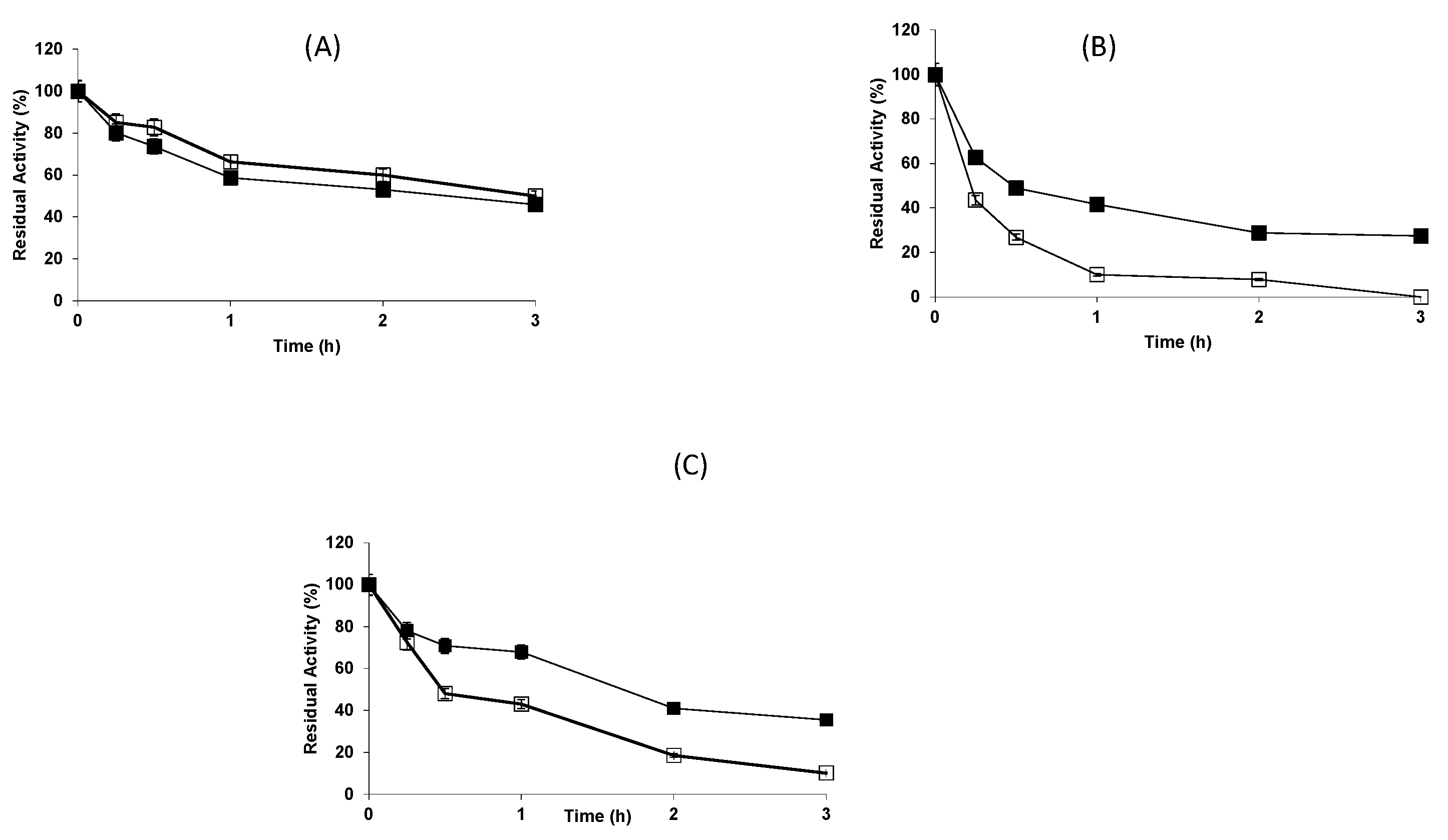
2.2. Effect of the Buffers on the Stability of the Different Biocatalysts
2.3. Effect of Loading and Buffer on Enzyme Activity versus Different Substrates
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Determination of Enzyme Activity with p-NPB
3.2.2. Hydrolysis of Triacetin
3.2.3. Hydrolysis of (R) or (S)-Methyl Mandelate
3.2.4. Immobilization of the Lipases on Octyl-Agarose Beads
3.2.5. Enzyme Thermal Inactivation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabbani, G.; Ahmad, E.; Ahmad, A.; Khan, R.H. Structural Features, Temperature Adaptation and Industrial Applications of Microbial Lipases from Psychrophilic, Mesophilic and Thermophilic Origins. Int. J. Biol. Macromol. 2023, 225, 822–839. [Google Scholar] [CrossRef]
- Mahfoudhi, A.; Benmabrouk, S.; Fendri, A.; Sayari, A. Fungal Lipases as Biocatalysts: A Promising Platform in Several Industrial Biotechnology Applications. Biotechnol. Bioeng. 2022, 119, 3370–3392. [Google Scholar] [CrossRef]
- Remonatto, D.; Miotti, R.H.; Monti, R.; Bassan, J.C.; de Paula, A.V. Applications of Immobilized Lipases in Enzymatic Reactors: A Review. Process Biochem. 2022, 114, 1–20. [Google Scholar] [CrossRef]
- Salgado, C.A.; dos Santos, C.I.A.; Vanetti, M.C.D. Microbial Lipases: Propitious Biocatalysts for the Food Industry. Food Biosci. 2022, 45, 101509. [Google Scholar] [CrossRef]
- He, H.; Han, H.; Shi, H.; Tian, Y.; Sun, F.; Song, Y.; Li, Q.; Zhu, G. Construction of Thermophilic Lipase-Embedded Metal-Organic Frameworks via Biomimetic Mineralization: A Biocatalyst for Ester Hydrolysis and Kinetic Resolution. ACS Appl. Mater. Interfaces 2016, 8, 24517–24524. [Google Scholar] [CrossRef]
- Goswami, D.; Basu, J.K.; De, S. Lipase Applications in Oil Hydrolysis with a Case Study on Castor Oil: A Review. Crit. Rev. Biotechnol. 2013, 33, 81–96. [Google Scholar] [CrossRef]
- Sun, B.; Chen, J.; Sun, C.; Jiang, B.; Zhang, P.; Ma, Y.; Zheng, S.; Tang, K. Enhanced MOF-Immobilized Lipase CALA with Polyethylene Glycol for Efficient Stereoselective Hydrolysis of Aromatic Acid Esters. Biochem. Eng. J. 2022, 189, 108707. [Google Scholar] [CrossRef]
- Carvalho, A.C.L.d.M.; de Oliveira, B.R.; Lima, G.V.; Negreiro, J.M.; Oliveira, M.C.F.; de Lemos, T.L.G.; da Silva, M.R.; Fonseca, T.d.S.; Bezerra, R.M.; dos Santos, J.C.S.; et al. Resolution of Racemic Aryloxy-Propan-2-Yl Acetates via Lipase-Catalyzed Hydrolysis: Preparation of Enantiomerically Pure/Enantioenriched Mexiletine Intermediates and Analogs. Catalysts 2022, 12, 1566. [Google Scholar] [CrossRef]
- Stergiou, P.Y.; Foukis, A.; Filippou, M.; Koukouritaki, M.; Parapouli, M.; Theodorou, L.G.; Hatziloukas, E.; Afendra, A.; Pandey, A.; Papamichael, E.M. Advances in Lipase-Catalyzed Esterification Reactions. Biotechnol. Adv. 2013, 31, 1846–1859. [Google Scholar] [CrossRef] [PubMed]
- Dhake, K.P.; Thakare, D.D.; Bhanage, B.M. Lipase: A Potential Biocatalyst for the Synthesis of Valuable Flavour and Fragrance Ester Compounds. Flavour. Fragr. J. 2013, 28, 71–83. [Google Scholar] [CrossRef]
- Sousa, R.R.; Silva, A.S.A.; Fernandez-Lafuente, R.; Ferreira-Leitão, V.S. Solvent-Free Esterifications Mediated by Immobilized Lipases: A Review from Thermodynamic and Kinetic Perspectives. Catal. Sci. Technol. 2021, 11, 5696–5711. [Google Scholar] [CrossRef]
- Pereira, A.d.S.; de Souza, A.H.; Fraga, J.L.; Villeneuve, P.; Torres, A.G.; Amaral, P.F.F. Lipases as Effective Green Biocatalysts for Phytosterol Esters’ Production: A Review. Catalysts 2022, 12, 88. [Google Scholar] [CrossRef]
- Moussavou Mounguengui, R.W.; Brunschwig, C.; Baréa, B.; Villeneuve, P.; Blin, J. Are Plant Lipases a Promising Alternative to Catalyze Transesterification for Biodiesel Production? Prog. Energy Combust. Sci. 2013, 39, 441–456. [Google Scholar] [CrossRef]
- Narwal, S.K.; Gupta, R. Biodiesel Production by Transesterification Using Immobilized Lipase. Biotechnol. Lett. 2013, 35, 479–490. [Google Scholar] [CrossRef]
- Bajaj, A.; Lohan, P.; Jha, P.N.; Mehrotra, R. Biodiesel Production through Lipase Catalyzed Transesterification: An Overview. J. Mol. Catal. B Enzym. 2010, 62, 9–14. [Google Scholar] [CrossRef]
- Sahin, N.; Akoh, C.C.; Karaali, A. Lipase-Catalyzed Acidolysis of Tripalmitin with Hazelnut Oil Fatty Acids and Stearic Acid to Produce Human Milk Fat Substitutes. J. Agric. Food Chem. 2005, 53, 5779–5783. [Google Scholar] [CrossRef]
- Akil, E.; Pereira, A.d.S.; El-Bacha, T.; Amaral, P.F.F.; Torres, A.G. Efficient Production of Bioactive Structured Lipids by Fast Acidolysis Catalyzed by Yarrowia lipolytica Lipase, Free and Immobilized in Chitosan-Alginate Beads, in Solvent-Free Medium. Int. J. Biol. Macromol. 2020, 163, 910–918. [Google Scholar] [CrossRef]
- Shimada, Y.; Maruyama, K.; Sugihara, A.; Moriyama, S.; Tominaga, Y. Purification of Docosahexaenoic Acid from Tuna Oil by a Two-Step Enzymatic Method: Hydrolysis and Selective Esterification. J. Am. Oil Chem. Soc. 1997, 74, 1441–1446. [Google Scholar] [CrossRef]
- Zou, X.; Nadege, K.; Ninette, I.; Wen, Y.; Wu, S.; Jiang, X.; Zhang, H.; Jin, Q.; Wang, X. Preparation of Docosahexaenoic Acid-Rich Diacylglycerol-Rich Oil by Lipase-Catalyzed Glycerolysis of Microbial Oil from Schizochytrium Sp. in a Solvent-Free System. JAOCS J. Am. Oil Chem. Soc. 2020, 97, 263–270. [Google Scholar] [CrossRef]
- Okulus, M.; Rychlicka, M.; Gliszczyńska, A. Enzymatic Production of Biologically Active 3-Methoxycinnamoylated Lysophosphatidylcholine via Regioselctive Lipase-Catalyzed Acidolysis. Foods 2022, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Soumanou, M.M.; Pérignon, M.; Villeneuve, P. Lipase-Catalyzed Interesterification Reactions for Human Milk Fat Substitutes Production: A Review. Eur. J. Lipid Sci. Technol. 2013, 115, 270–285. [Google Scholar] [CrossRef]
- Utama, Q.D.; Sitanggang, A.B.; Adawiyah, D.R.; Hariyadi, P. Lipase-Catalyzed Interesterification for the Synthesis of Medium-Long-Medium (MLM) Structured Lipids. Food Technol. Biotechnol. 2019, 57, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Nie, K.; Wang, F.; Deng, L. Optimization of the Lipase-Catalyzed Selective Amidation of Phenylglycinol. Front. Bioeng. Biotechnol. 2020, 7, 486. [Google Scholar] [CrossRef] [PubMed]
- Manova, D.; Gallier, F.; Tak-Tak, L.; Yotava, L.; Lubin-Germain, N. Lipase-Catalyzed Amidation of Carboxylic Acid and Amines. Tetrahedron Lett. 2018, 59, 2086–2090. [Google Scholar] [CrossRef]
- Zhang, L.; Li, F.; Wang, C.; Zheng, L.; Wang, Z.; Zhao, R.; Wang, L. Lipase-Mediated Amidation of Anilines with 1,3-Diketones via C–C Bond Cleavage. Catalysts 2017, 7, 115. [Google Scholar] [CrossRef]
- Vongvilai, P.; Ramström, O. Dynamic Asymmetric Multicomponent Resolution: Lipase-Mediated Amidation of a Double Dynamic Covalent System. J. Am. Chem. Soc. 2009, 131, 14419–14425. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Park, K.M. Lipase and Its Unique Selectivity: A Mini-Review. J. Chem. 2022, 2022, 7609019. [Google Scholar] [CrossRef]
- Chen, H.; Meng, X.; Xu, X.; Liu, W.; Li, S. The Molecular Basis for Lipase Stereoselectivity. Appl. Microbiol. Biotechnol. 2018, 102, 3487–3495. [Google Scholar] [CrossRef]
- Tsai, S.W. Enantiopreference of Candida antarctica Lipase B toward Carboxylic Acids: Substrate Models and Enantioselectivity Thereof. J. Mol. Catal. B Enzym. 2016, 127, 98–116. [Google Scholar] [CrossRef]
- Lee, J.; Kim, N.H.; Choi, Y.; Yang, E.; Yu, H.; Kwon, C.W.; Chang, P.S. Divergent Substrate Specificities and Regioselectivities of Three Lipase Isoforms from Cordyceps Militaris: Combinatorial Advantages for Entomopathogenicity and Prospects as Biocatalysts. Enzym. Microb. Technol. 2022, 161, 110117. [Google Scholar] [CrossRef]
- Dong, Z.; Olofsson, K.; Linares-Pastén, J.A.; Karlsson, E.N. Investigation of Structural Features of Two Related Lipases and the Impact on Fatty Acid Specificity in Vegetable Fats. Int. J. Mol. Sci. 2022, 23, 7072. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.; Sanfilippo, C. Stereoselective Promiscuous Reactions Catalyzed by Lipases. Int. J. Mol. Sci. 2022, 23, 2675. [Google Scholar] [CrossRef]
- Samsonowicz-Górski, J.; Koszelewski, D.; Kowalczyk, P.; Śmigielski, P.; Hrunyk, A.; Kramkowski, K.; Wypych, A.; Szymczak, M.; Lizut, R.; Ostaszewski, R. Promiscuous Lipase-Catalyzed Knoevenagel–Phospha–Michael Reaction for the Synthesis of Antimicrobial β-Phosphono Malonates. Int. J. Mol. Sci. 2022, 23, 8819. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Koszelewski, D.; Gawdzik, B.; Samsonowicz-Górski, J.; Kramkowski, K.; Wypych, A.; Lizut, R.; Ostaszewski, R. Promiscuous Lipase-Catalyzed Markovnikov Addition of H-Phosphites to Vinyl Esters for the Synthesis of Cytotoxic α-Acyloxy Phosphonate Derivatives. Materials 2022, 15, 1975. [Google Scholar] [CrossRef] [PubMed]
- Guezane-Lakoud, S.; Toffano, M.; Aribi-Zouioueche, L. Promiscuous Lipase Catalyzed a New P–C Bond Formation: Green and Efficient Protocol for One-Pot Synthesis of α-Aminophosphonates. Heteroat. Chem. 2017, 28, e21408. [Google Scholar] [CrossRef]
- Uppenberg, J.; Hansen, M.T.; Patkar, S.; Jones, T.A. The Sequence, Crystal Structure Determination and Refinement of Two Crystal Forms of Lipase B from Candida antarctica. Structure 1994, 2, 293–308. [Google Scholar] [CrossRef]
- Brzozowski, A.M.; Derewenda, U.; Derewenda, Z.S.; Dodson, G.G.; Lawson, D.M.; Turkenburg, J.P.; Bjorkling, F.; Huge-Jensen, B.; Patkar, S.A.; Thim, L. A Model for Interfacial Activation in Lipases from the Structure of a Fungal Lipase-Inhibitor Complex. Nature 1991, 351, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Verger, R. ‘Interfacial Activation’ of Lipases: Facts and Artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- van Tilbeurgh, H.; Egloff, M.-P.; Martinez, C.; Rugani, N.; Verger, R.; Cambillau, C. Interfacial Activation of the Lipase–Procolipase Complex by Mixed Micelles Revealed by X-ray Crystallography. Nature 1993, 362, 814–820. [Google Scholar] [CrossRef]
- Grochulski, P.; Li, Y.; Schrag, J.D.; Bouthillier, F.; Smith, P.; Harrison, D.; Rubin, B.; Cygler, M. Insights into Interfacial Activation from an Open Structure of Candida rugosa Lipase. J. Biol. Chem. 1993, 268, 12843–12847. [Google Scholar] [CrossRef]
- Derewenda, U.; Brzozowski, A.M.; Lawson, D.M.; Derewenda, Z.S. Catalysis at the Interface: The Anatomy of a Conformational Change in a Triglyceride Lipase. Biochemistry 1992, 31, 1532–1541. [Google Scholar] [CrossRef]
- Reis, P.; Holmberg, K.; Watzke, H.; Leser, M.E.; Miller, R. Lipases at Interfaces: A Review. Adv. Colloid. Interface Sci. 2009, 147–148, 237–250. [Google Scholar] [CrossRef]
- Aloulou, A.; Rodriguez, J.A.; Fernandez, S.; van Oosterhout, D.; Puccinelli, D.; Carrière, F. Exploring the Specific Features of Interfacial Enzymology Based on Lipase Studies. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2006, 1761, 995–1013. [Google Scholar] [CrossRef]
- Dragoi, B.; Dumitriu, E. Are Interfacial Biocatalysts Important Tools For Nonpolluting Technologies? 1. Production, Structural Aspects And Mechanism of Lipases. Environ. Eng. Manag. J. 2005, 4, 1–24. [Google Scholar] [CrossRef]
- Schmid, R.D.; Verger, R. Lipases: Interfacial Enzymes with Attractive Applications. Angew. Chem. 1998, 37, 1608–1633. [Google Scholar] [CrossRef]
- Schoemaker, H.E.; Mink, D.L.; WubboLts, M.G. Dispelling the Myths—Biocatalysis in Industrial Synthesis. Science 2003, 299, 1694–1697. [Google Scholar] [CrossRef]
- Ferrer, M.; Martínez-Martínez, M.; Bargiela, R.; Streit, W.R.; Golyshina, O.V.; Golyshin, P.N. Estimating the Success of Enzyme Bioprospecting through Metagenomics: Current Status and Future Trends. Microb. Biotechnol. 2016, 9, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Vieites, J.M.; Guazzaroni, M.-E.; Beloqui, A.; Golyshin, P.N.; Ferrer, M. Metagenomics Approaches in Systems Microbiology. FEMS Microbiol. Rev. 2009, 33, 236–255. [Google Scholar] [CrossRef]
- Martínez-Martínez, M.; Alcaide, M.; Tchigvintsev, A.; Reva, O.; Polaina, J.; Bargiela, R.; Guazzaroni, M.-E.; Chicote, Á.; Canet, A.; Valero, F.; et al. Biochemical Diversity of Carboxyl Esterases and Lipases from Lake Arreo (Spain): A Metagenomic Approach. Appl. Environ. Microbiol. 2013, 79, 3553–3562. [Google Scholar] [CrossRef] [PubMed]
- Peña-García, C.; Martínez-Martínez, M.; Reyes-Duarte, D.; Ferrer, M. High Throughput Screening of Esterases, Lipases and Phospholipases in Mutant and Metagenomic Libraries: A Review. Comb. Chem. High. Throughput Screen. 2016, 19, 605–615. [Google Scholar] [CrossRef]
- Romero, P.A.; Arnold, F.H. Exploring Protein Fitness Landscapes by Directed Evolution. Nat. Rev. Mol. Cell Biol. 2009, 10, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Packer, M.S.; Liu, D.R. Methods for the Directed Evolution of Proteins. Nat. Rev. Genet. 2015, 16, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Renata, H.; Wang, Z.J.; Arnold, F.H. Expanding the Enzyme Universe: Accessing Non-Natural Reactions by Mechanism-Guided Directed Evolution. Angew. Chem. Int. Ed. 2015, 54, 3351–3367. [Google Scholar] [CrossRef] [PubMed]
- Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Design of Artificial Enzymes Bearing Several Active Centers: New Trends, Opportunities and Problems. Int. J. Mol. Sci. 2022, 23, 5304. [Google Scholar] [CrossRef] [PubMed]
- Alonso, S.; Santiago, G.; Cea-Rama, I.; Fernandez-Lopez, L.; Coscolín, C.; Modregger, J.; Ressmann, A.K.; Martínez-Martínez, M.; Marrero, H.; Bargiela, R.; et al. Genetically Engineered Proteins with Two Active Sites for Enhanced Biocatalysis and Synergistic Chemo- and Biocatalysis. Nat. Catal. 2020, 3, 319–328. [Google Scholar] [CrossRef]
- Fernandez-Lopez, L.; Roda, S.; Gonzalez-Alfonso, J.L.; Plou, F.J.; Guallar, V.; Ferrer, M. Design and Characterization of In-One Protease-Esterase PluriZyme. Int. J. Mol. Sci. 2022, 23, 13337. [Google Scholar] [CrossRef] [PubMed]
- Santiago, G.; Martínez-Martínez, M.; Alonso, S.; Bargiela, R.; Coscolín, C.; Golyshin, P.N.; Guallar, V.; Ferrer, M. Rational Engineering of Multiple Active Sites in an Ester Hydrolase. Biochemistry 2018, 57, 2245–2255. [Google Scholar] [CrossRef]
- Kan, S.; Aoyagi-Scharber, M.; Le, S.Q.; Vincelette, J.; Ohmi, K.; Bullens, S.; Wendt, D.J.; Christianson, T.M.; Tiger, P.M.N.; Brown, J.R.; et al. Delivery of an Enzyme-IGFII Fusion Protein to the Mouse Brain Is Therapeutic for Mucopolysaccharidosis Type IIIB. Proc. Natl. Acad. Sci. USA 2014, 111, 14870–14875. [Google Scholar] [CrossRef]
- Lee, D.; Lloyd, N.D.R.; Pretorius, I.S.; Borneman, A.R. Heterologous Production of Raspberry Ketone in the Wine Yeast Saccharomyces cerevisiae via Pathway Engineering and Synthetic Enzyme Fusion. Microb. Cell Fact. 2016, 15, 49. [Google Scholar] [CrossRef]
- Liu, H.-H.; Wang, C.; Lu, X.-Y.; Huang, H.; Tian, Y.; Ji, X.-J. Improved Production of Arachidonic Acid by Combined Pathway Engineering and Synthetic Enzyme Fusion in Yarrowia lipolytica. J. Agric. Food Chem. 2019, 67, 9851–9857. [Google Scholar] [CrossRef]
- Rabeharindranto, H.; Castaño-Cerezo, S.; Lautier, T.; Garcia-Alles, L.F.; Treitz, C.; Tholey, A.; Truan, G. Enzyme-Fusion Strategies for Redirecting and Improving Carotenoid Synthesis in S. cerevisiae. Metab. Eng. Commun. 2019, 8, e00086. [Google Scholar] [CrossRef]
- Camagna, M.; Grundmann, A.; Bär, C.; Koschmieder, J.; Beyer, P.; Welsch, R. Enzyme Fusion Removes Competition for Geranylgeranyl Diphosphate in Carotenogenesis. Plant Physiol. 2019, 179, 1013–1027. [Google Scholar] [CrossRef]
- Nogueira, M.; Enfissi, E.M.A.; Welsch, R.; Beyer, P.; Zurbriggen, M.D.; Fraser, P.D. Construction of a Fusion Enzyme for Astaxanthin Formation and Its Characterisation in Microbial and Plant Hosts: A New Tool for Engineering Ketocarotenoids. Metab. Eng. 2019, 52, 243–252. [Google Scholar] [CrossRef]
- Du, L.; Cui, X.; Li, H.; Wang, Y.; Fan, L.; He, R.; Jiang, F.; Yu, A.; Xiao, D.; Ma, L. Enhancing the Enzymatic Hydrolysis Efficiency of Lignocellulose Assisted by Artificial Fusion Enzyme of Swollenin-Xylanase. Ind. Crop. Prod. 2021, 173, 114106. [Google Scholar] [CrossRef]
- Xia, Y.; Wu, Z.; He, R.; Gao, Y.; Qiu, Y.; Cheng, Q.; Ma, X.; Wang, Z. Simultaneous Degradation of Two Mycotoxins Enabled by a Fusion Enzyme in Food-Grade Recombinant Kluyveromyces lactis. Bioresour. Bioprocess. 2021, 8, 62. [Google Scholar] [CrossRef]
- Liao, L.; Zhang, Y.; Wang, Y.; Fu, Y.; Zhang, A.; Qiu, R.; Yang, S.; Fang, B. Construction and Characterization of a Novel Glucose Dehydrogenase-Leucine Dehydrogenase Fusion Enzyme for the Biosynthesis of l-Tert-Leucine. Microb. Cell Fact. 2021, 20, 3. [Google Scholar] [CrossRef]
- Fabara, A.N.; Fraaije, M.W. Production of Indigo through the Use of a Dual-Function Substrate and a Bifunctional Fusion Enzyme. Enzym. Microb. Technol. 2020, 142, 109692. [Google Scholar] [CrossRef] [PubMed]
- Mourelle-Insua, Á.; Aalbers, F.S.; Lavandera, I.; Gotor-Fernández, V.; Fraaije, M.W. What to Sacrifice? Fusions of Cofactor Regenerating Enzymes with Baeyer-Villiger Monooxygenases and Alcohol Dehydrogenases for Self-Sufficient Redox Biocatalysis. Tetrahedron 2019, 75, 1832–1839. [Google Scholar] [CrossRef]
- Baklouti, Z.; Delattre, C.; Pierre, G.; Gardarin, C.; Abdelkafi, S.; Michaud, P.; Dubessay, P. Biochemical Characterization of a Bifunctional Enzyme Constructed by the Fusion of a Glucuronan Lyase and a Chitinase from Trichoderma sp. Life 2020, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Wang, C.; Shao, S.; Wang, Q.; Zhou, X.; Bai, Y.; Chen, L.; Lu, C.; Zhao, M.; Wang, Z. Efficient Oxidation of Methyl Glycolate to Methyl Glyoxylate Using a Fusion Enzyme of Glycolate Oxidase, Catalase and Hemoglobin. Catalysts 2020, 10, 943. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme Immobilisation in Biocatalysis: Why, What and How. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Woodley, J.M.; Fernandez-Lafuente, R. Is Enzyme Immobilization a Mature Discipline? Some Critical Considerations to Capitalize on the Benefits of Immobilization. Chem. Soc. Rev. 2022, 51, 6251–6290. [Google Scholar] [CrossRef]
- Di Cosimo, R.; Mc Auliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial Use of Immobilized Enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef]
- Liese, A.; Hilterhaus, L. Evaluation of Immobilized Enzymes for Industrial Applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Berenguer-Murcia, Á.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of Enzymes via Immobilization: Multipoint Covalent Attachment and Other Stabilization Strategies. Biotechnol. Adv. 2021, 52, 107821. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.T.; Gu, M.B. Enzyme Stabilization by Nano/Microsized Hybrid Materials. Eng. Life Sci. 2013, 13, 49–61. [Google Scholar] [CrossRef]
- Kim, J.; Grate, J.W.; Wang, P. Nanostructures for Enzyme Stabilization. Chem. Eng. Sci. 2006, 61, 1017–1026. [Google Scholar] [CrossRef]
- Stepankova, V.; Bidmanova, S.; Koudelakova, T.; Prokop, Z.; Chaloupkova, R.; Damborsky, J. Strategies for Stabilization of Enzymes in Organic Solvents. ACS Catal. 2013, 3, 2823–2836. [Google Scholar] [CrossRef]
- Klibanov, A.M. Stabilization of Enzymes against Thermal Inactivation. Adv. Appl. Microbiol. 1983, 29, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.V.; Ananthanarayan, L. Enzyme Stability and Stabilization—Aqueous and Non-Aqueous Environment. Process Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Berenguer-Murcia, Á.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Enzyme Immobilization Strategies to Improve Enzyme Performance. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Bilal, M.; Asgher, M.; Cheng, H.; Yan, Y.; Iqbal, H.M.N. Multi-Point Enzyme Immobilization, Surface Chemistry, and Novel Platforms: A Paradigm Shift in Biocatalyst Design. Crit. Rev. Biotechnol. 2019, 39, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Wahab, R.A.; Elias, N.; Abdullah, F.; Ghoshal, S.K. On the Taught New Tricks of Enzymes Immobilization: An All-Inclusive Overview. React. Funct. Polym. 2020, 152, 104613. [Google Scholar] [CrossRef]
- Bilal, M.; Zhao, Y.; Noreen, S.; Shah, S.Z.H.; Bharagava, R.N.; Iqbal, H.M.N. Modifying Bio-Catalytic Properties of Enzymes for Efficient Biocatalysis: A Review from Immobilization Strategies Viewpoint. Biocatal. Biotransformation 2019, 37, 159–182. [Google Scholar] [CrossRef]
- El-Aziz, S.M.A.; Faraag, A.H.I.; Ibrahim, A.M.; Albrakati, A.; Bakkar, M.R. Tyrosinase Enzyme Purification and Immobilization from Pseudomonas sp. EG22 Using Cellulose Coated Magnetic Nanoparticles: Characterization and Application in Melanin Production. World J. Microbiol. Biotechnol. 2023, 40, 10. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.C.H.T.; Dodds, K.C.; McKenna, C.; Cowan, R.M.; Ganin, A.Y.; Campopiano, D.J.; Yiu, H.H.P. Direct Purification and Immobilization of His-Tagged Enzymes Using Unmodified Nickel Ferrite NiFe2O4 Magnetic Nanoparticles. Sci. Rep. 2023, 13, 21549. [Google Scholar] [CrossRef]
- Zhou, L.; Ouyang, Y.; Kong, W.; Ma, T.; Zhao, H.; Jiang, Y.; Gao, J.; Ma, L. One Pot Purification and Co-Immobilization of His-Tagged Old Yellow Enzyme and Glucose Dehydrogenase for Asymmetric Hydrogenation. Enzym. Microb. Technol. 2022, 156, 110001. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Zhuang, N.; Zhang, A.; Chen, K.; Xu, N.; Xin, F.; Zhang, W.; Dong, W.; Jiang, M. Immobilization and Purification of Enzymes With the Novel Affinity Tag ChBD-AB From Chitinolyticbacter meiyuanensis SYBC-H1. Front. Bioeng. Biotechnol. 2020, 8, 544642. [Google Scholar] [CrossRef]
- Ahmad, A.; Patta, A.M.; Natsir, H. Purification and Immobilization of L-Asparaginase Enzyme from the Thermophilic Bacteria Bacillus licheniformis Strain HSA3-1a. Int. J. Pharma Bio Sci. 2013, 4, B274–B280. [Google Scholar]
- Rodrigues, R.C.; Virgen-Ortíz, J.J.; dos Santos, J.C.S.; Berenguer-Murcia, Á.; Alcantara, A.R.; Barbosa, O.; Ortiz, C.; Fernandez-Lafuente, R. Immobilization of Lipases on Hydrophobic Supports: Immobilization Mechanism, Advantages, Problems, and Solutions. Biotechnol. Adv. 2019, 37, 746–770. [Google Scholar] [CrossRef] [PubMed]
- Manoel, E.A.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of Lipases on Hydrophobic Supports Involves the Open Form of the Enzyme. Enzym. Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef]
- Kim, K.K.; Song, H.K.; Shin, D.H.; Hwang, K.Y.; Suh, S.W. The Crystal Structure of a Triacylglycerol Lipase from Pseudomonas cepacia Reveals a Highly Open Conformation in the Absence of a Bound Inhibitor. Structure 1997, 5, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Cygler, M.; Schrag, J.D. Structure and Conformational Flexibility of Candida rugosa Lipase. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 1999, 1441, 205–214. [Google Scholar] [CrossRef]
- Jaeger, K.-E.; Ransac, S.; Koch, H.B.; Ferrato, F.; Dijkstra, B.W. Topological Characterization and Modeling of the 3D Structure of Lipase from Pseudomonas aeruginosa. FEBS Lett. 1993, 332, 143–149. [Google Scholar] [CrossRef]
- dos Santos, J.C.S.; Rueda, N.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Tuning the Catalytic Properties of Lipases Immobilized on Divinylsulfone Activated Agarose by Altering Its Nanoenvironment. Enzym. Microb. Technol. 2015, 77, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rueda, N.; dos Santos, J.C.S.; Torres, R.; Ortiz, C.; Barbosa, O.; Fernandez-Lafuente, R. Improved Performance of Lipases Immobilized on Heterofunctional Octyl-Glyoxyl Agarose Beads. RSC Adv. 2015, 5, 11212–11222. [Google Scholar] [CrossRef]
- Virgen-Ortíz, J.J.; Tacias-Pascacio, V.G.; Hirata, D.B.; Torrestiana-Sanchez, B.; Rosales-Quintero, A.; Fernandez-Lafuente, R. Relevance of Substrates and Products on the Desorption of Lipases Physically Adsorbed on Hydrophobic Supports. Enzym. Microb. Technol. 2017, 96, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Diamanti, E.; Arana-Peña, S.; Ramos-Cabrer, P.; Comino, N.; Carballares, D.; Fernandez-Lafuente, R.; López-Gallego, F. Intraparticle Macromolecular Migration Alters the Structure and Function of Proteins Reversibly Immobilized on Porous Microbeads. Adv. Mater. Interfaces 2022, 9, 2200263. [Google Scholar] [CrossRef]
- Arana-Peña, S.; Rios, N.S.; Carballares, D.; Mendez-Sanchez, C.; Lokha, Y.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Effects of Enzyme Loading and Immobilization Conditions on the Catalytic Features of Lipase from Pseudomonas fluorescens Immobilized on Octyl-Agarose Beads. Front. Bioeng. Biotechnol. 2020, 8, 36. [Google Scholar] [CrossRef]
- Zaak, H.; Siar, E.-H.; Kornecki, J.F.; Fernandez-Lopez, L.; Pedrero, S.G.; Virgen-Ortíz, J.J.; Fernandez-Lafuente, R. Effect of Immobilization Rate and Enzyme Crowding on Enzyme Stability under Different Conditions. The Case of Lipase from Thermomyces lanuginosus Immobilized on Octyl Agarose Beads. Process Biochem. 2017, 56, 117–123. [Google Scholar] [CrossRef]
- Fernandez-Lopez, L.; Pedrero, S.G.; Lopez-Carrobles, N.; Gorines, B.C.; Virgen-Ortíz, J.J.; Fernandez-Lafuente, R. Effect of Protein Load on Stability of Immobilized Enzymes. Enzym. Microb. Technol. 2017, 98, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lopez, L.; Bartolome-Cabrero, R.; Rodriguez, M.D.; Dos Santos, C.S.; Rueda, N.; Fernandez-Lafuente, R. Stabilizing Effects of Cations on Lipases Depend on the Immobilization Protocol. RSC Adv. 2015, 5, 83868–83875. [Google Scholar] [CrossRef]
- Kornecki, J.F.; Carballares, D.; Morellon-Sterling, R.; Siar, E.H.; Kashefi, S.; Chafiaa, M.; Arana-Peña, S.; Rios, N.S.; Gonçalves, L.R.B.B.; Fernandez-Lafuente, R. Influence of Phosphate Anions on the Stability of Immobilized Enzymes. Effect of Enzyme Nature, Immobilization Protocol and Inactivation Conditions. Process Biochem. 2020, 95, 288–296. [Google Scholar] [CrossRef]
- Abellanas-Perez, P.; Carballares, D.; Fernandez-Lafuente, R.; Rocha-Martin, J. Glutaraldehyde Modification of Lipases Immobilized on Octyl Agarose Beads: Roles of the Support Enzyme Loading and Chemical Amination of the Enzyme on the Final Enzyme Features. Int. J. Biol. Macromol. 2023, 248, 125853. [Google Scholar] [CrossRef] [PubMed]
- Abellanas-Perez, P.; Carballares, D.; Rocha-Martin, J.; Fernandez-Lafuente, R. The Nature of the Buffer Alters the Effects of the Chemical Modification on the Stability of Immobilized Lipases. Process Biochem. 2023, 133, 20–27. [Google Scholar] [CrossRef]
- Abellanas-Perez, P.; Carballares, D.; Rocha-Martin, J.; Fernandez-Lafuente, R. The Effects of the Chemical Modification on Immobilized Lipase Features Are Affected by the Enzyme Crowding in the Support. Biotechnol. Prog. 2023, 12, e3394. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and Its Derivatives as Supports for Enzyme Immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef] [PubMed]
- Mangiagalli, M.; Ami, D.; Divitiis, M.; Brocca, S.; Catelani, T.; Natalello, A.; Lotti, M. Short-chain Alcohols Inactivate an Immobilized Industrial Lipase through Two Different Mechanisms. Biotechnol. J. 2022, 17, 2100712. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R. Lipase from Thermomyces lanuginosus: Uses and Prospects as an Industrial Biocatalyst. J. Mol. Catal. B Enzym. 2010, 62, 197–212. [Google Scholar] [CrossRef]
- Brzozowski, A.M.; Savage, H.; Verma, C.S.; Turkenburg, J.P.; Lawson, D.M.; Svendsen, A.; Patkar, S. Structural Origins of the Interfacial Activation in Thermomyces (Humicola) lanuginosa Lipase. Biochemistry 2000, 39, 15071–15082. [Google Scholar] [CrossRef]
- Cajal, Y.; Svendsen, A.; Girona, V.; Patkar, S.A.; Alsina, M.A. Interfacial Control of Lid Opening in Thermomyces Lanuginosa Lipase. Biochemistry 2000, 39, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Larsson, K.M.; Kirk, O. One Biocatalyst—Many Applications: The Use of Candida antarctica B-Lipase in Organic Synthesis. Biocatal. Biotransform. 1998, 16, 181–204. [Google Scholar] [CrossRef]
- Kirk, O.; Christensen, M.W. Lipases from Candida antarctica: Unique Biocatalysts from a Unique Origin. Org. Process Res. Dev. 2002, 6, 446–451. [Google Scholar] [CrossRef]
- Martinelle, M.; Holmquist, M.; Hult, K. On the Interfacial Activation of Candida antarctica Lipase A and B as Compared with Humicola lanuginosa Lipase. Biochim. Et Biophys. Acta-Lipids Lipid Metab. 1995, 1258, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Volpato, G.; Filice, M.; Ayub, M.A.Z.; Guisan, J.M.; Palomo, J.M. Single-Step Purification of Different Lipases from Staphylococcus Warneri. J. Chromatogr. A 2010, 1217, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lorente, G.; Palomo, J.M.; Fuentes, M.; Mateo, C.; Guisán, J.M.; Fernández-Lafuente, R. Self-Assembly of Pseudomonas fluorescens Lipase into Bimolecular Aggregates Dramatically Affects Functional Properties. Biotechnol. Bioeng. 2003, 82, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Palomo, J.M.; Peñas, M.M.; Fernández-Lorente, G.; Mateo, C.; Pisabarro, A.G.; Fernández-Lafuente, R.; Ramírez, L.; Guisán, J.M. Solid-Phase Handling of Hydrophobins: Immobilized Hydrophobins as a New Tool to Study Lipases. Biomacromolecules 2003, 4, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; He, J.; Sun, Y.; Reynolds, M.; Zhang, L.; Han, S.; Liang, S.; Sui, H.; Lin, Y. Display of Fungal Hydrophobin on the Pichia pastoris Cell Surface and Its Influence on Candida antarctica Lipase B. Appl. Microbiol. Biotechnol. 2016, 100, 5883–5895. [Google Scholar] [CrossRef]
- Zhang, K.; Jin, Z.; Wang, P.; Zheng, S.P.; Han, S.Y.; Lin, Y. Improving the Catalytic Characteristics of Lipase-Displaying Yeast Cells by Hydrophobic Modification. Bioprocess. Biosyst. Eng. 2017, 40, 1689–1699. [Google Scholar] [CrossRef]
- Ribitsch, D.; Guebitz, G.M. Tuning of Adsorption of Enzymes to Polymer. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2021; pp. 293–315. [Google Scholar]
- Rocha-Pino, Z.; Ramos-López, J.I.; Gimeno, M.; Barragán-Aroche, F.; Durán-Valencia, C.; López-Ramírez, S.; Shirai, K. Enhanced Oil Recovery by Hydrophobins from Lecanicillium lecanii. Fuel 2018, 224, 10–16. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lombardo, D.; Guy, O. Effect of Alcohols on the Hydrolysis Catalyzed by Human Pancreatic Carboxylic-Ester Hydrolase. Biochim. Et Biophys. Acta (BBA)-Enzymol. 1981, 657, 425–437. [Google Scholar] [CrossRef]
- Hernandez, K.; Garcia-Verdugo, E.; Porcar, R.; Fernandez-Lafuente, R. Hydrolysis of Triacetin Catalyzed by Immobilized Lipases: Effect of the Immobilization Protocol and Experimental Conditions on Diacetin Yield. Enzym. Microb. Technol. 2011, 48, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters Necessary to Define an Immobilized Enzyme Preparation. Process Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biocatalyst | Activity (U/g) with 50 mM Triacetin | Activity (U/g) with 10 mM (R)-Methyl Mandelate | Activity (U/g) with 10 mM (S)-Methyl Mandelate |
|---|---|---|---|
| Octyl-TLL (1 mg/g) Phosphate | 18.16 ± 0.91 | Under detection limit | Under detection limit |
| Octyl-TLL (1 mg/g) Tris-HCL | 17.04 ± 0.68 | Under detection limit | Under detection limit |
| Octyl-TLL (1 mg/g) Hepes | 24.18 ± 1.21 | Under detection limit | Under detection limit |
| Octyl-TLL (40 mg/g) Phosphate | 162.04 ± 6.43 | 0.045 ± 0.002 | 0.033 ± 0.001 |
| Octyl-TLL (40 mg/g) Tris-HCL | 187.10 ± 9.31 | 0.035 ± 0.001 | 0.040 ± 0.002 |
| Octyl-TLL (40 mg/g) Hepes | 165.67 ± 8.22 | 0.038 ± 0.001 | 0.047 ± 0.002 |
| Octyl-CALB (1 mg/g) Phosphate | 2.61 ± 0.11 | 8.86 ± 0.35 | 0.523 ± 0.026 |
| Octyl-CALB (1 mg/g) Tris-HCL | 1.76 ± 0.08 | 6.85 ± 0.27 | 0.509 ± 0.025 |
| Octyl-CALB (1 mg/g) Hepes | 1.87 ± 0.07 | 6.77 ± 0.33 | 0.516 ± 0.020 |
| Octyl-CALB (24 mg/g) Phosphate | 82.73 ± 3.30 | 148.69 ± 5.95 | 11.06 ± 0.44 |
| Octyl-CALB (24 mg/g) Tris-HCL | 62.89 ± 2.51 | 160.97 ± 8.13 | 15.22 ± 0.76 |
| Octyl-CALB (24 mg/g) Hepes | 51.03 ± 2.04 | 177.05 ± 8.81 | 12.75 ± 0.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abellanas-Perez, P.; Carballares, D.; Rocha-Martin, J.; Fernandez-Lafuente, R. The Effects of Buffer Nature on Immobilized Lipase Stability Depend on Enzyme Support Loading. Catalysts 2024, 14, 105. https://doi.org/10.3390/catal14020105
Abellanas-Perez P, Carballares D, Rocha-Martin J, Fernandez-Lafuente R. The Effects of Buffer Nature on Immobilized Lipase Stability Depend on Enzyme Support Loading. Catalysts. 2024; 14(2):105. https://doi.org/10.3390/catal14020105
Chicago/Turabian StyleAbellanas-Perez, Pedro, Diego Carballares, Javier Rocha-Martin, and Roberto Fernandez-Lafuente. 2024. "The Effects of Buffer Nature on Immobilized Lipase Stability Depend on Enzyme Support Loading" Catalysts 14, no. 2: 105. https://doi.org/10.3390/catal14020105