

Bioprocessing and Screening of Indigenous Wastes for Hyper Production of Fungal Lipase
 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Strain Identification and Culturing
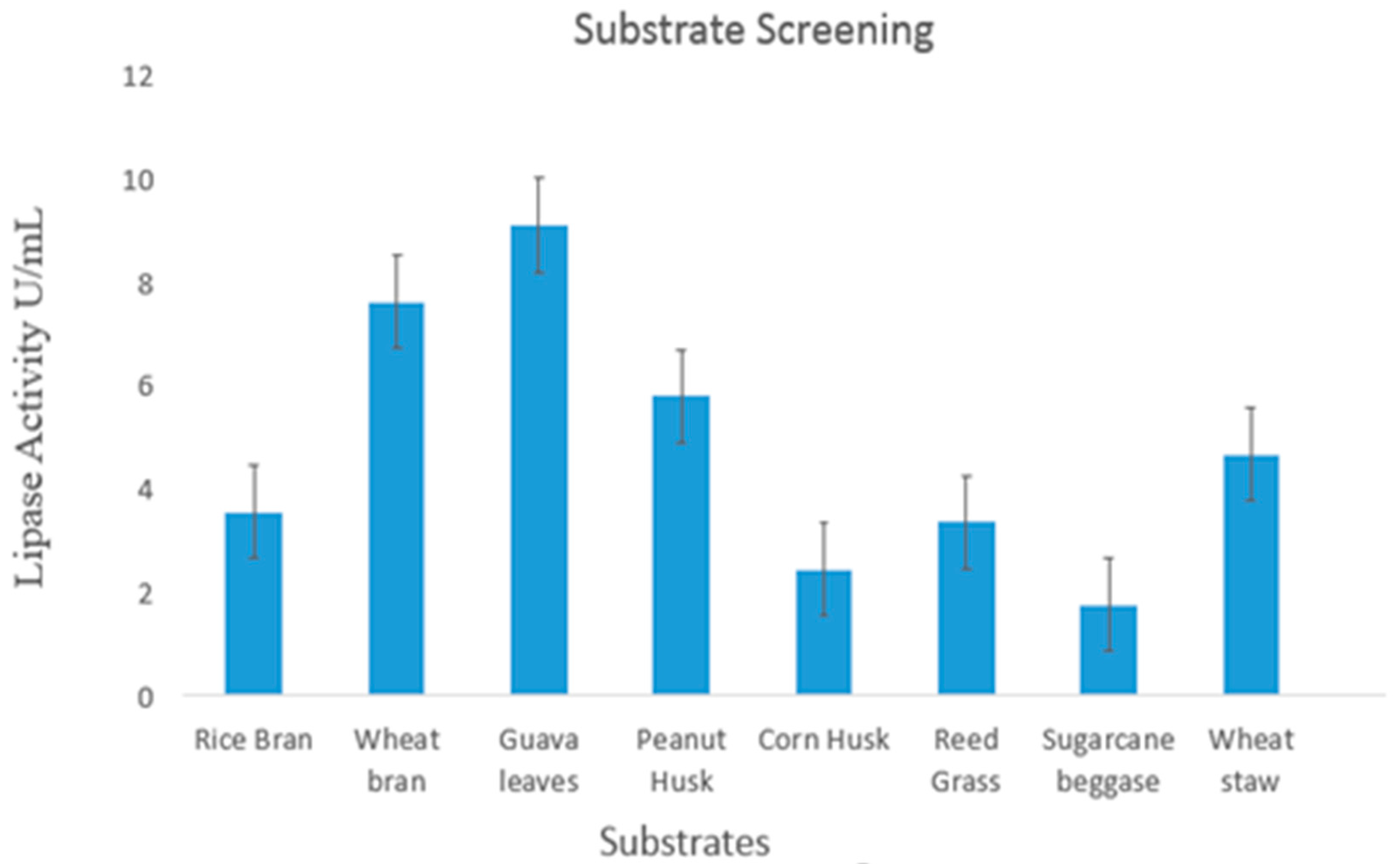
2.2. Substrate Screening
2.3. Optimization of Different Physical Parameters through Response Surface Methodology (RSM)
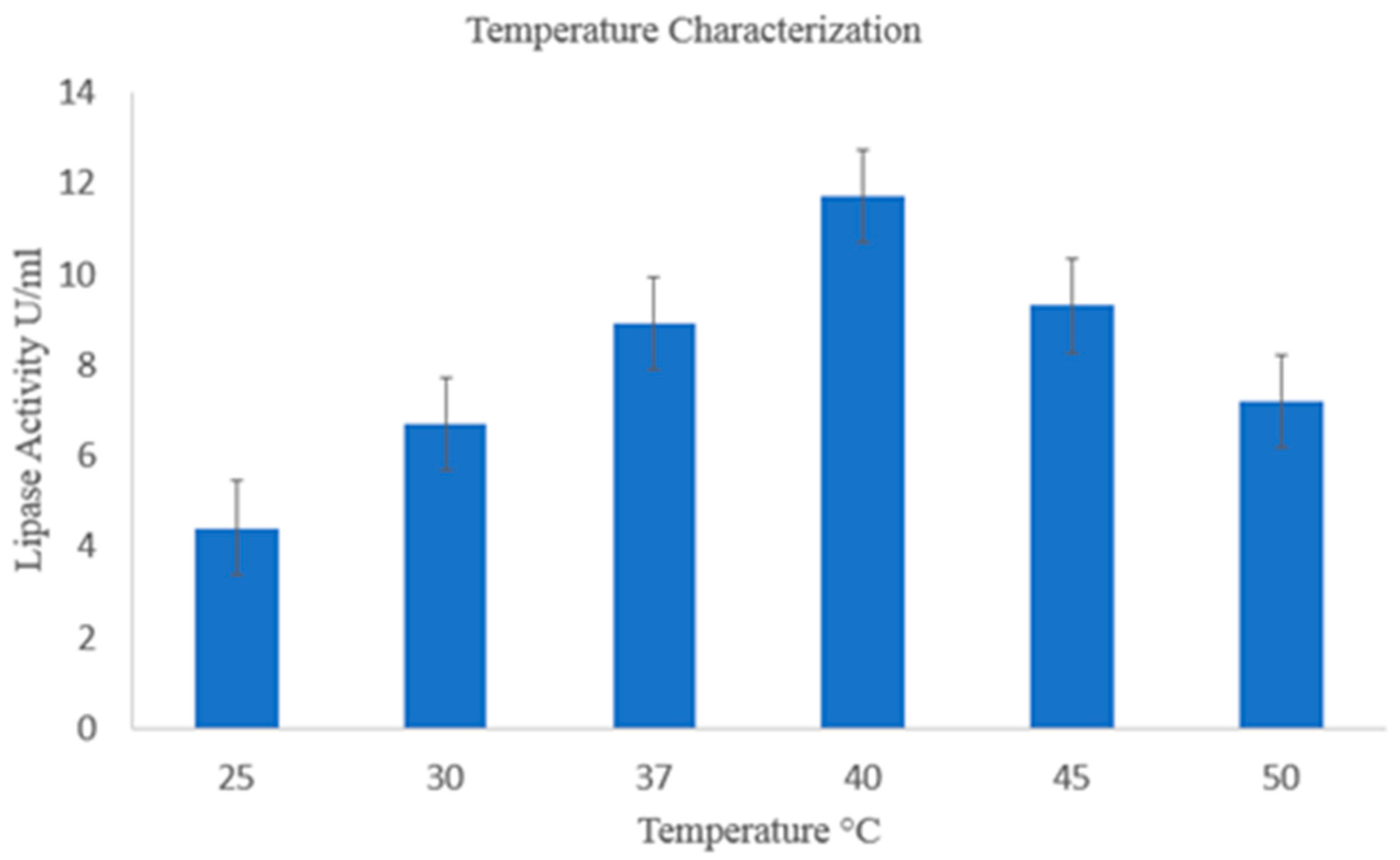
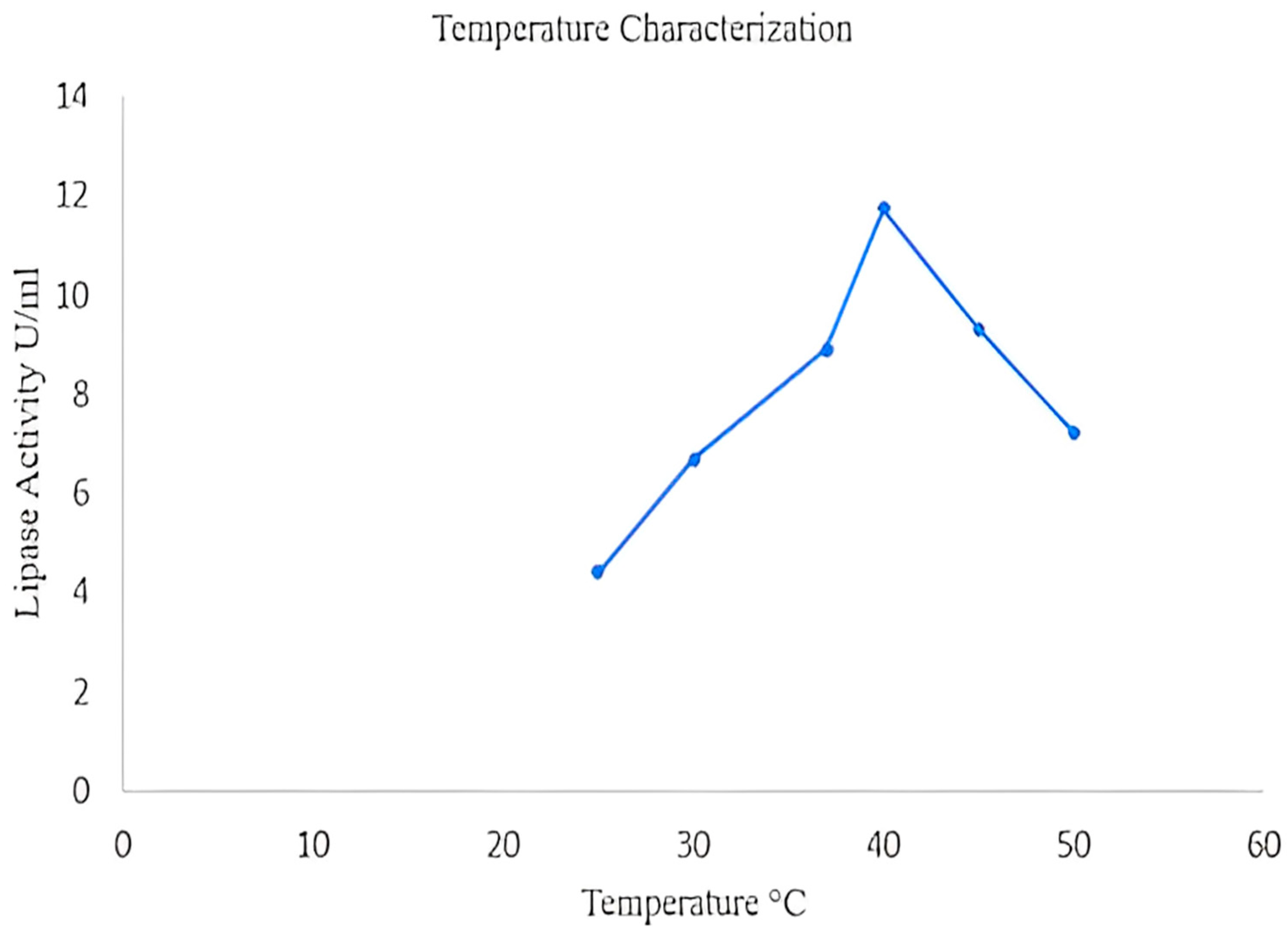
2.3.1. Temperature Optimization of Lipase
2.3.2. pH Optimization of Lipase
2.3.3. Optimization of Incubation Period for Lipase
2.3.4. Inoculum Size Optimization for Lipase Production
2.3.5. Optimization of Substrate Concentration for Lipase
2.4. Statistical Graphs for Lipase (3D and Contour)
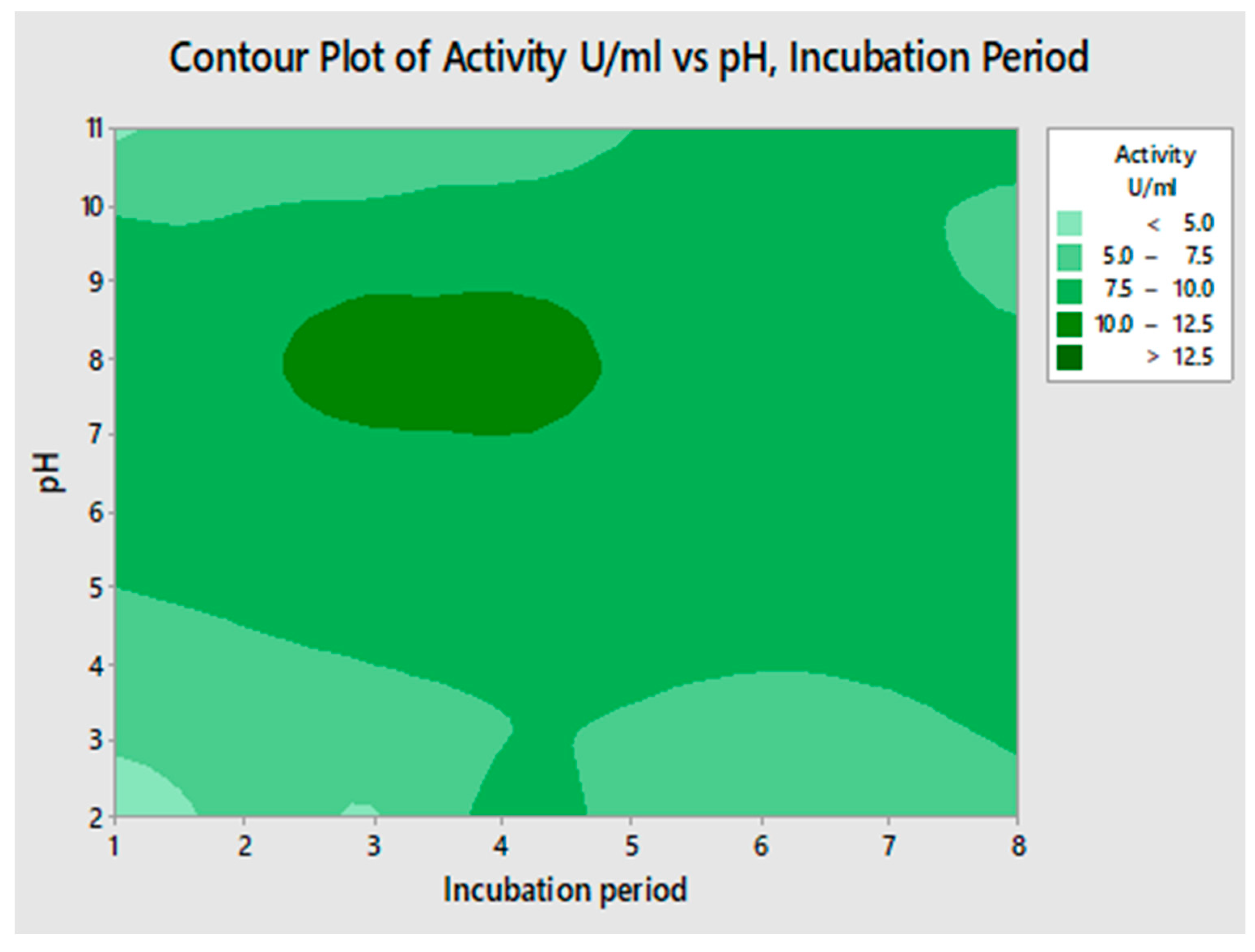
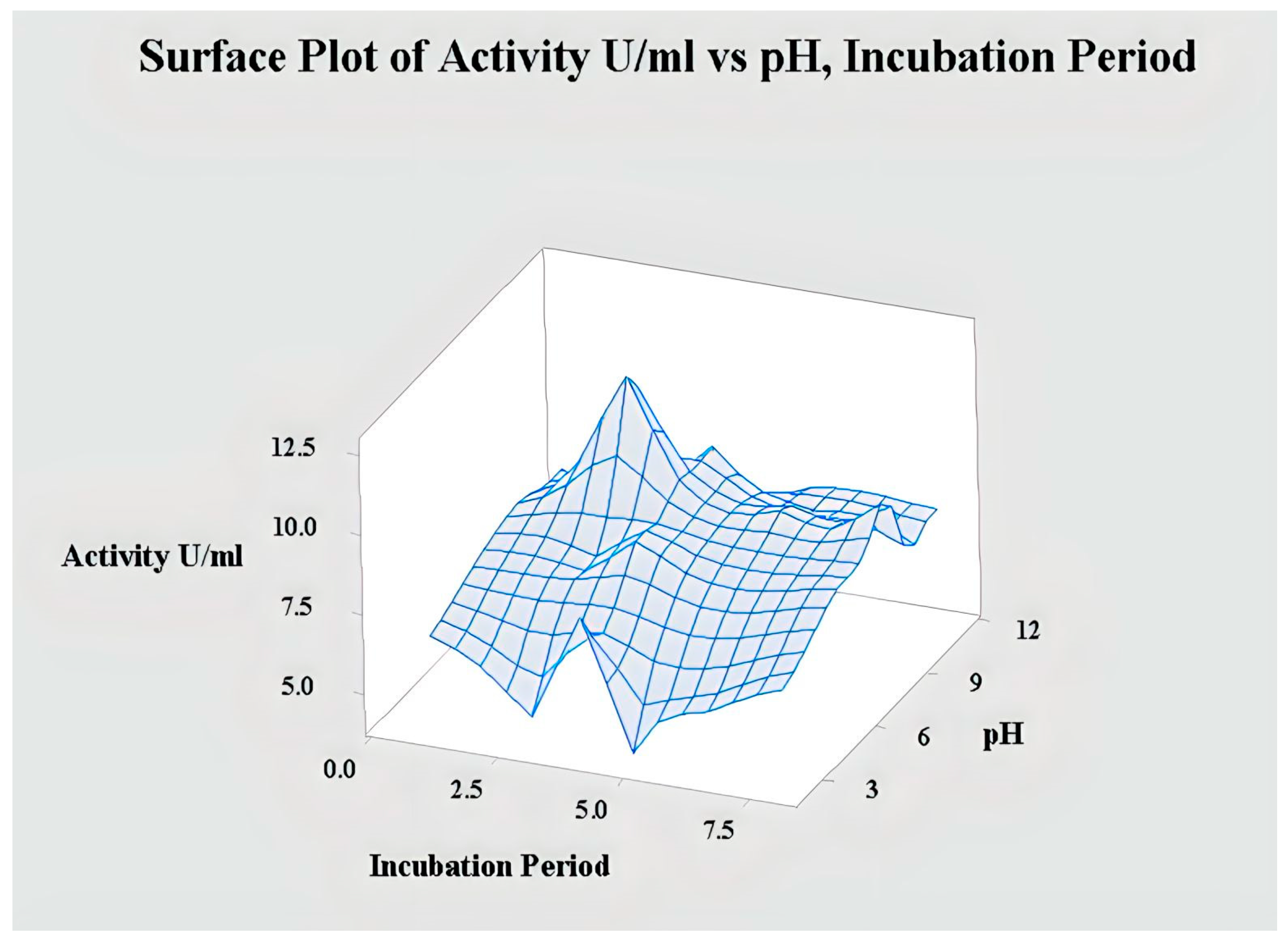
2.4.1. Interaction of pH and Incubation Period for Lipase Activity
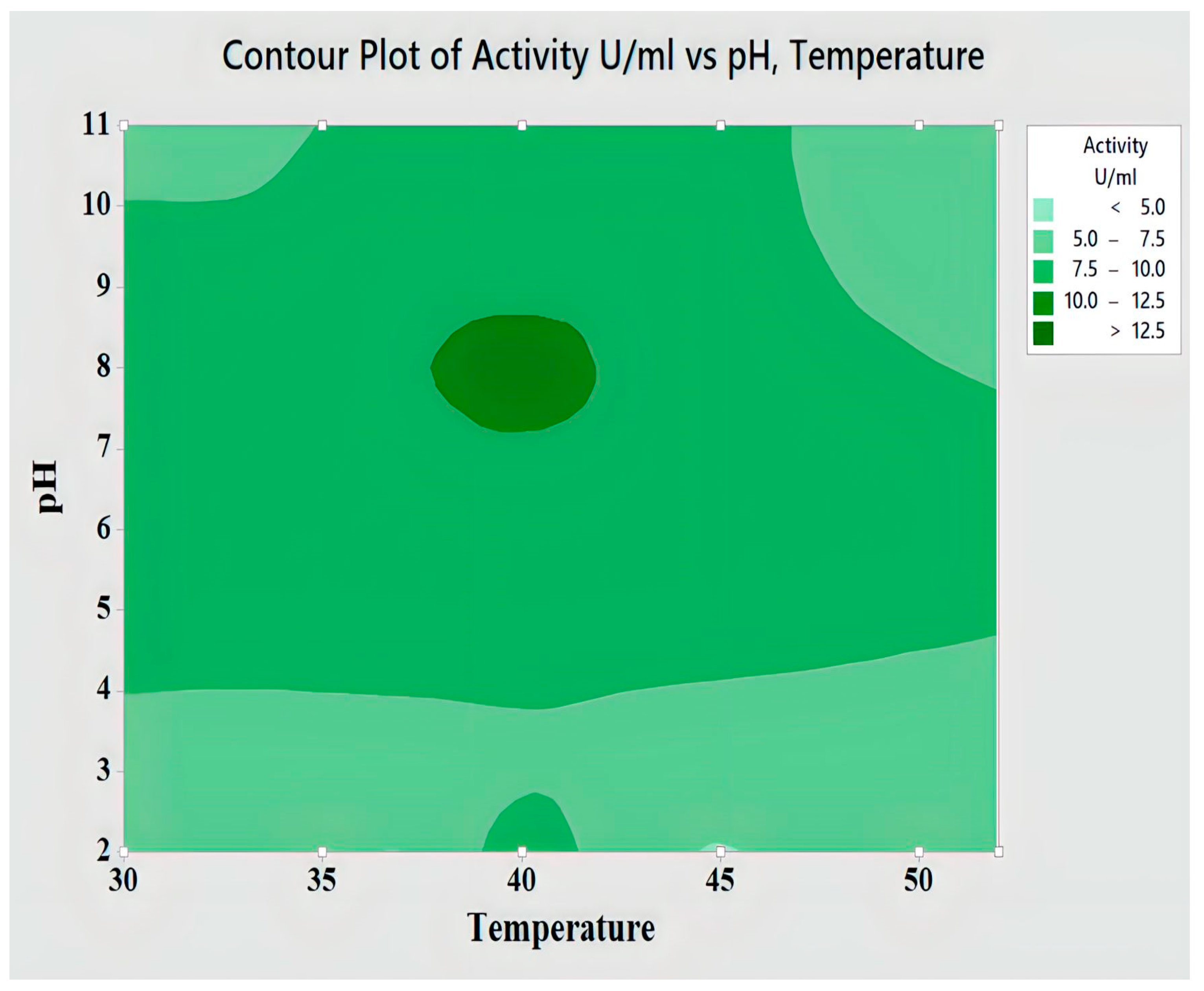
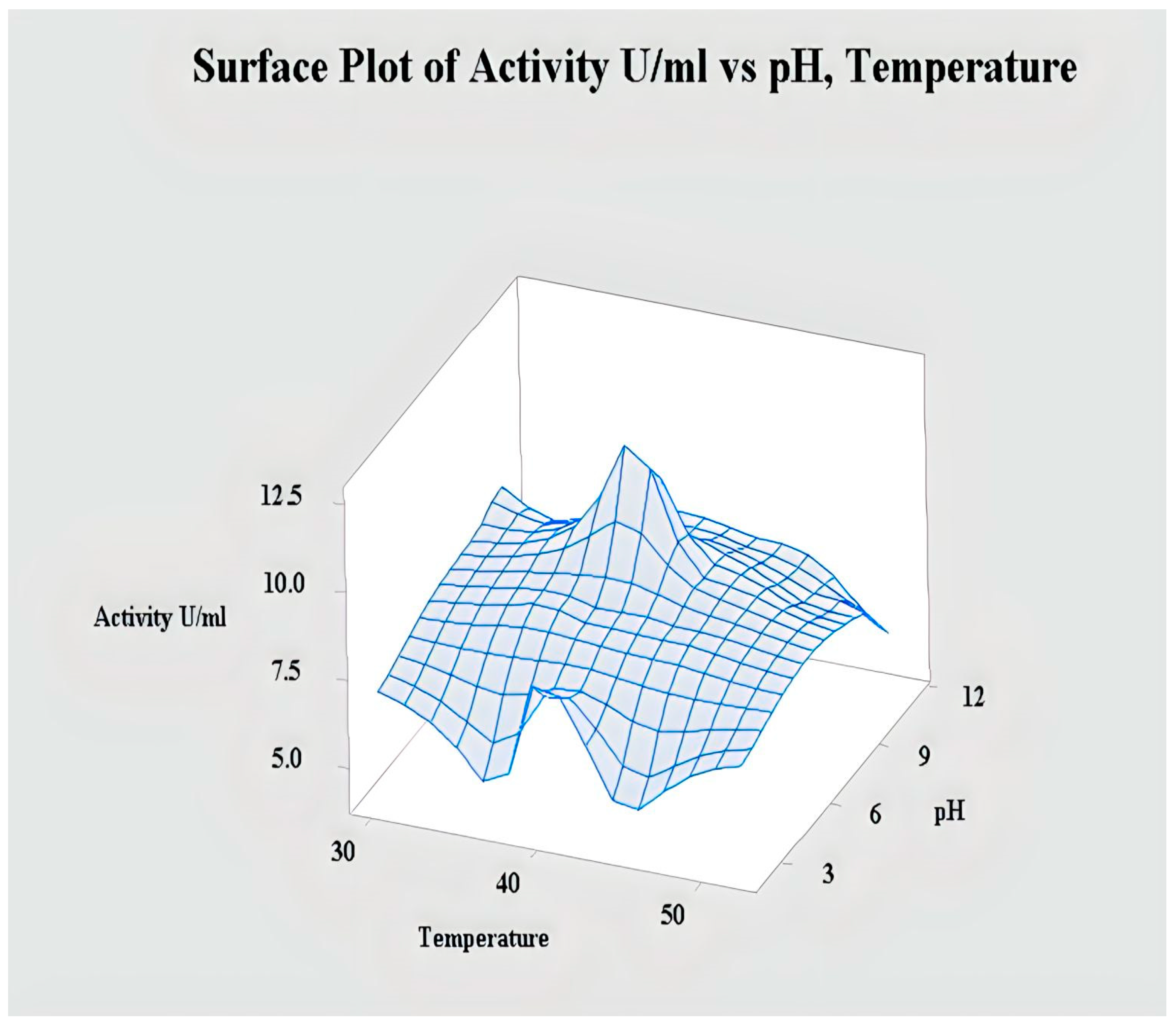
2.4.2. Interaction of pH and Temperature for Lipase Activity
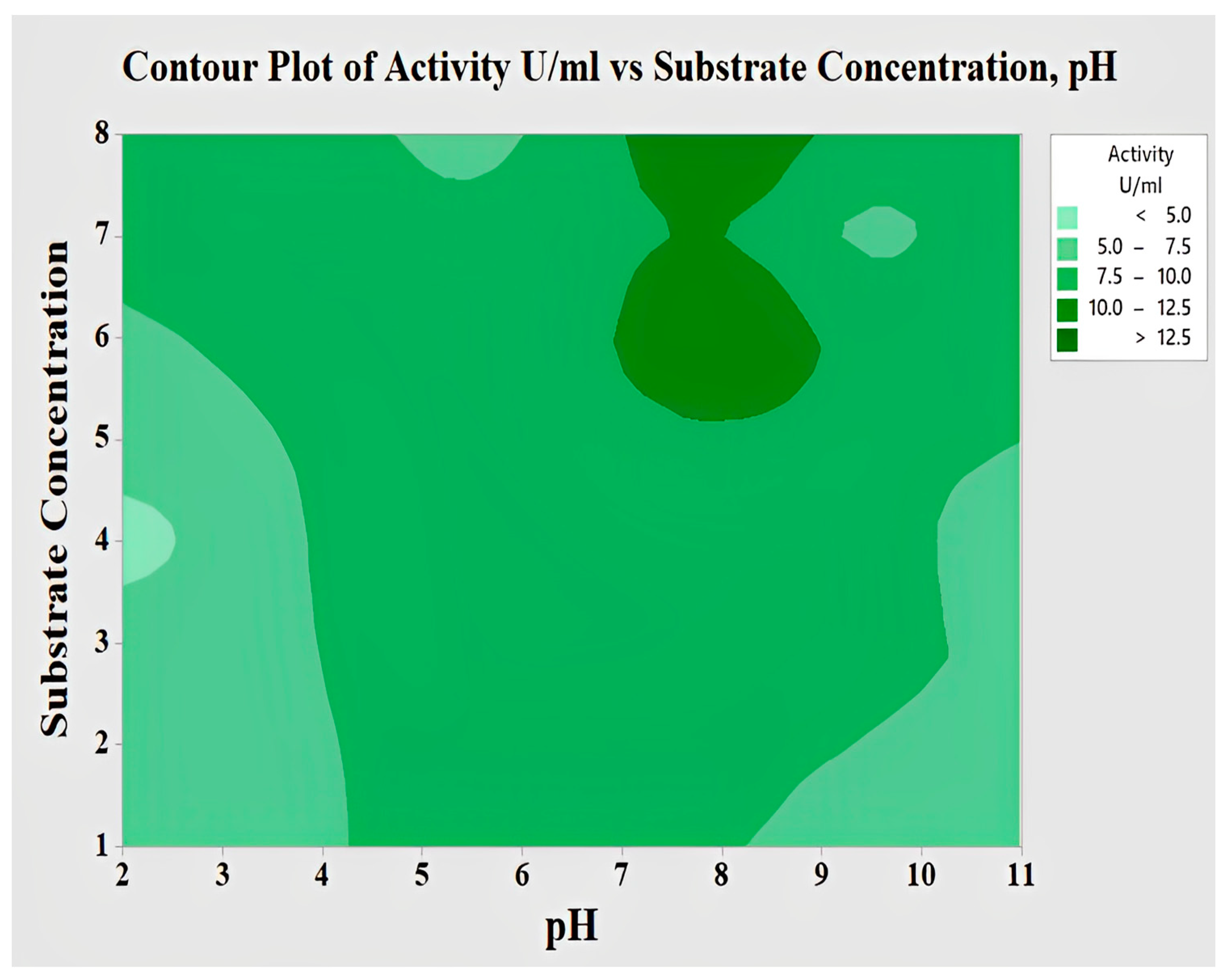
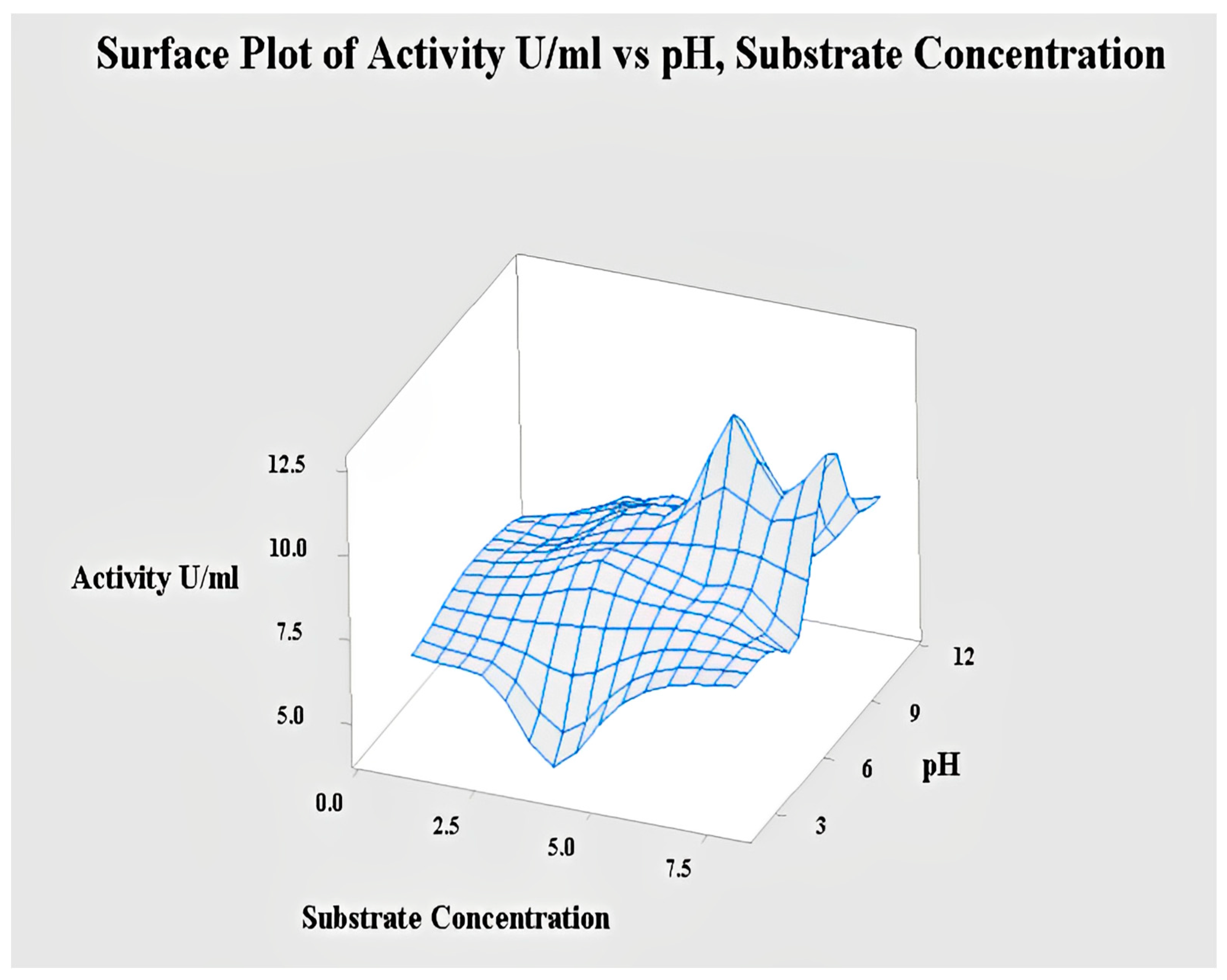
2.4.3. Interaction of pH and Substrate Concentration for Lipase
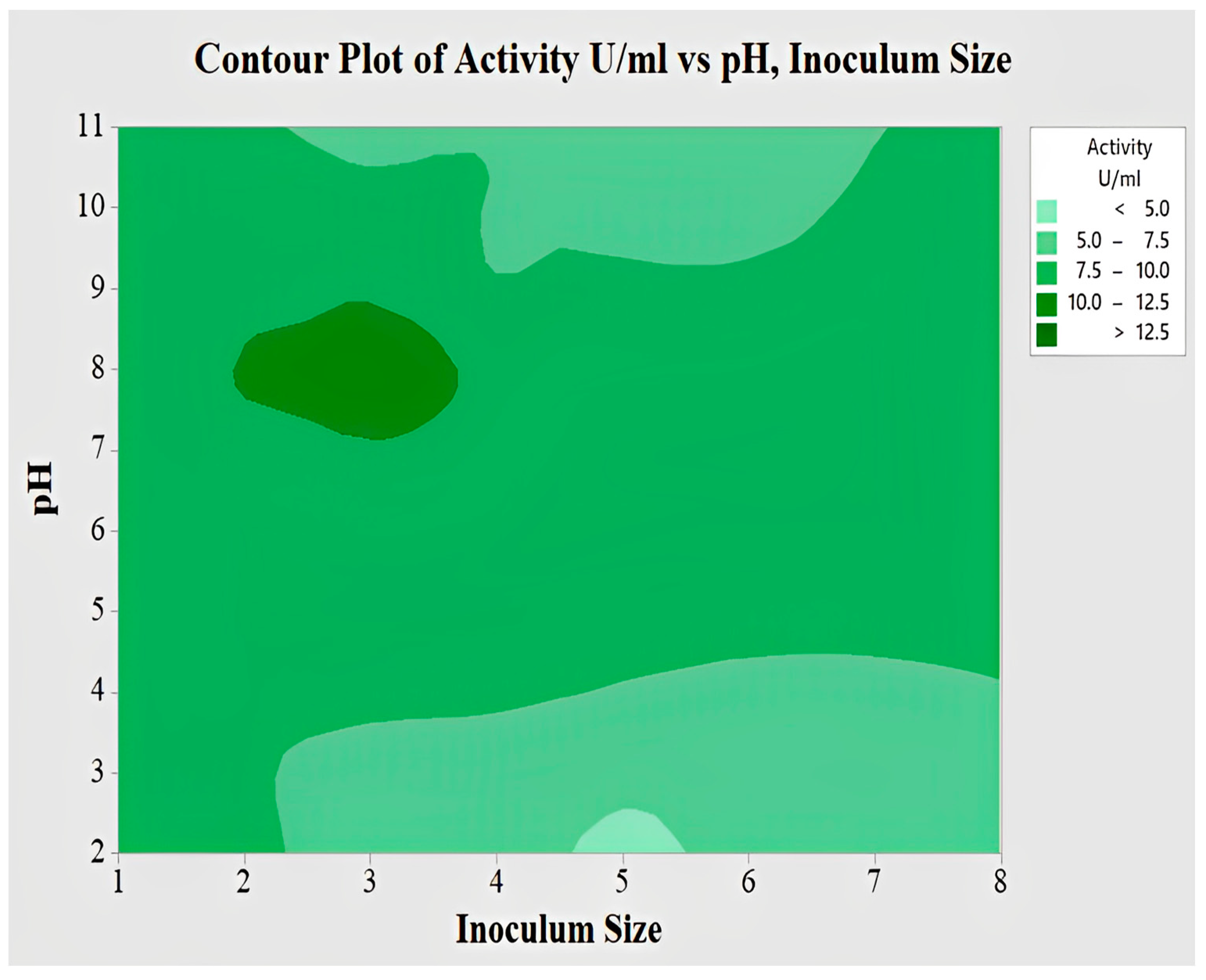
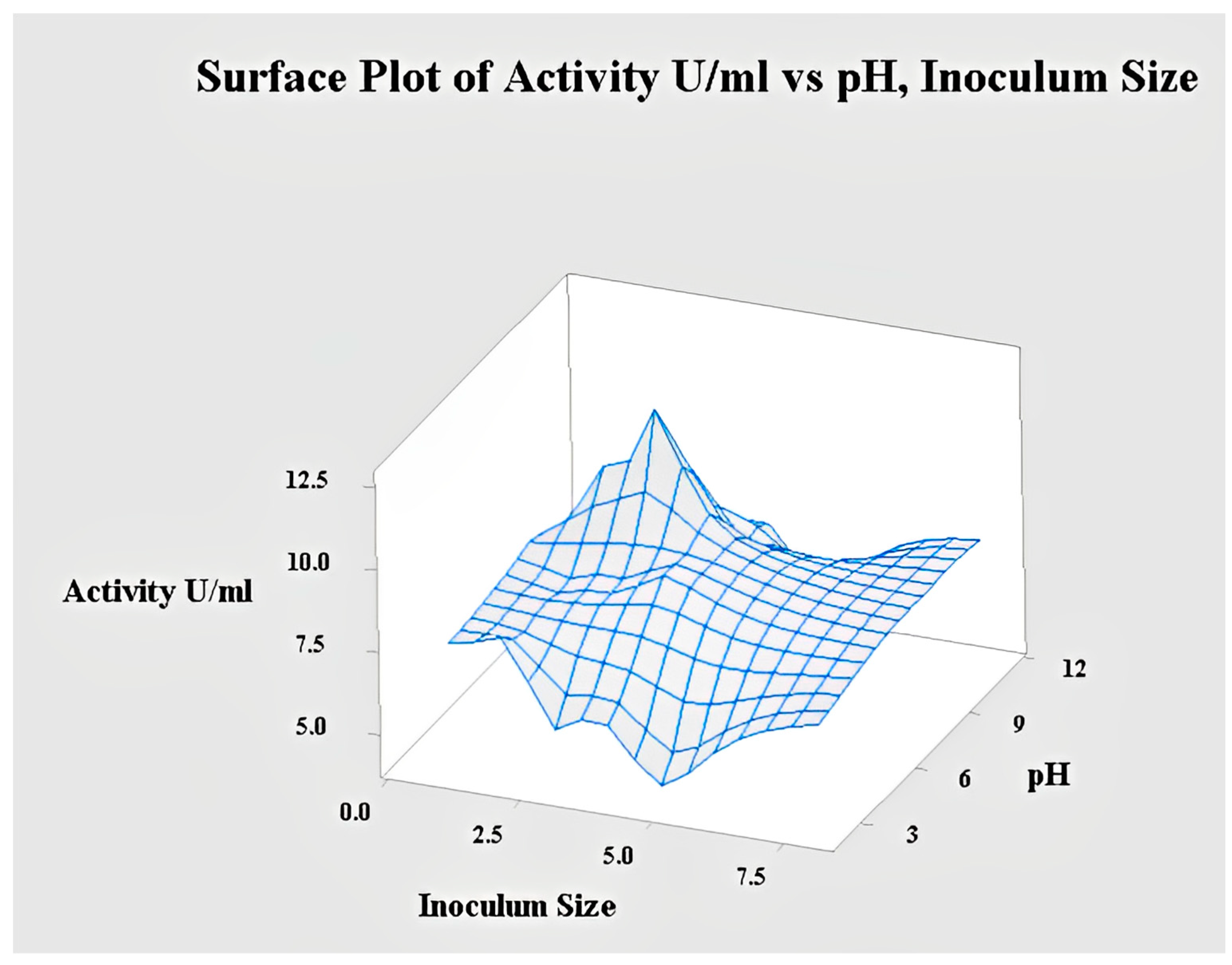
2.4.4. Interaction of pH and Inoculum Size for Lipase
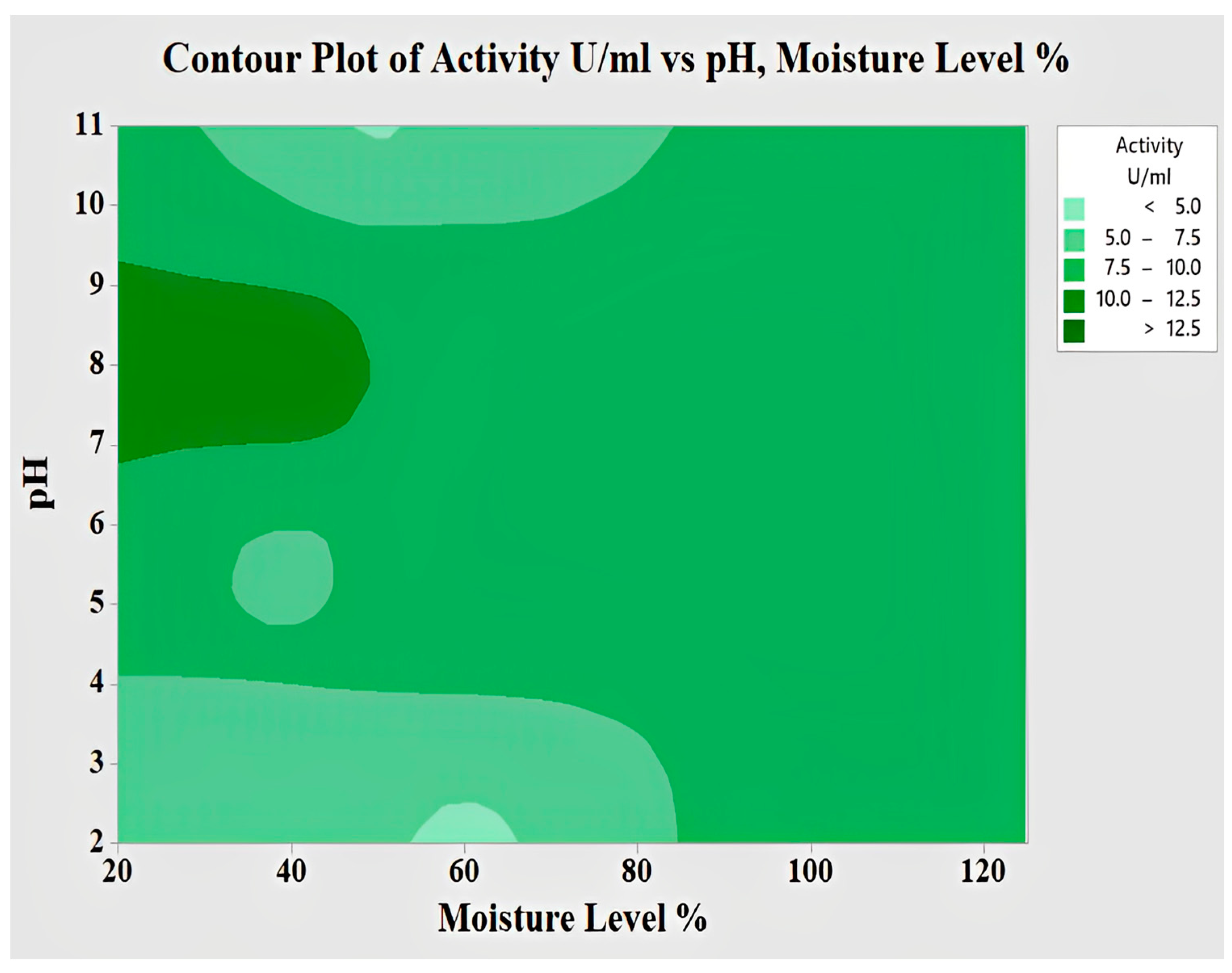
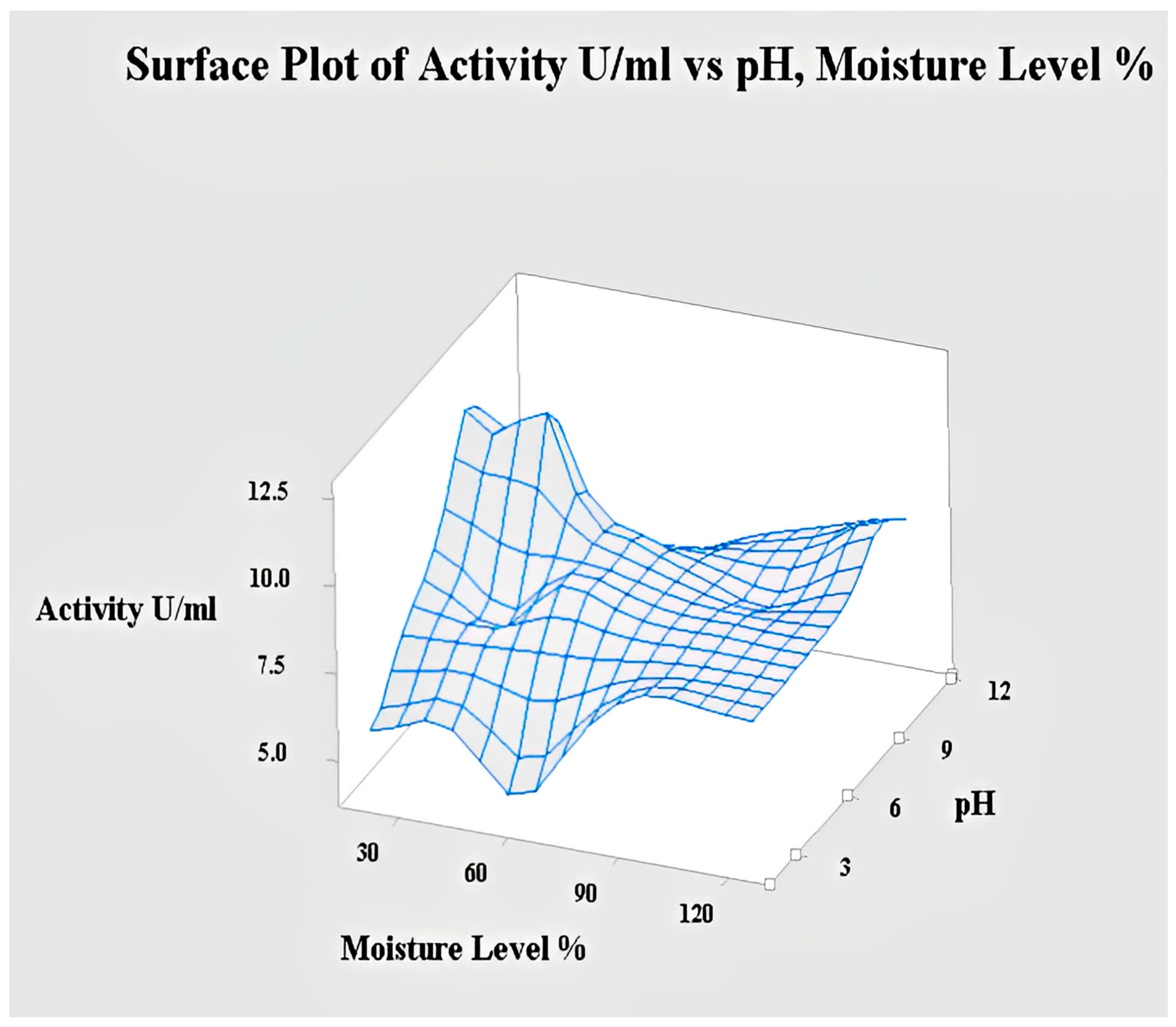
2.4.5. Interaction of pH and Moisture Content for Lipase Activity
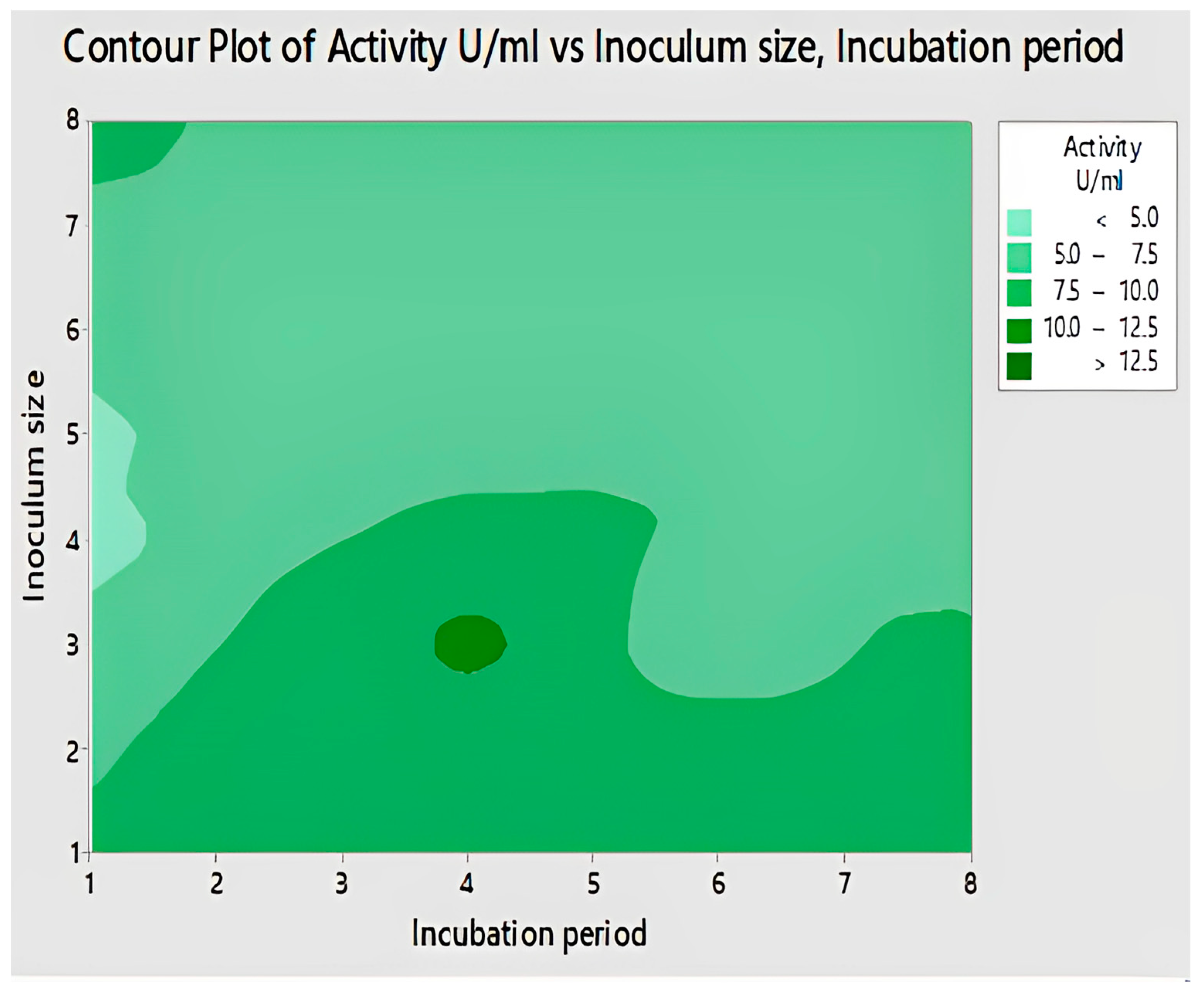
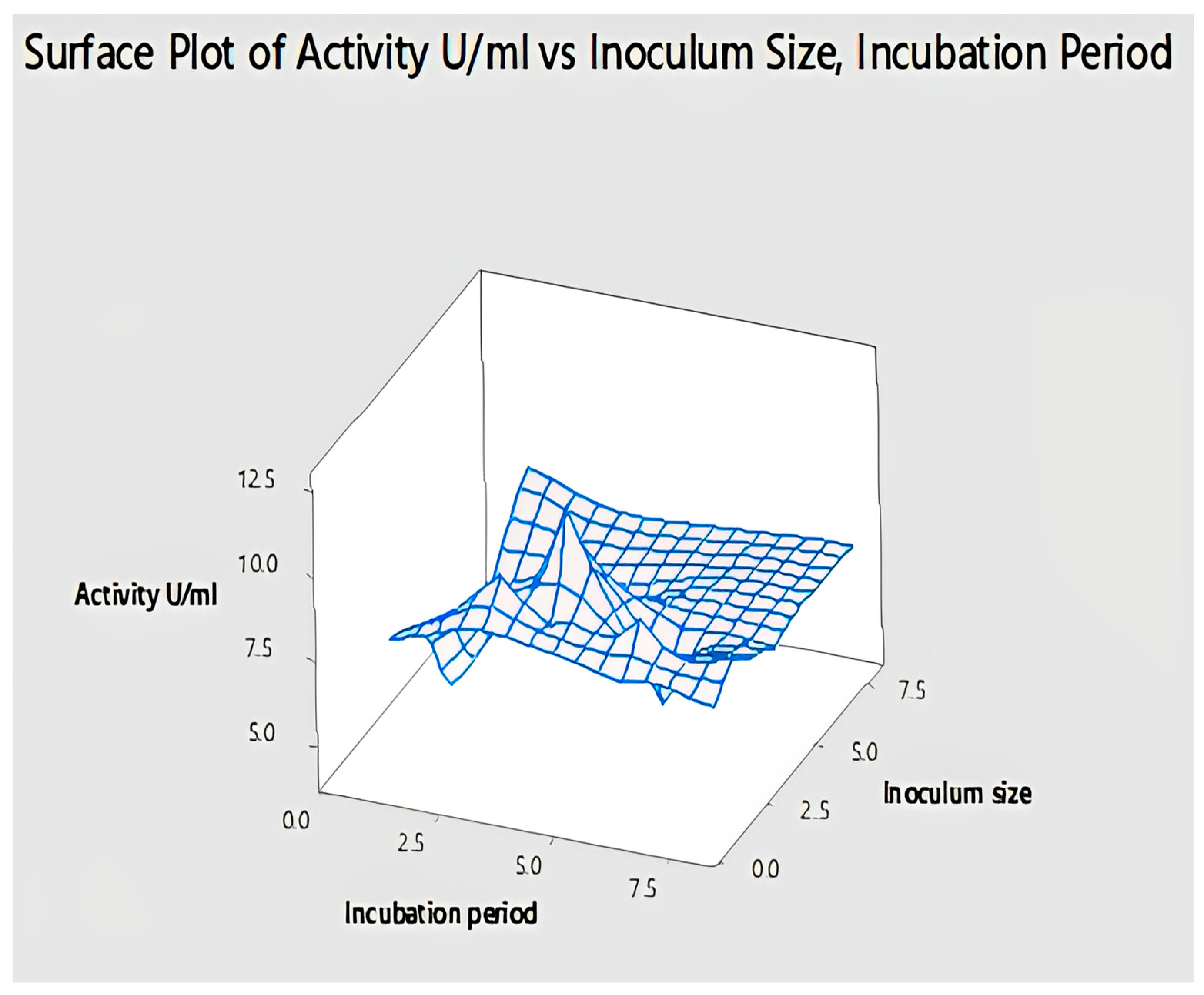
2.4.6. Interaction of Inoculum Size and Incubation Period for Lipase
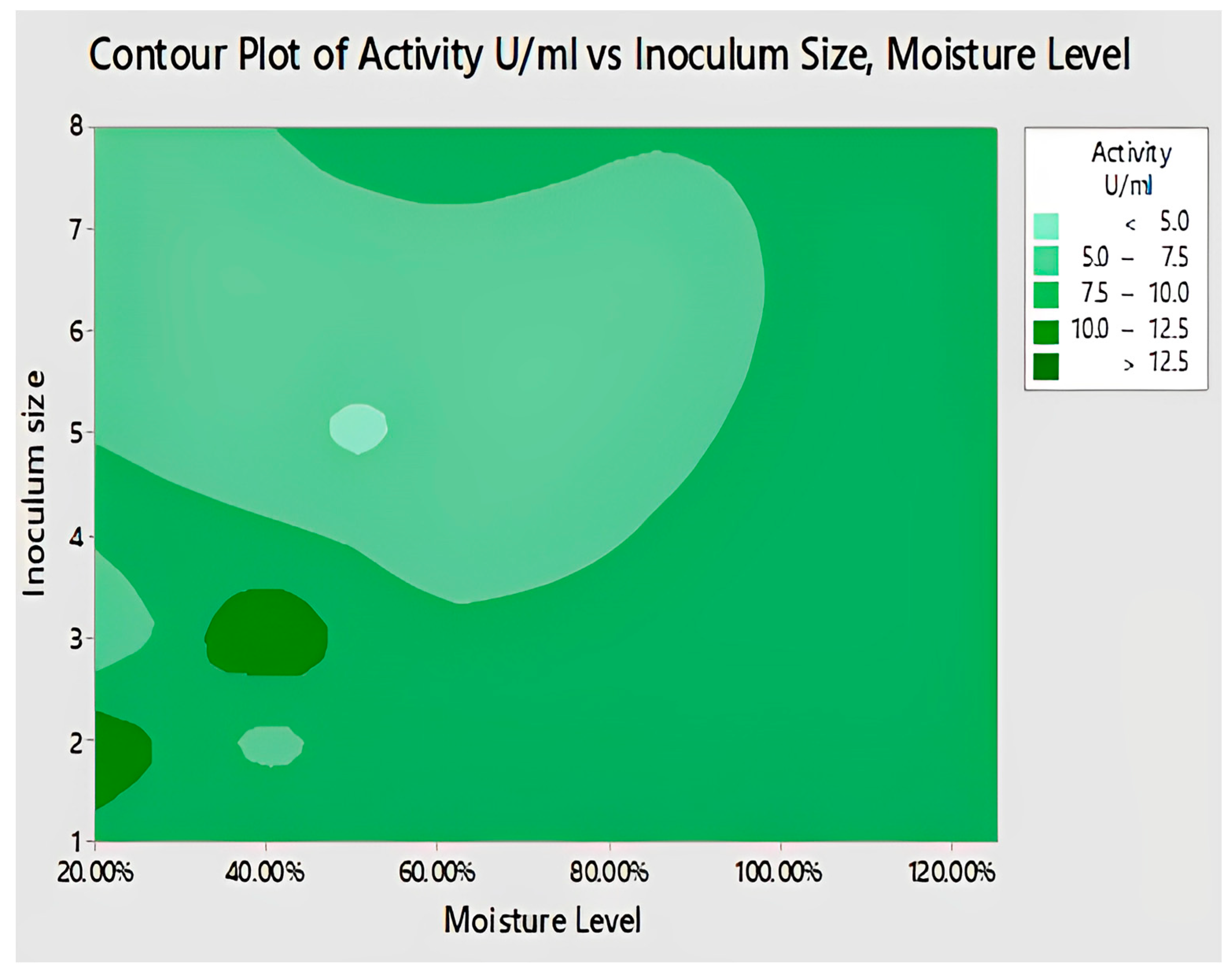
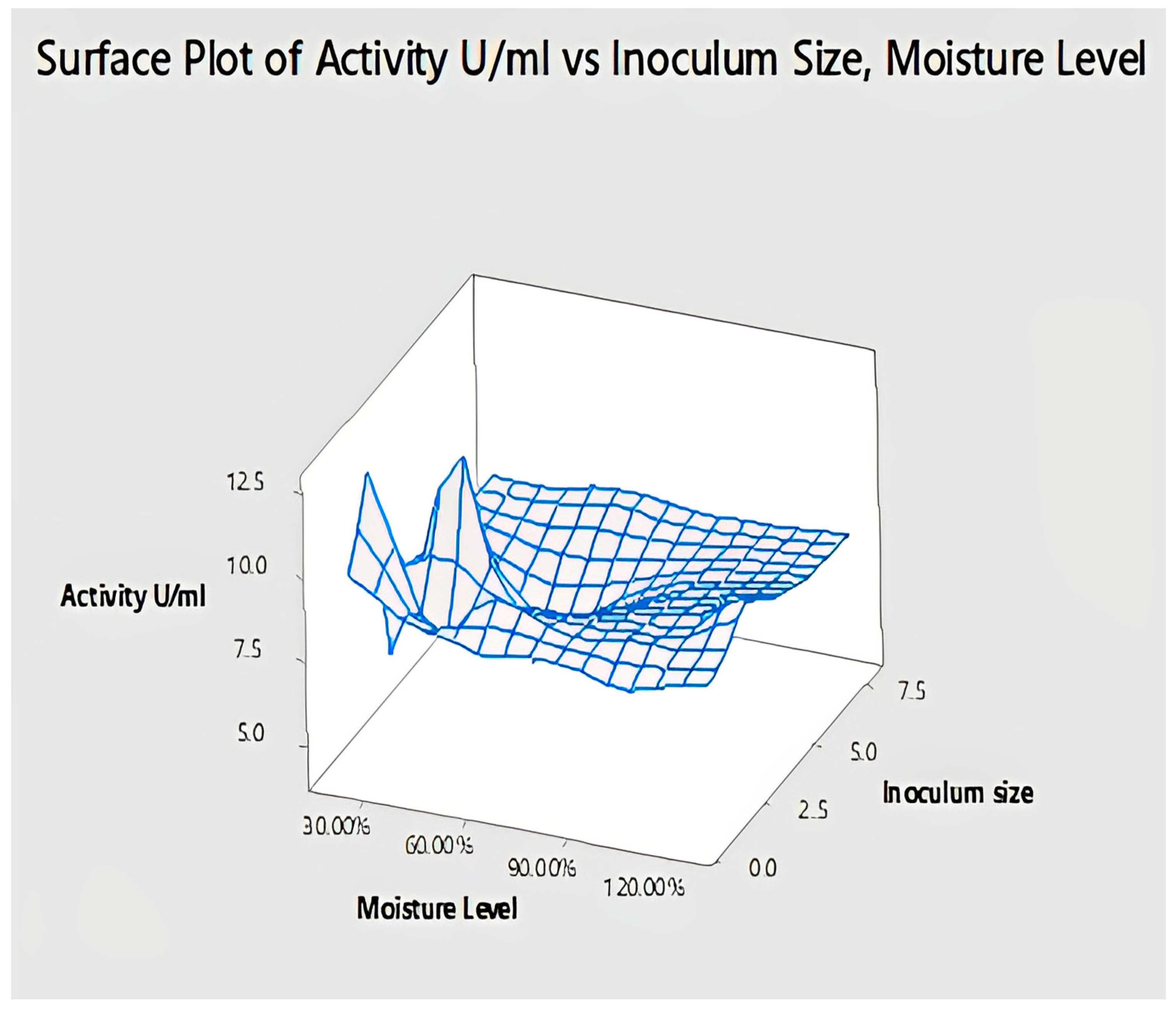
2.4.7. Interaction of Inoculum Size and Moisture Level for Lipase Activity
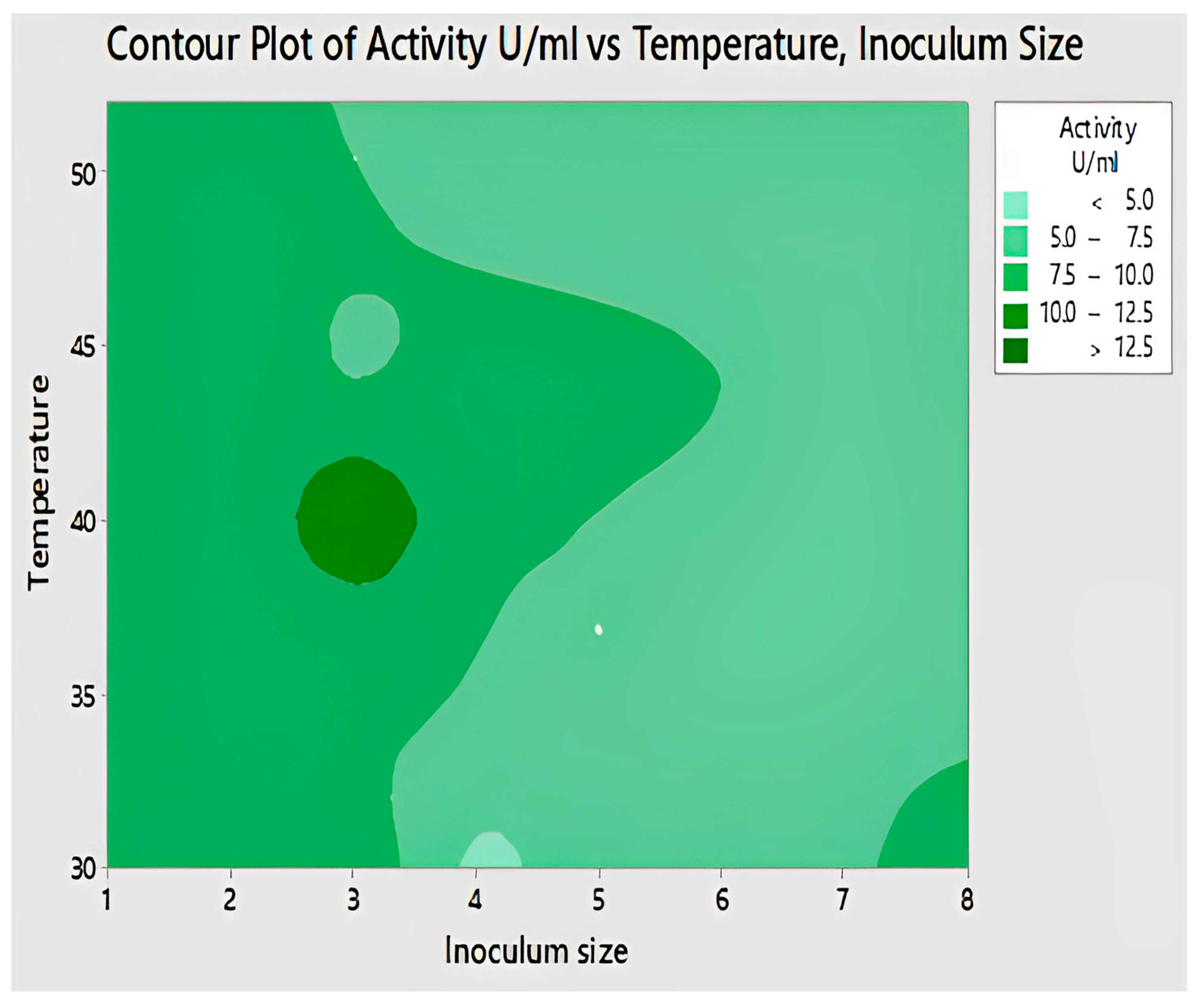
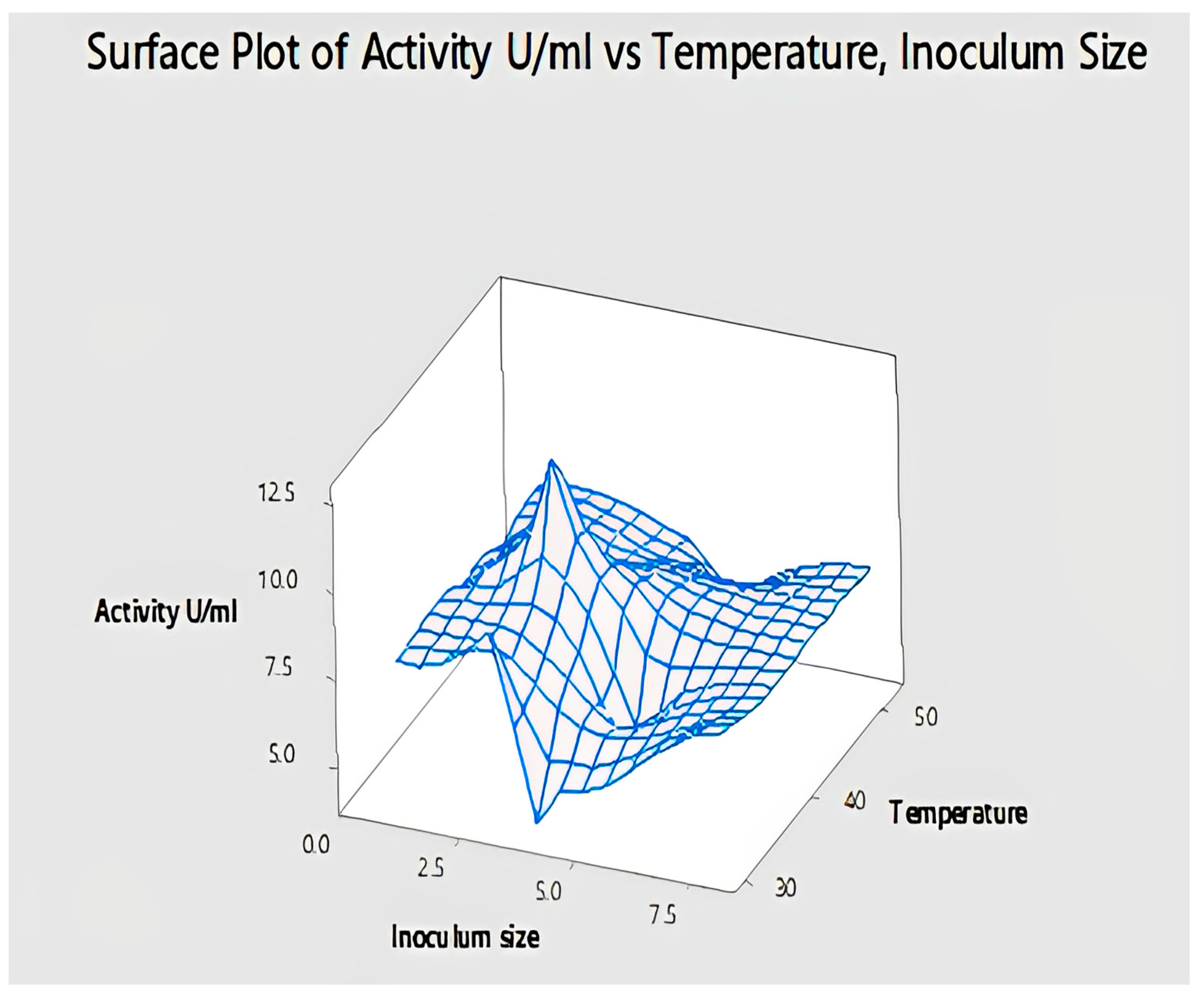
2.4.8. Interaction of Temperature and Inoculum Size for Lipase
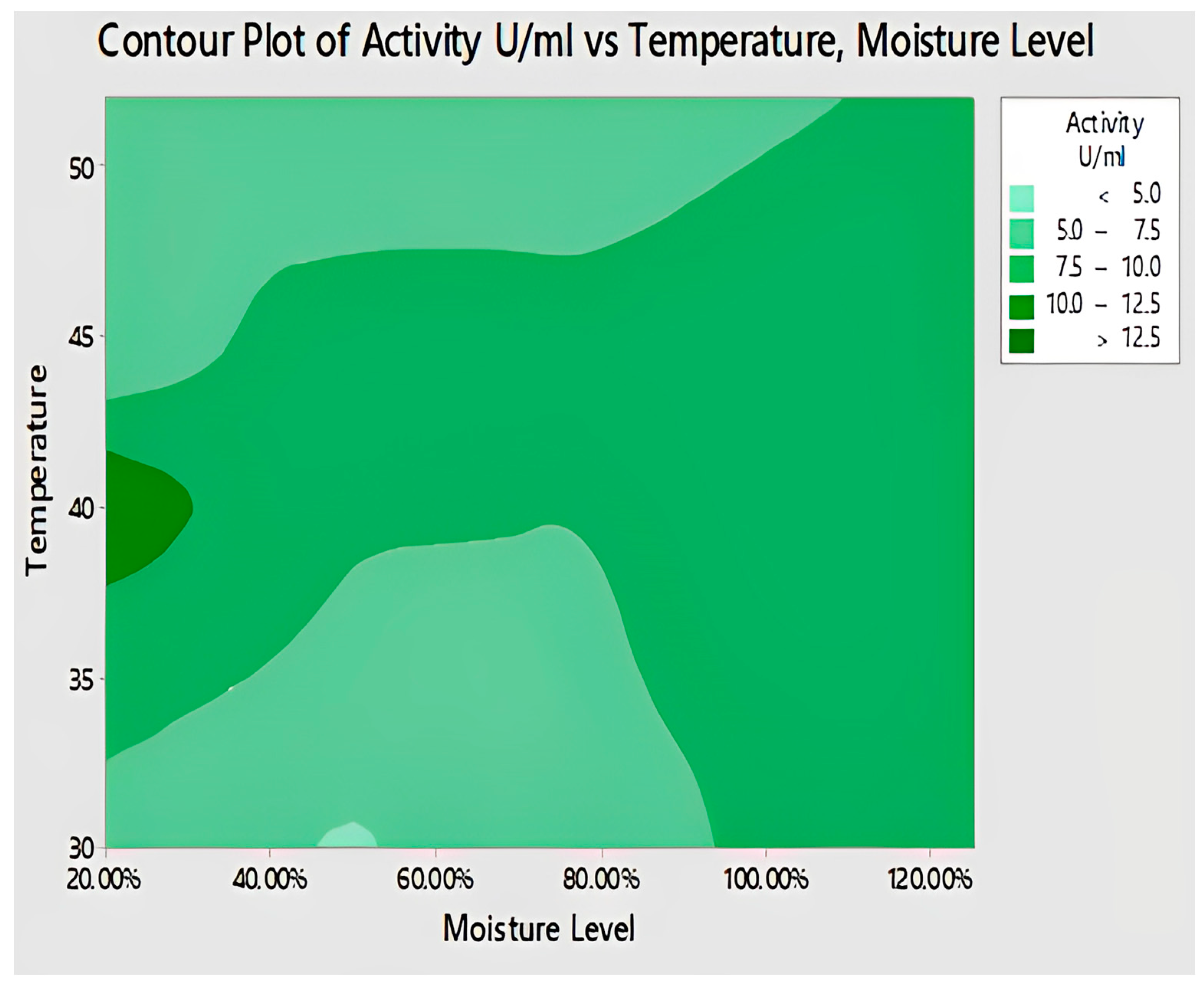
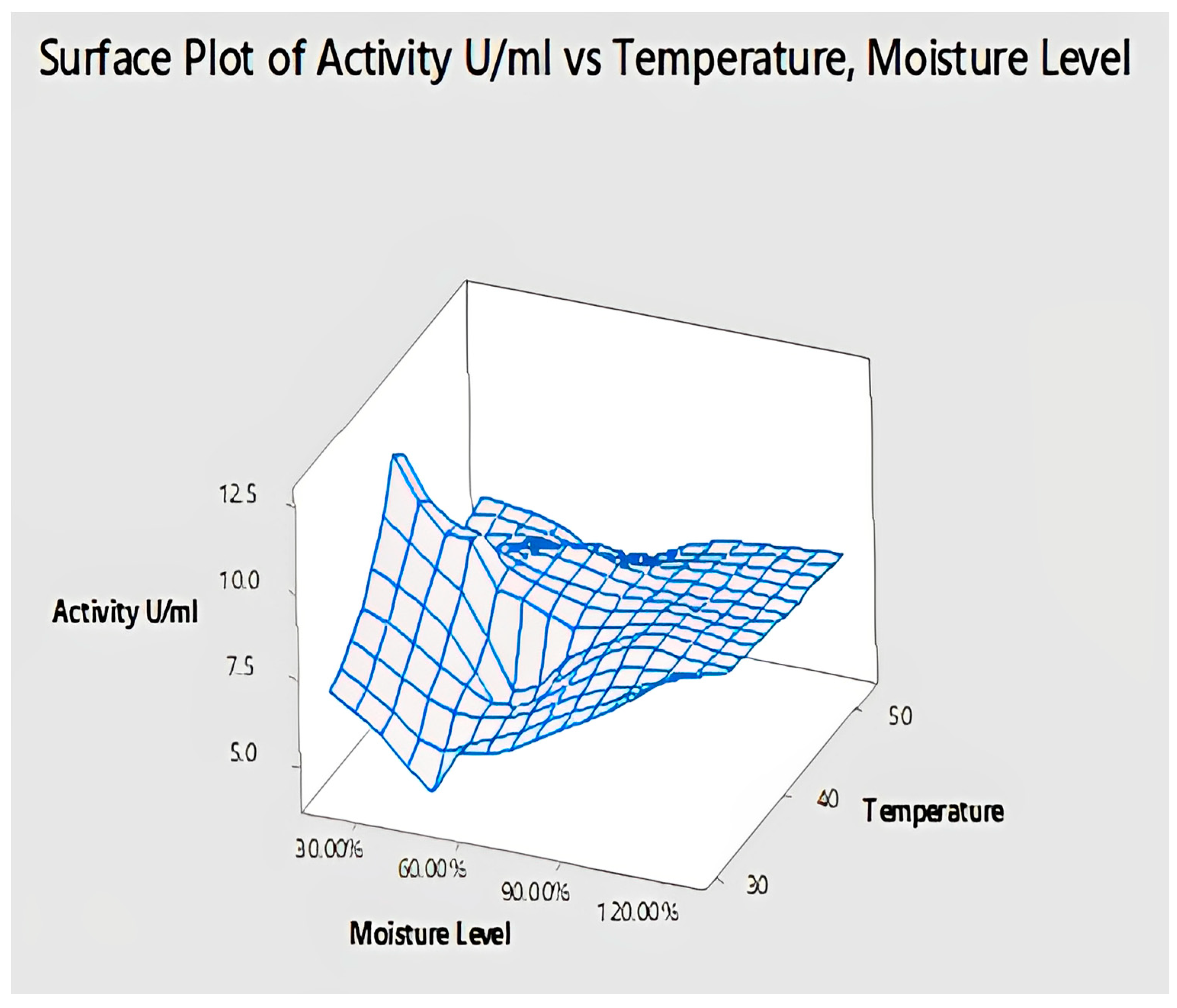
2.4.9. Interaction of Temperature and Moisture Level for Lipase Activity
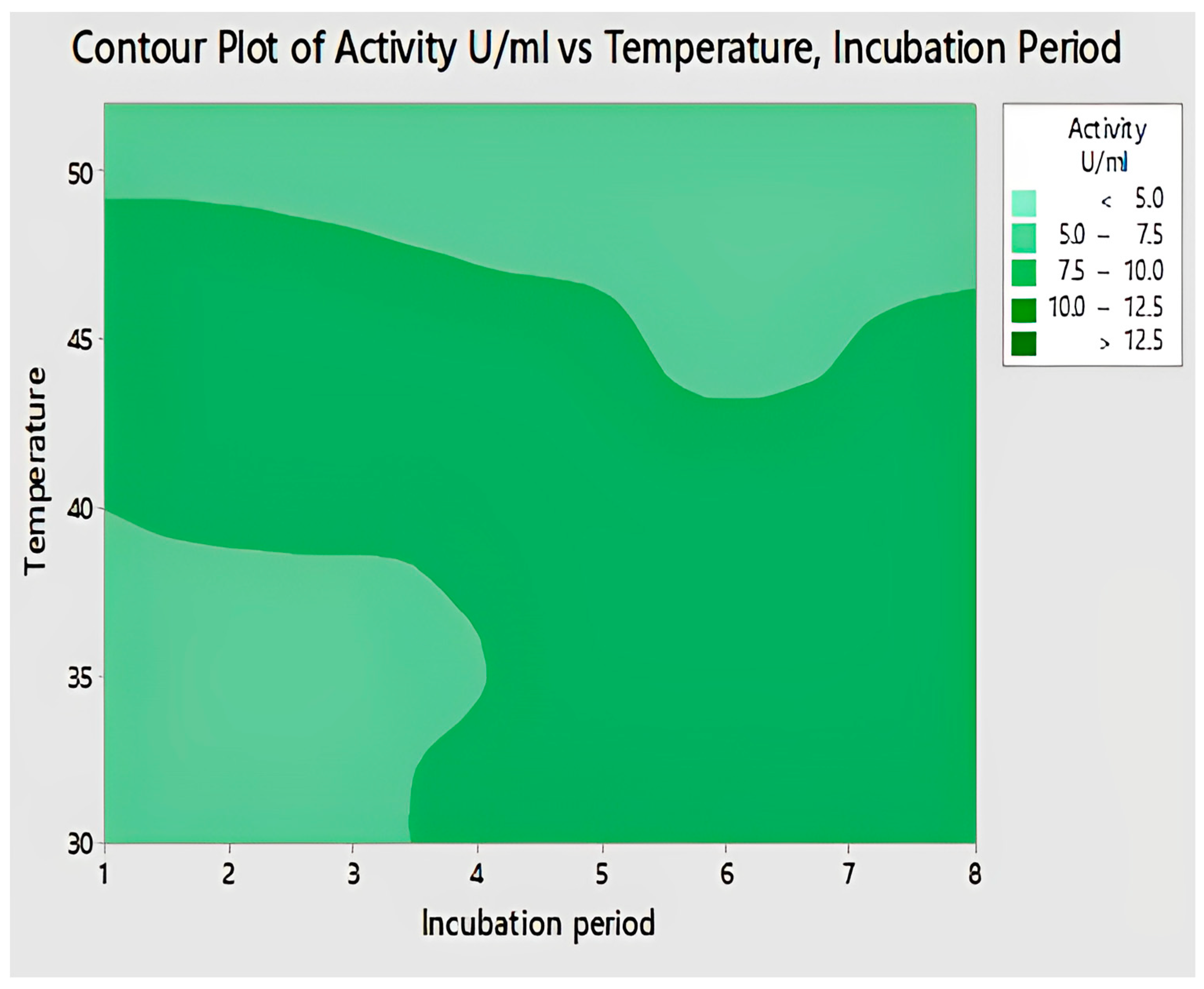
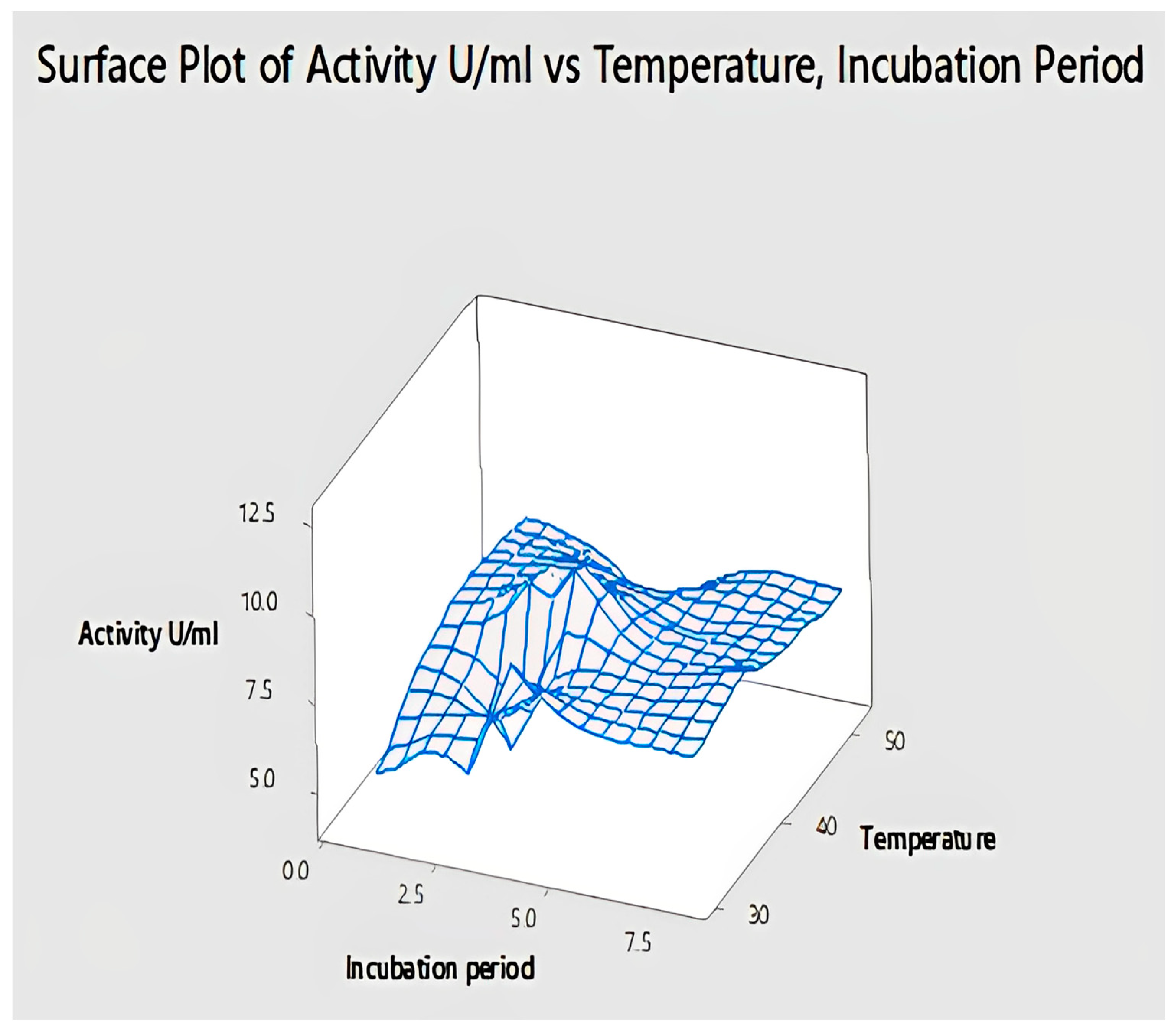
2.4.10. Interaction of Temperature and Incubation Period for Lipase Activity
2.5. Interpretations of Regression Surface Analysis
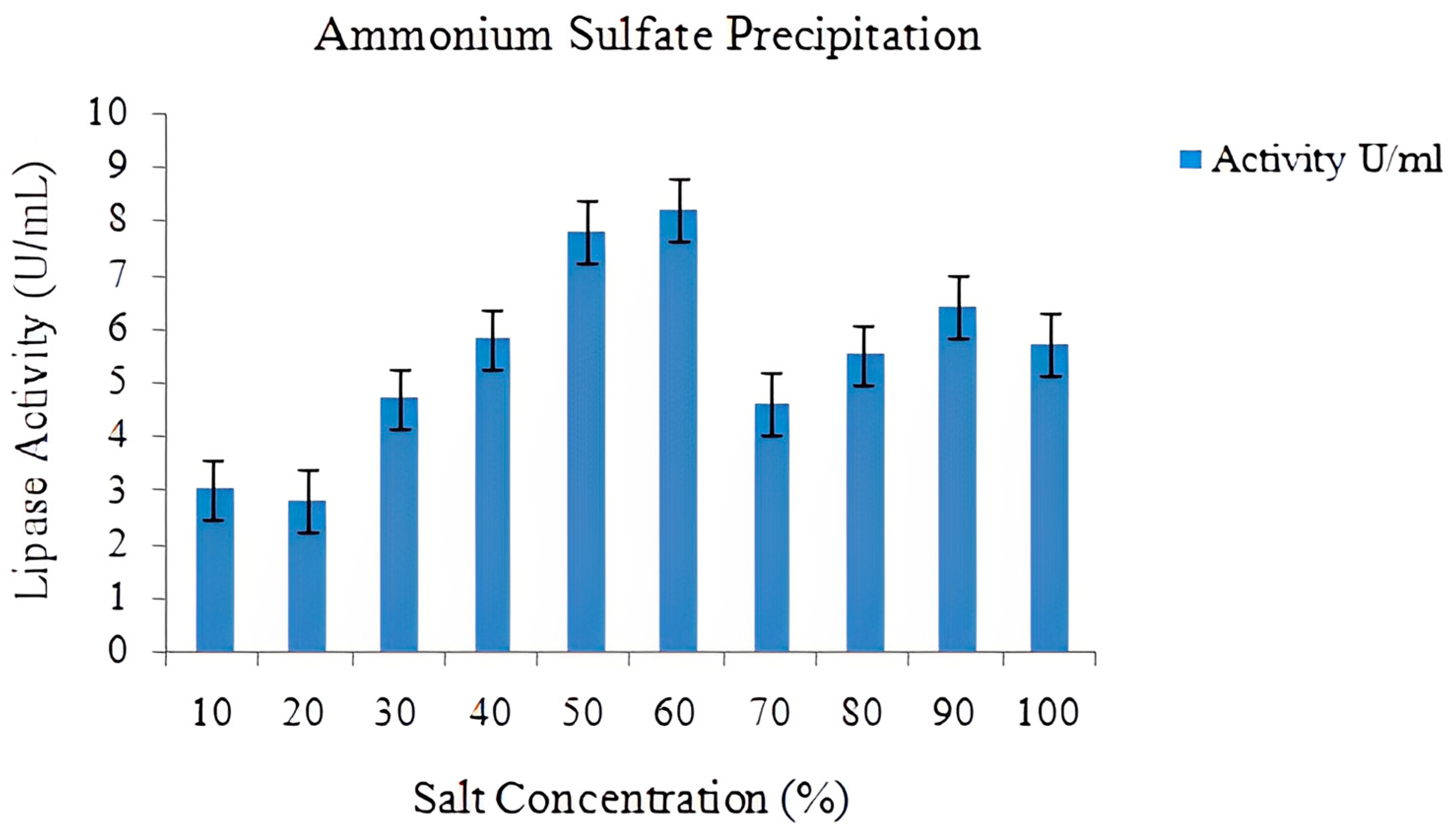
2.6. Lipase Purification
2.7. Protein Estimation
2.8. Characterization of Fungal Lipase
2.8.1. Effect of Temperature on Lipase Activity
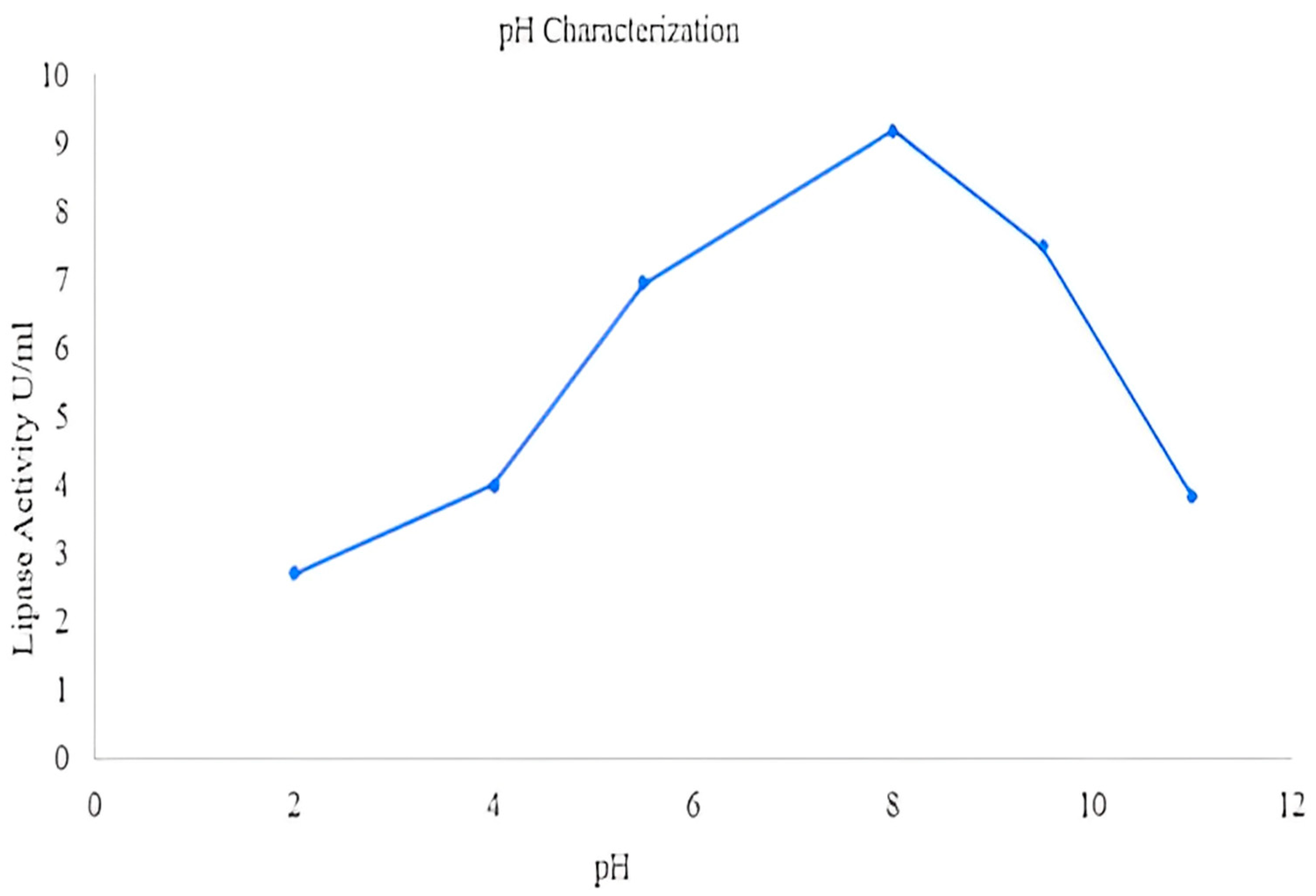
2.8.2. Effect of pH on Lipase Activity
3. Materials and Methods
3.1. Substrate Collection
3.2. Strain for Lipase Production
3.3. Screening of Fungi for Lipase Production
3.4. Vegetative Culturing
3.5. Screening of Substrate
3.6. Enzyme Extraction
3.7. Enzyme Assay
3.8. Standard Curve of Para-Nitrophenol
3.9. Optimization of Lipase by Response Surface Methodology
3.10. Preparation of Standard Curve of Bovine Serum Albumin (BSA)
3.11. Protein Content Determination
3.12. Protein Estimation
3.13. Bradford Reagent
3.14. Bradford Assay
3.15. Large Scale Enzyme Production
3.16. Purification of Lipase
3.16.1. Ammonium Sulfate Precipitation
3.16.2. Dialysis
3.16.3. Column Chromatography
3.17. Characterization of Fungal Lipase
3.17.1. Effect of Temperature on Lipase Activity
3.17.2. Effect of pH
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, S.; Yang, X. Technology prospectingon enzymes: Application, marketing and engineering. Comput. Struc. Biotec. J. 2012, 2, 2012–9017. [Google Scholar] [CrossRef]
- Hassan, S.; Sani, Y. Biogasoline: Anout of the boxsolution to the food for fuel and land use competitions. Energy Convers. Manag. 2015, 89, 349–367. [Google Scholar] [CrossRef]
- Klibanov, A.M. Improving enzymes by using the minorganic solvents. Nature 2001, 409, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kumar, M. Microbialenzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef]
- Karmakar, M.; Ray, R. Current trends in research and application of microbial cellulases. Res. J. Microbiol. 2011, 6, 41–53. [Google Scholar] [CrossRef]
- Rollinson, A.N.; Jones, J. Ureaasa hydrogen carrier: Urea as a hydrogen carrier: A perspective on its potential for safe, sustainable and long-term energy supply. Energy Environ. Sci. 2011, 4, 1216–1224. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J. Advances in enzyme immobilization. Biotechnol. Lett. 2009, 31, 1639–1650. [Google Scholar] [CrossRef]
- Villeneuve, P. Plant lipases and their applications in oils and fats modification. Eur. J. Lipid Sci. Tec. 2003, 105, 308–317. [Google Scholar] [CrossRef]
- Klomklao, S.; Benjakul, S. Seafood enzymes: Biochemical properties and their impacton quality. Food Biochem. Food Proc. 2012, 271–273. [Google Scholar] [CrossRef]
- Khan, F.I.; Lan, D. The lid domainin lipases: Structural and functional determinan to enzymatic properties. Front. Bioeng. Biotech. 2017, 5, 16. [Google Scholar] [CrossRef]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Proc. Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Bharathi, D.; Rajalakshmi, G. Microbial lipases: An overview of screening, production and purification. Biocatal. Agri. Biotech. 2019, 22, 101368. [Google Scholar] [CrossRef]
- Hasan, F. Industrial applications of microbial lipases. Enzym. Microb. Tech. 2006, 39, 235–251. [Google Scholar] [CrossRef]
- Sharma, S.; Kanwar, S.S. Organic solvent tolerant lipases and applications. Sci. World J. 2014. [CrossRef] [PubMed]
- Hubbe, M.A.; Lavoine, N.; Lucia, L.A.; Dou, C. Formulating bioplastic composites for biodegradability, recycling, and performance: Areview. BioResources 2021, 16, 1. [Google Scholar] [CrossRef]
- Anobom, C.D.; Pinheiro, A.S. From structuretoc atalysis: Recent developments in the biotechnological applications of lipases. BioMed Res. Int. 2014, 2014, 684506. [Google Scholar] [CrossRef]
- Antczak, M.S.; Kubiak, A. Enzymatic biodiesel synthesis– key factors affecting efficiency of the process. Renew. Energy 2009, 34, 1185–1194. [Google Scholar] [CrossRef]
- Farrell, R.L.; Hata, K. Solving pitch problems in pulp and paper processes by the use of enzymes or fungi. In Biotechnology in the Pulp &Paper Industry; Springer: Berlin, Heidelberg, 2006; pp. 197–212. [Google Scholar] [CrossRef]
- Stoytcheva, M.; Montero, G. Analytical methods for lipases activity determination: A review. Anal. Chem. 2012, 8, 400–407. [Google Scholar] [CrossRef]
- Vakhlu, J. Yeast lipases: Enzyme purification, biochemical properties and gene cloning. Electron. J. Biotech. 2006, 9. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G. Thermophilic fungi: Their physiology and enzymes. Microbio. Mol. Bio. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef]
- Wolski, E.; Menusi, E. Partial characterization of lipases produced by a newly isolated Penicillium sp. in solid state and submerged fermentation: A comparative study. LWT-Food Sci. Tech. 2009, 42, 1557–1560. [Google Scholar] [CrossRef]
- Manan, M.; Webb, C. Design as pectsofsolid state fermentation as applied to microbial bioprocessing. J. Appllied Biotech. Bioeng. 2017, 4, 91. [Google Scholar] [CrossRef]
- Thomas, L.; Larroche, C. Current developments in solid-state fermentation. Biochem. Eng. J. 2013, 81, 146–161. [Google Scholar] [CrossRef]
- Lizardi-Jiménez, M.A.; Hernández Martínez, R. Solid state fermentation (SSF): Diversity of applications to valorize waste and biomass. Biotech. 2017, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Sivaramakrishnan, S.; Gangadharan, D. Edible oil cakes. In Biotechnology for Agro-Industrial Residues Utilisation; Springer: Berlin/Heidelberg, Germany, 2009; pp. 253–271. [Google Scholar]
- Granato, D.; deAraújo Calado, V.M. The use and importance of design of experiments (DOE) in process modelling in food science and technology. Math. Stat. Methods Food Sci. Tech. 2014, 1, 1–18. [Google Scholar]
- Muthuvelayudham, R.; Viruthagiri, T. Application of central composite design based response surface methodology in parameter optimization and on cellulase production using agricultural waste. Intr. J. Chem. Biol. Eng. 2010, 3, 97–104. [Google Scholar]
- Castro Ríos, K.; Montoya Estrada, C.N. Physicochemical treatments for the reduction of aflatoxins and Aspergillus niger in corn grains (Zea mays). J. Sci. Food Agri. 2020, 101, 3707–3713. [Google Scholar] [CrossRef]
- Fleuri, L.F.; deOliveira, M.C.; deLara Campos Arcuri, M.; Capoville, B.L.; Pereira, M.S.; Delgado, C.H.O.; Novelli, P.K. Production of fungal lipases using wheat bran and soybean bran and incorporation of sugarcane bagasse as a co-substrate in solid-state fermentation. Food Sci. Biotechnol. 2014, 23, 1199–1205. [Google Scholar] [CrossRef]
- Kumar, D.S.; Ray, S. Fungal lipase production by solid state fermentation- an overview. J. Anal. Bioanal. Tech. 2014, 6, 1–10. [Google Scholar]
- Goren, A.Y.; Recepoğlu, Y.K.; Khataee, A. Language of response surface methodology as an experimental strategy for electrochemical wastewater treatment process optimization. In Artificial Intelligence and Data Science in Environmental Sensing; Academic Press: Cambridge, MA, USA, 2022; pp. 57–92. [Google Scholar]
- Bagewadi, Z.K.; Mulla, S.I. Response surface methodology based optimization of keratinase production from Trichoderma harzianum isolate HZN12 using chicken feather waste and its application in dehairing of hide. J. Envir. Chem. Engi. 2018, 6, 4828–4839. [Google Scholar] [CrossRef]
- Nema, A.; Patnala, S.H. Production and optimization of lipase using Aspergillus niger MTCC 872 by solid-state fermentation. Bull. Natl. Res. Cent. 2019, 43, 82. [Google Scholar] [CrossRef]
- Mehta, A.; Bodh, U. Isolation of a novel lipase producing fungal isolate Aspergillus fumigatus and production optimization of enzyme. Biocatal. Biotransformation 2018, 36, 450–457. [Google Scholar] [CrossRef]
- El-Batal, A.I.; Farrag, A.A. Effect of environmental and nutritional parameters on the extracellular lipase production by Aspergillus niger. Int. Lett. Nat. Sci. 2016, 60, 18–29. [Google Scholar] [CrossRef]
- Mazhar, H.; Abbas, N.; Ali, S.; Sohail, A.; Hussain, Z.; Ali, S.S. Optimized production of lipase from Bacillus subtilis PCSIRNL-39. Afr. J. Biotech. 2017, 16, 1106–1115. [Google Scholar]
- Srivastava, N.; Srivastava, M. Applications of fungal cellulases in biofuel production: Advances and limitations. Renew. Sustain. Energy Rev. 2018, 82, 2379–2386. [Google Scholar] [CrossRef]
- Hasan, S.; Anwar, Z.; Khalid, W.; Afzal, F.; Zafar, M.; Ali, U.; Aljobair, M.O. Laccase Production from Local Biomass Using Solid State Fermentation. Fermentation 2023, 9, 179. [Google Scholar] [CrossRef]
- Mhetras, N.C.; Bastawde, K.B.; Gokhale, D.V. Purification and characterization of acidic lipase from Aspergillus niger NCIM 1207. Bioresour. Technol. 2009, 100, 1486–1490. [Google Scholar] [CrossRef]
- Khuraishi, A.A.; Jayalakshmi, M. Isolation and purification of anticoagulant enzymes from plant latex. J. Appl. Nat. Sci. 2019, 11, 217–222. [Google Scholar] [CrossRef]
- Sethi, B.K.; Nanda, P.K.; Sahoo, S. Characterization of biotechnologically relevant extracellular lipase produced by Aspergillus terreus NCFT 4269. 10. Braz. J. Microb. 2016, 47, 143–149. [Google Scholar] [CrossRef]
- Okunwaye, T.; Obibuzor, J.; Okogbenin, E. Purifcation and biochemical properties of lipase from Raphia palm fruit mesocarp. Afr. J. Biochem. Res. 2015, 9, 73–80. [Google Scholar]
- Chetana, R.; Meenu, S. Optimization of physicochemical parameters and medium components for enhancing the production of resveratrol in endophytic fungus Aspergillus niger. Res. J. Biotech. 2019, 14, 10. [Google Scholar]
- deLima, E.E.; Franco, D.G. Biochemical characterization of apartially purified protease from Aspergillusterreus 7461 and its application as an environmentally friendly dehairingagent for leather industry. Prep. Biochem. Biotech. 2021, 51, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Bonjoch, N.P.; Tamayo, P.R. Protein content quantification by Bradford method. In Handbook of Plant Ecophysiology Techniques; Springer: Berlin/Heidelberg, Germany, 2001; pp. 283–295. [Google Scholar]
- Irshad, M.; Anwar, Z. Bio-processing of agro-industrial waste orange peel for induced production of pectinase by Trichoderma viridi; its purification and characterization. Turk. J. Biochem. 2014, 39, 9–18. [Google Scholar] [CrossRef]
- Chakraborty, K.; Raj, R.P. An extra-cellular alkaline metallolipase from Bacillus licheniformisMTCC 6824: Purification and biochemical characterization. Food chem. 2008, 109, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Bárcenas-moreno, G.E.M.A. Adaptation of soil microbial communities to temperature: Comparison of fungi and bacteria in a laboratory experiment. Glob. Change Biol. 2009, 15, 2950–2957. [Google Scholar] [CrossRef]
- Gupta, R.; Rathi, P.; Gupta, N.; Bradoo, S. Lipase assays for conventional and molecular screening: An overview. Biotech. Ap. Biochem. 2003, 37, 63–71. [Google Scholar] [CrossRef]
- Jones, C.G.; DanielHare, J.; Compton, S.J. Measuring plant protein with the Bradford assay: 1. Evaluation and standard method. J. Chem. Ecol. 1989, 15, 979–992. [Google Scholar] [CrossRef]
- Alnawajha, M.M.; Abdullah, S.R.S.; Hasan, H.A.; Othman, A.R.; Kurniawan, S.B. Effectiveness of using water-extracted Leucaena leucocephala seeds as a coagulant for turbid water treatment: Effects of dosage, pH, mixing speed, mixing time, and settling time. Biomass Conv. Bioref. 2022, 1–14. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford method for protein quantitation. In The Protein Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2009; pp. 17–24. [Google Scholar]
- Rashid, R.; Anwar, Z. Chitosan-alginate immobilized lipase based catalytic constructs: Development, characterization and potential applications. Int.J. Biol. Macromol. 2018, 119, 992–1001. [Google Scholar] [CrossRef]
- Prado, M.R.; Brito, É.H. Subculture on potato dextrose agar as a complement to the broth microdilution assay for Malassezia pachydermatis. J. Microbiol. Methods 2008, 75, 341–343. [Google Scholar] [CrossRef]
- Nadeem, F.; Mehmood, T.; Anwar, Z.; Saeed, S.; Bilal, M.; Meer, B. Optimization of bioprocess steps through response surface methodology for the production of immobilized lipase using Chaetomium globosum via solid-state fermentation. Biomass Convers. Biorefinery. 2011, 11, 1–12. [Google Scholar] [CrossRef]
- Jha, A.K.; Sit, N. Comparison of response surface methodology (RSM) and artificial neural network (ANN) modelling for supercritical fluid extraction of phytochemicals from Terminalia chebula pulp and optimization using RSM coupled with desirability function (DF) and genetic algorithm (GA) and ANN with GA. Ind. Crops Prod. 2021, 170, 113769. [Google Scholar]


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr # | Substrate Size (g) | pH | Temperature (°C) | Inoculum Size (mL) | Moisture Level (%) | Incubation Time (Days) | Activity U/mL |
|---|---|---|---|---|---|---|---|
| 1 | 4 | 2 | 45 | 5 | 60 | 5 | 4.202 |
| 2 | 4 | 2 | 37 | 5 | 60 | 3 | 4.828 |
| 3 | 2 | 2 | 40 | 2 | 100 | 4 | 8.212 |
| 4 | 2 | 2 | 45 | 3 | 20% | 6 | 5.691 |
| 5 | 6 | 5.5 | 45 | 3 | 100 | 3 | 7.975 |
| 6 | 8 | 5.5 | 40 | 2 | 40 | 3 | 7.061 |
| 7 | 4 | 5.5 | 40 | 4 | 60 | 4 | 8.922 |
| 8 | 6 | 5.5 | 37 | 2 | 100 | 8 | 8.347 |
| 9 | 8 | 8 | 40 | 2 | 20 | 8 | 11.76 |
| 10 | 6 | 8 | 40 | 3 | 40 | 3 | 12.52 |
| 11 | 4 | 8 | 45 | 2 | 60 | 6 | 8.872 |
| 12 | 2 | 8 | 45 | 1 | 100 | 8 | 7.501 |
| 13 | 4 | 9.5 | 30 | 8 | 60 | 1 | 7.823 |
| 14 | 4 | 9.5 | 30 | 3 | 125 | 4 | 9.413 |
| 15 | 4 | 9.5 | 40 | 1 | 60 | 6 | 7.738 |
| 16 | 7 | 9.5 | 50 | 4 | 60 | 8 | 7.146 |
| 17 | 4 | 11 | 30 | 5 | 50 | 1 | 4.625 |
| 18 | 4 | 11 | 30 | 5 | 60 | 3 | 6.080 |
| 19 | 1 | 11 | 52 | 3 | 60 | 4 | 5.640 |
| 20 | 4 | 11 | 45 | 3 | 60 | 8 | 7.603 |
| Analysis of Variance | |||||
|---|---|---|---|---|---|
| Source | DF | Adj SS | Adj MS | F-Value | p-Value |
| Model | 19 | 87.801 | 4.62111 | 0.000 | 0.000 |
| Linear | 5 | 4.6304 | 0.92608 | 0.000 | 0.000 |
| Ph | 1 | 0.9406 | 0.94063 | 0.000 | 0.000 |
| Temperature | 1 | 1.3550 | 1.35496 | 0.000 | 0.000 |
| Inoculum Size | 1 | 1.3483 | 1.34826 | 0.000 | 0.000 |
| Moisture Level | 1 | 1.1967 | 1.19666 | 0.000 | 0.000 |
| Incubation Days | 1 | 2.2672 | 2.26723 | 0.000 | 0.000 |
| Square | 5 | 26.0522 | 5.21044 | 0.000 | 0.000 |
| pH × Ph | 1 | 6.6295 | 6.82954 | 0.000 | 0.000 |
| Temperature × Temperature | 1 | 0.9211 | 0.92106 | 0.000 | 0.000 |
| Inoculum size × Inoculum size | 1 | 3.7382 | 3.73816 | 0.000 | 0.000 |
| Moisture level × Moisture level | 1 | 2.9145 | 2.91453 | 0.000 | 0.000 |
| Incubation Period × IncubationPeriod | 1 | 0.0527 | 0.05272 | 0.000 | 0.000 |
| 2-Way Interaction | 9 | 24.4761 | 2.71956 | 0.000 | 0.000 |
| pH × Temperature | 1 | 1.6593 | 1.65935 | 0.000 | 0.000 |
| pH × Inoculum size | 7 | 0.0138 | 0.01380 | 0.000 | 0.000 |
| pH × Moisture Level | 1 | 4.1829 | 4.18292 | 0.000 | 0.000 |
| pH × Incubation Period | 1 | 2.9296 | 2.92965 | 0.000 | 0.000 |
| Temperature × Inoculum Size | 1 | 0.5634 | 0.56339 | 0.000 | 0.000 |
| Temperature × Moisture Level | 1 | 0.5634 | 1.43103 | 0.000 | 0.000 |
| Temperature × Incubation Period | 1 | 0.4733 | 0.47331 | 0.000 | 0.000 |
| Inoculum Size × Moisture Level | 1 | 0.8363 | 0.83635 | 0.000 | 0.000 |
| Inoculum Size × Incubation Period | 1 | 3.5240 | 3.52397 | 0.000 | 0.000 |
| Error | 0 | ||||
| Total | 19 | 87.8011 | |||
| Purification Step | Volume (mL) | Total Enzyme Activity (U/mL) | Total Protein Content (U/mg) | Specific Activity (mg) | Purification Fold |
|---|---|---|---|---|---|
| Crude Lipase | 250 | 3623 | 138.55 | 26.15 | 1 |
| (NH4)2SO4 | 100 | 1635 | 47.68 | 34.29 | 1.3 |
| Dialyzed | 25 | 411 | 8.562 | 48.03 | 1.83 |
| Pure lipase | 10 | 224 | 1.69 | 132.72 | 5.07 |
| Sr # | Substrate Size (g) | pH | Temperature (°C) | Inoculum Size (mL) | Moisture Level (%) | Incubation Time (Days) |
|---|---|---|---|---|---|---|
| 1 | 4 | 2 | 45 | 5 | 60% | 5 |
| 2 | 4 | 2 | 37 | 5 | 60% | 3 |
| 3 | 2 | 2 | 40 | 2 | 100% | 4 |
| 4 | 2 | 2 | 45 | 3 | 20% | 6 |
| 5 | 6 | 5.5 | 45 | 3 | 100% | 3 |
| 6 | 8 | 5.5 | 40 | 2 | 40% | 3 |
| 7 | 4 | 5.5 | 40 | 4 | 60% | 4 |
| 8 | 6 | 5.5 | 37 | 2 | 100% | 8 |
| 9 | 8 | 8 | 40 | 2 | 20% | 8 |
| 10 | 6 | 8 | 40 | 3 | 40% | 3 |
| 11 | 4 | 8 | 45 | 2 | 60% | 6 |
| 12 | 2 | 8 | 45 | 1 | 100% | 8 |
| 13 | 4 | 9.5 | 30 | 8 | 60% | 1 |
| 14 | 4 | 9.5 | 30 | 3 | 125% | 4 |
| 15 | 4 | 9.5 | 40 | 1 | 60% | 6 |
| 16 | 7 | 9.5 | 50 | 4 | 60% | 8 |
| 17 | 4 | 11 | 30 | 5 | 50% | 1 |
| 18 | 4 | 11 | 30 | 5 | 60% | 3 |
| 19 | 1 | 11 | 52 | 3 | 60% | 4 |
| 20 | 4 | 11 | 45 | 3 | 60% | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, U.; Anwar, Z.; Hasan, S.; Zafar, M.; Ain, N.u.; Afzal, F.; Khalid, W.; Rahim, M.A.; Mrabti, H.N.; AL-Farga, A.; et al. Bioprocessing and Screening of Indigenous Wastes for Hyper Production of Fungal Lipase. Catalysts 2023, 13, 853. https://doi.org/10.3390/catal13050853
Ali U, Anwar Z, Hasan S, Zafar M, Ain Nu, Afzal F, Khalid W, Rahim MA, Mrabti HN, AL-Farga A, et al. Bioprocessing and Screening of Indigenous Wastes for Hyper Production of Fungal Lipase. Catalysts. 2023; 13(5):853. https://doi.org/10.3390/catal13050853
Chicago/Turabian StyleAli, Usman, Zahid Anwar, Shoaib Hasan, Muddassar Zafar, Noor ul Ain, Fareed Afzal, Waseem Khalid, Muhammad Abdul Rahim, Hanae Naceiri Mrabti, Ammar AL-Farga, and et al. 2023. "Bioprocessing and Screening of Indigenous Wastes for Hyper Production of Fungal Lipase" Catalysts 13, no. 5: 853. https://doi.org/10.3390/catal13050853