
Diclofenac Biodegradation by Microorganisms and with Immobilised Systems—A Review

Abstract
:1. Introduction
2. Diclofenac—Characteristics and Distribution in the Environment
3. Toxicity of Diclofenac to Non-Target Organisms
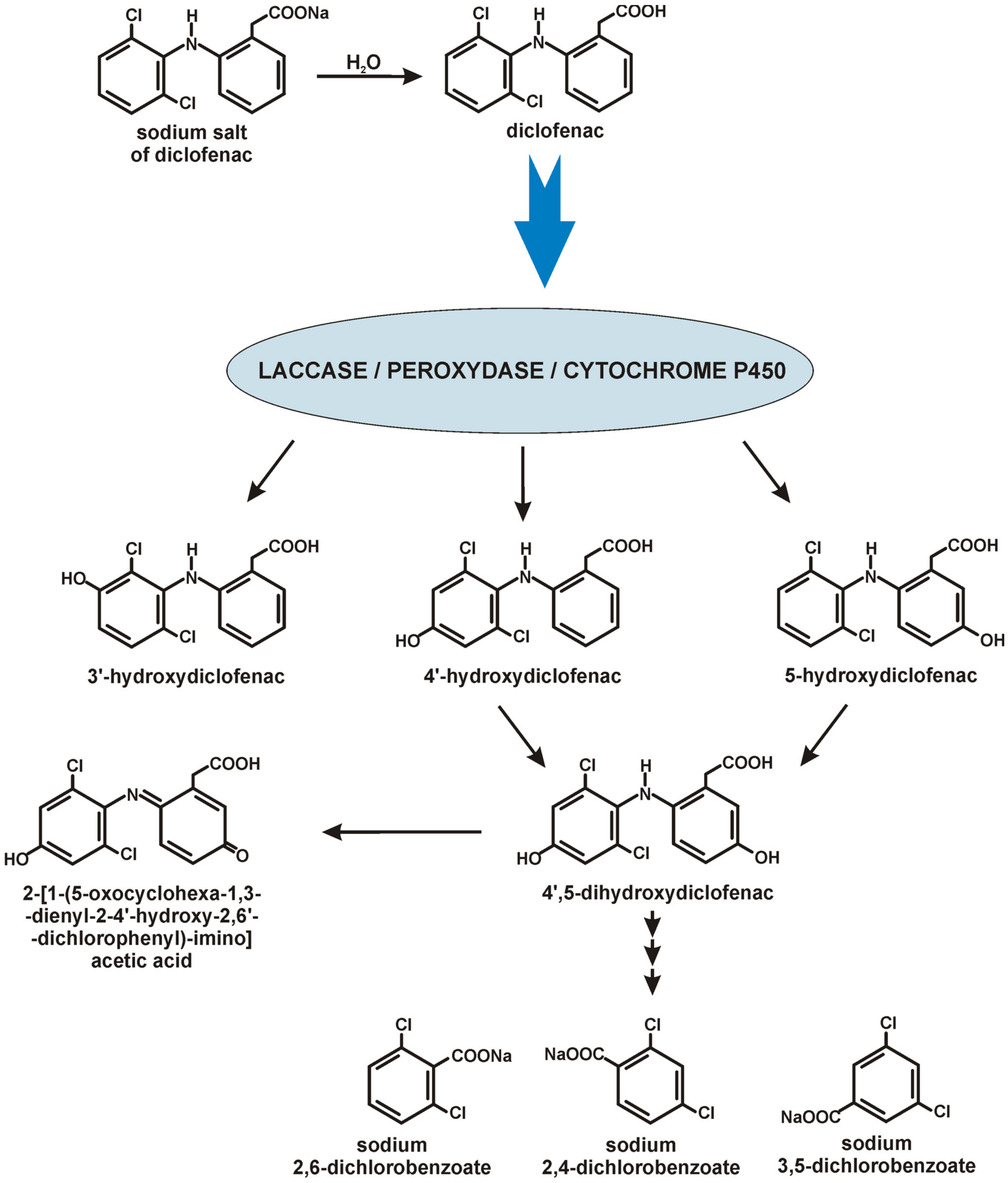
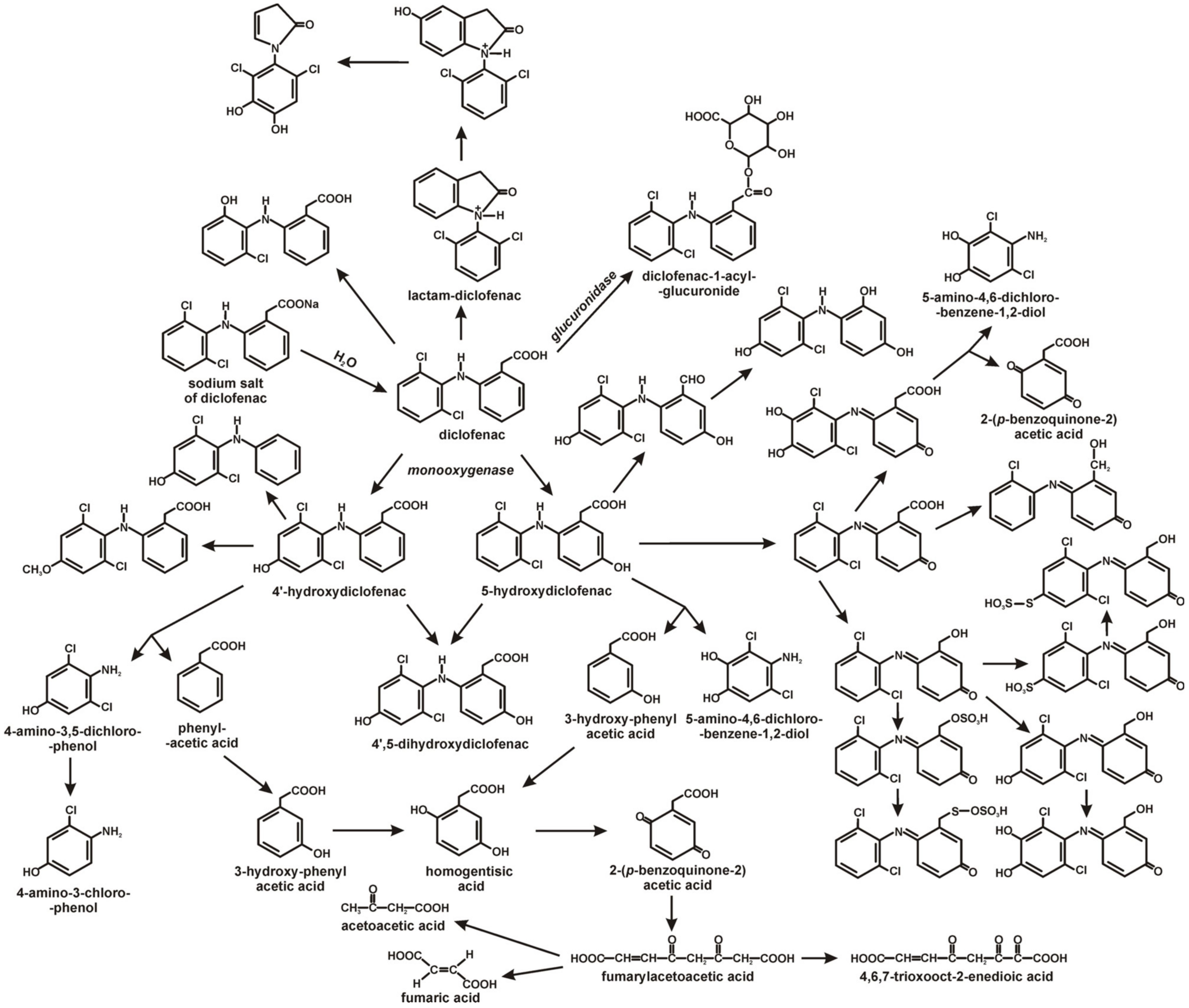
4. Diclofenac Biodegradation by Bacteria and Fungi
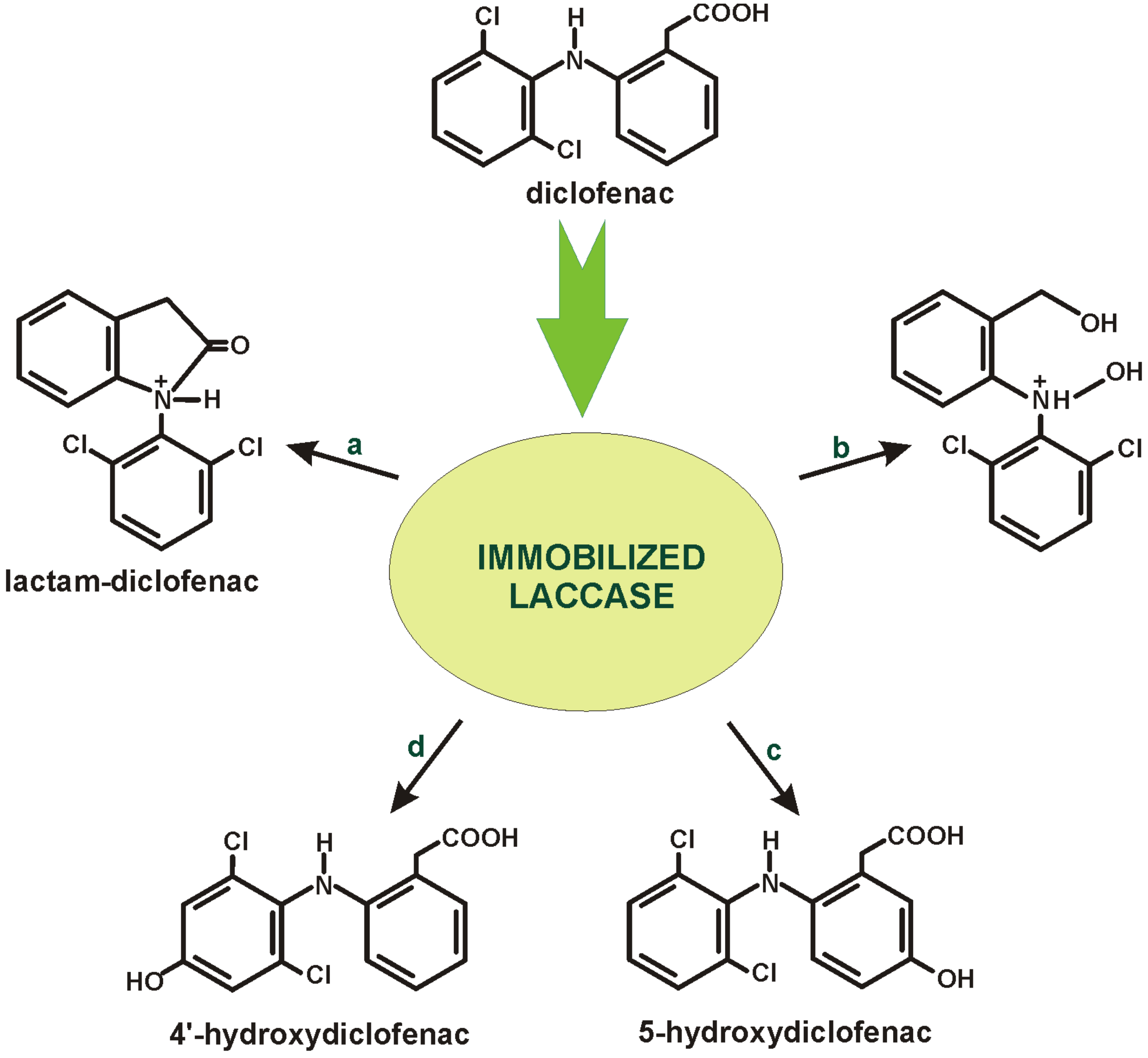
5. Diclofenac Biodegradation in Immobilised Systems
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Monteiro-Steagall, B.P.; Steagall, P.V.M.; Lascelles, B.D.X. Systematic review of non-steroidal anti-inflammatory drug-induced adverse effects in dogs. J. Vet. Intern. Med. 2013, 27, 1011–1019. [Google Scholar] [CrossRef]
- Brune, K.; Patrignani, P. New insights into the use of currently available non-steroidal anti-inflammatory drugs. J. Pain Res. 2015, 8, 105–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, U.; Wojcieszyńska, D. Biodegradation of non-steroidal anti-inflammatory drugs and their Influence on soil microorganisms. In Microbes and Enzymes in Soil Health and Bioremediation; Kumar, A., Sharma, S., Eds.; Springer: Singapore, 2019; pp. 379–401. Available online: https://link.springer.com/content/pdf/10.1007%2F978-981-13-9117-0_16.pdf (accessed on 10 February 2023).
- Du, Y.; Jia, X.; Zhong, L.; Jiao, Y.; Zhang, Z.; Wang, Z.; Feng, Y.; Bilal, M.; Cui, J.; Jia, S. Metal-organic frameworks with different dimensionalities: An ideal host platform for enzyme@MOF composites. Coord. Chem. Rev. 2022, 454, 214327. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, W.; Cai, Y. Laccase immobilization for water purification: A comprehensive review. Chem. Eng. J. 2021, 403, 126272. [Google Scholar] [CrossRef]
- Krasniqi, V.; Dimovski, A.; Domjanović, I.K.; Bilić, I.; Božina, N. How polymorphisms of the cytochrome P450 genes affect ibuprofen and diclofenac metabolism and toxicity. Arch. Ind. Hyg. Toxicol. 2016, 67, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Gao, H.; Zhang, Y.; Zhang, Y.; Zhou, X.; Li, C.; Gao, H. Developmental Toxicity of diclofenac and elucidation of gene regulation in zebrafish (Danio rerio). Sci. Rep. 2014, 4, 4841. [Google Scholar] [CrossRef] [Green Version]
- Navrozidou, E.; Remmas, N.; Melidis, P.; Karpouzas, D.G.; Tsiamis, G.; Ntougias, S. Biodegradation potential and diversity of diclofenac-degrading microbiota in an immobilised cell biofilter. Processes 2019, 7, 554. [Google Scholar] [CrossRef] [Green Version]
- Żur, J.; Piński, A.; Wojcieszyńska, D.; Smułek, W.; Guzik, U. Diclofenac degradation-enzymes, genetic background and cellular alterations triggered in diclofenac-metabolising strain Pseudomonas moorei KB4. Int. J. Mol. Sci. 2020, 21, 6786. [Google Scholar] [CrossRef]
- Moreira, I.S.; Bessa, V.S.; Murgolo, S.; Piccirillo, C.; Mascolo, G.; Castro, P.M.L. Biodegradation of diclofenac by the bacterial strain Labrys portucalensis F11. Ecotoxicol. Environ. Saf. 2018, 152, 104–113. [Google Scholar] [CrossRef]
- Biel-Maeso, M.; Corada-Fernandez, C.; Lara-Martin, P.A. Monitoring the occurrence of pharmaceuticals in soil irrigated with reclaimed wastewater. Environ. Pollut. 2018, 235, 312–321. [Google Scholar] [CrossRef]
- Ebele, A.J.; Abdallah, M.A.-E.; Harrad, S. Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerg. Contam. 2017, 3, 1–16. [Google Scholar] [CrossRef]
- Scheurell, M.; Franke, S.; Shah, R.M.; Hühnerfuss, H. Occurrence of diclofenac and its metabolites in surface water and effluent samples from Karachi, Pakistan. Chemosphere 2009, 77, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.M.; Sydnes, L.K.; Alarif, W.M.; Al-Lihaibi, S.S.; Aly, M.M.; Aanrud, S.G.; Kallenborn, R. Diclofenac and two of its photooxidation products in the marine environment: Their toxicology and occurence in Red Sea coastal waters. Environ. Chem. Ecotoxicol. 2019, 1, 19–25. [Google Scholar] [CrossRef]
- Čelić, M.; Gros, M.; Farre, M.; Barcelo, D.; Petrovic, M. Pharmaceuticals as chemical markers of wastewater contamination in the vulnerable area of the Ebro Delta (Spain). Sci. Total. Environ. 2019, 652, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Baranowska, I.; Kowalski, B. Using HPLC method with DAD detection for the simultaneous determination of 15 drugs in srface water and wastewater. Polish J. Environ. Stud. 2011, 20, 21–28. [Google Scholar]
- Reis-Santos, P.; Paisa, M.; Duarte, B.; Caçador, I.; Freitas, A.; Vila Pouca, A.S.; Barbosa, J.; Leston, S.; Rosa, J.; Ramos, F.; et al. Screening of human and veterinary pharmaceuticals in estuarine waters: A baseline assessment for the Tejo estuary. Mar. Pollut. Bull. 2018, 135, 1079–1084. [Google Scholar] [CrossRef]
- Lolić, A.; Paěga, P.; Santos, L.H.M.L.M.; Ramos, S.; Correia, M.; Delerue-Matos, C. Assessment of non-steroidal anti-inflammatory and analgesic pharmaceuticals in seawaters of North of Portugal: Occurrence and environmental risk. Sci. Total. Environ. 2015, 508, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Huebner, M.; Weber, E.; Niessner, R.; Boujday, S.; Knopp, D. Rapid analysis of diclofenac in freshwater and wastewater by a monoclonal antibody-based highly sensitive ELISA. Anal. Bioanal. Chem. 2015, 407, 8873–8882. [Google Scholar] [CrossRef]
- Yang, L.; He, J.T.; Su, S.H.; Cui, Y.F.; Huang, D.L.; Wang, G.C. Occurrence, distribution, and attenuation of pharmaceuticals and personal care products in the riverside groundwater of the Beiyun River of Beijing, China. Environ. Sci. Pollut. Res. 2017, 24, 15838–15851. [Google Scholar] [CrossRef]
- Cherik, D.; Benali, M.; Louhab, K. Occurrence, ecotoxicology, removal of diclofenac by adsorption on activated carbon and biodegradation and its effect on bacterial community: A review. World Sci. News 2015, 10, 116–144. [Google Scholar]
- Hanif, H.; Waseem, A.; Kali, S.; Qureshi, N.A.; Majid, M.; Iqbal, M.; Ur-Rehman, T.; Tahir, M.; Yousaf, S.; Iqbal, M.M.; et al. Environmental risk assessment of diclofenac residues in surface waters and wastewaters: A hidden global threat to aquatic ecosystem. Environ. Monit. Assess. 2020, 192, 204. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, G.; Sampath, S.; Selvaraj, K.K.; Larsson, D.G.J.; Ramaswamy, B.R. Non-steroidal ant-inflammatory drugs in Indian rivers. Environ. Sci. Pollut. Res. 2014, 21, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Madikizela, L.M.; Chimuka, L. Occurrence of naproxen, ibuprofen, and diclofenac residues in wastewater and river water of KwaZulu-Natal Province in South Africa. Environ. Monit. Assess. 2017, 189, 348. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Cifoni, M.; Baratti, M.; Pieraccini, G.; Di Marzio, W.D.; Galassi, D.M.P. Four scenarios of environmental risk of diclofenac in European groundwater ecosystems. Environ. Pollut. 2021, 287, 117315. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Alonso, S.; Merino, L.M.; Esteban, S.; de Alda, M.L.; Barcelo, D.; Duran, J.J.; Lopez-Martinez, J.; Acena, J.; Perez, S.; Mastoianni, N.; et al. Occurrence of pharmaceutical, recreational and psychotropic drug residues in surface water on the northern Antarctic Penisula region. Environ. Pollut. 2017, 229, 241–254. [Google Scholar] [CrossRef]
- Lapworth, D.J.; Baran, N.; Stuart, M.E.; Ward, R.S. Emerging organic contaminants in grounwater: A review of sources, fate and occurrence. Environ. Pollut. 2012, 163, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Fedrizzi, D.; Kosfeld, V.; Schlechtriem, C.; Ganz, V.; Derrer, S.; Rentsch, D.; Hollender, J. Biotransformation changes bioaccumulation and toxicity of diclofenac in aquatic organisms. Environ. Sci. Technol. 2020, 54, 4400–4408. [Google Scholar] [CrossRef]
- Joachim, S.; Beaudouin, R.; Daniele, G.; Geffard, A.; Bado-Nilles, A.; Tebby, C.; Palluel, O.; Dedourge-Geffard, O.; Fieu, M.; Bonnard, M.; et al. Effects of diclofenac on sentinel species and aquatic communities in semi-natural conditions. Ecotoxicol. Environ. Saf. 2021, 211, 111812. [Google Scholar] [CrossRef]
- Swan, G.E.; Cuthbert, R.; Quevedo, M.; Green, R.E.; Pain, D.J.; Bartels, P.; Cunningham, A.A.; Duncan, N.; Meharg, A.A.; Oaks, J.L.; et al. Toxicity of diclofenac to Gyps vultures. Biol. Lett. 2006, 2, 279–282. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, V.; Wolter, K.; Cuthbert, R.; Duncan, N. Veterinary diclofenac threatens Africa’s endangered vulture species. Regul. Toxicol. Pharmacol. 2009, 53, 205–208. [Google Scholar] [CrossRef]
- Sathishkumar, P.; Meena, R.A.A.; Palanisami, T.; Ashokkumar, V.; Palvannan, T.F.L. Occurrence, interactive effects and ecological risk of diclofenac in environmental compartments and biota-a review. Sci. Total Environ. 2020, 698, 134057. [Google Scholar] [CrossRef] [PubMed]
- Feito, R.; Valcárcel, Y.; Catalá, M. Biomarker assessment of toxicity with miniaturised bioassays: Diclofenac as a case study. Ecotoxicology 2012, 21, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Parolini, M. Toxicity of the non-steroidal anti-inflammatory drugs (NSAIDs) acetylsalicylic acid, paracetamol, diclofenac, ibuprofen and naproxen towards freshwater invertebrates: A review. Sci. Total Environ. 2020, 740, 140043. [Google Scholar] [CrossRef] [PubMed]
- Näslund, J.; Asker, N.; Fick, J.; Larsson, D.G.J.; Norrgren, L. Naproxen affects multiple organs in fish but is still an environmentally better alternative to diclofenac. Aquat. Toxicol. 2020, 227, 105583. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Schmieg, H.; Scheurer, M.; Köhler, H.R.; Triebskorn, R. Impact of the NSAID diclofenac on survival, development, behaviour and health of embryonic and juvenile stages of brown trout, Salmo trutta f. fario. Sci. Total Environ. 2017, 607, 1026–1036. [Google Scholar] [CrossRef]
- Ajima, M.N.O.; Ogo, O.A.; Audu, B.S.; Ugwoegbu, K.C. Chronic diclofenac (DCF) exposure alters both enzymatic and haematological profile of African catfish, Clarias gariepinus. Drug Chem. Toxicol. 2015, 38, 383–390. [Google Scholar] [CrossRef]
- Cardoso-Vera, J.D.; Islas-Flores, H.; SanJuan-Reyes, N.; Montero-Castro, E.I.; Galar-Martínez, M.; García-Medina, S.; Elizalde-Velázquez, A.; Dublán-García, O.; Gómez-Oliván, L.B. Comparative study of diclofenac-induced embryotoxicity and teratogenesis in Xenopus laevis and Lithobates catesbeianus, using the frog embryo teratogenesis assay: Xenopus (FETAX). Sci. Total Environ. 2017, 574, 467–475. [Google Scholar] [CrossRef]
- Kummerová, M.; Zezulka, S.; Babula, P.; Tříska, J. Possible ecological risk of two pharmaceuticals diclofenac and paracetamol demonstrated on a model plant Lemna minor. J. Hazard. Mater. 2016, 302, 351–361. [Google Scholar] [CrossRef]
- Gonzalez-Rey, M.; Bebianno, M.J. Effects of non-steroidal anti-inflammatory drug (NSAID) diclofenac exposure in mussel Mytilus galloprovincialis. Aquat. Toxicol. 2014, 148, 221–230. [Google Scholar] [CrossRef]
- Parolini, M.; Binelli, A.; Provini, A. Assessment of the potential cyto–genotoxicity of the non-steroidal anti-inflammatory drug (NSAID) diclofenac on the zebra mussel (Dreissena polymorpha). Water Air Soil Pollut. 2011, 217, 589–601. [Google Scholar] [CrossRef]
- Saravanan, M.; Hur, J.-H.; Arul, N.; Ramesh, M. Toxicological effects of clofibric acid and diclofenac on plasma thyroid hormones of an Indian major carp, Cirrhinus mrigala during short and long-term exposures. Environ. Toxicol. Pharmacol. 2014, 38, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.; Crane, M.; Merrington, G.; Ryan, J. Environmental quality standards for diclofenac derived under the European water framework directive: 2. Avian secondary poisoning. Environ. Sci. Eur. 2022, 34, 28. [Google Scholar] [CrossRef]
- Diniz, M.S.; Salgado, R.; Pereira, V.J.; Carvalho, G.; Oehmen, A.; Reis, M.A.M.; Noronha, J.P. Ecotoxicity of ketoprofen, diclofenac, atenolol and their photolysis byproducts un zebrafish (Danio rerio). Sci. Total Environ. 2015, 505, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Żur, J.; Marchlewicz, A.; Piński, A.; Guzik, U.; Wojcieszyńska, D. Degradation of diclofenac by new bacterial strains and its influence on the physiological status of cells. J. Hazard. Mater. 2021, 403, 124000. [Google Scholar] [CrossRef] [PubMed]
- Aissaoui, S.; Sifou, M.; Ouled-Haddar, H.; Benguedouar, L.; Lahouel, M. Toxicity assessment of diclofenac and its biodegradation metabolites toward mice. Toxicol. Environ. Health Sci. 2017, 9, 284–290. [Google Scholar] [CrossRef]
- Memmert, U.; Peither, A.; Burri, R.; Weber, K.; Schmidt, T.; Sumpter, J.P.; Hartmann, A. Diclofenac: New data on chronic toxicity and bioconcentration in fish. Environ. Toxicol. Chem. 2013, 32, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Siemieniuk, A.; Ludynia, M.; Rudnicka, M. Response of two crop plants, Zea mays L. and Solanum lycopersicum L., to diclofenac and naproxen. Int. J. Mol. Sci. 2021, 22, 8856. [Google Scholar] [CrossRef]
- Copolovici, L.; Timis, D.; Taschina, M.; Copolovici, D.; Cioca, G.; Bungau, S. Diclofenac influence on photosynthethic parameters and volatile organic compounds emission from Phaseolus vulgaris L. plants. Rev. Chem. 2017, 68, 2076–2078. [Google Scholar]
- Aracagök, D.Y.; Göker, H.; Cihangir, N. Biodegradation of diclofenac with fungal strains. Arch. Environ. Prot. 2018, 44, 55–62. Available online: https://journals.pan.pl/dlibra/publication/118181/edition/102790 (accessed on 10 February 2023).
- Kasonga, T.K.; Coetzee, M.A.A.; Kamika, I.; Momba, M.N.B. Assessing the fungal simultaneous removal efficiency of carbamazepine, diclofenac and ibuprofen in aquatic environment. Front. Microbiol. 2021, 12, 755972. [Google Scholar] [CrossRef]
- Domaradzka, D.; Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Toxicity of diclofenac and its biotransformation by Raoultella sp. DD4. Pol. J. Environ. Stud. 2016, 25, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Grandclément, C.; Piram, A.; Petit, M.-E.; Seyssiecq, I.; Laffont-Schwob, I.; Vanot, G.; Tiliacos, N.; Roche, N.; Doumenq, P. Biological removal and fate assessment of diclofenac using Bacillus subtilis and Brevibacillus laterosporus strains and ecotoxicological effects of diclofenac and 4′-hydroxy-diclofenac. J. Chem. 2020, 2020, 9789420. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Tyumina, E.A.; Kuzmina, M.V.; Vikhareva, E.V. Features of diclofenac biodegradation by Rhodococcus ruber IEGM 346. Sci. Rep. 2019, 9, 9159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murshid, S.; Dhakshinamoorthy, G.P. Biodegradation of sodium diclofenac and mefenamic acid: Kinetic studies, identification of metabolites and analysis of enzyme activity. Int. Biodeterior. Biodegrad. 2019, 144, 104756. [Google Scholar] [CrossRef]
- Stylianou, K.; Hapeshi, E.; Vasquez, M.I.; Fatta-Kassinos, D.; Vyrides, I. Diclofenac biodegradation by newly isolated Klebsiella sp. KSC: Microbial intermediates and ecotoxicological assessment. J. Environ. Chem. Eng. 2018, 6, 3242–3248. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheedb, T.; Nabeelb, F.; Iqbalc, H.M.N.; Zhao, Y. Hazardous contaminants in the environment and their laccase-assisted degradation—A review. J. Environ. Manag. 2019, 234, 253–264. [Google Scholar] [CrossRef]
- Neelkant, K.S.; Shankar, K.; Jayalakshmi, S.K.; Sreeramulu, K. Purification, biochemical characterisation, and facile immobilisation of laccase from Sphingobacterium ksn-11 and its application in transformation of diclofenac. Appl. Biochem. Biotechnol. 2020, 192, 831–844. [Google Scholar] [CrossRef]
- Zdarta, J.; Jankowska, K.; Wyszowska, M.; Kijeńska-Gawrońska, E.; Zgoła-Grześkowiak, A.; Pinelo, M.; Meyer, A.S.; Moszyński, D.; Jesionowski, T. Robust biodegradation of naproxen and diclofenac by laccase immobilised using electrospun nanofibers with enhanced stability and reusability. Mater. Sci. Eng. C 2019, 103, 109789. [Google Scholar] [CrossRef]
- Lonappan, L.; Liu, Y.; Rouissi, T.; Pourcela, F.; Brar, S.K.; Verma, M.; Surampalli, R.Y. Covalent immobilisation of laccase on citric acid functionalised micro- biochars derived from different feedstock and removal of diclofenac. Chem. Eng. 2018, 351, 985–994. [Google Scholar] [CrossRef]
- Lonappan, L.; Liu, Y.; Rouissi, T.; Brar, S.K.; Verma, M.; Surampalli, R.Y. Adsorptive immobilisation of agro-industrially produced crude laccase on various micro-biochars and degradation of diclofenac. Sci. Total Environ. 2018, 640–641, 1251–1258. [Google Scholar] [CrossRef]
- Masjoudi, M.; Golgoli, M.; Nejad, Z.G.; Sadeghzadeh, S.; Borghei, S.M. Pharmaceuticals removal by immobilised laccase on polyvinylidene fluoride nanocomposite with multi-walled carbon nanotubes. Chemosphere 2021, 263, 128043. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.N.; Hai, F.I.; Dosseto, A.; Richardson, C.; Price, W.E.; Nghiem, L.D. Continuous adsorption and biotransformation of micropollutants by granular activated carbon-bound laccase in a packed-bed enzyme reactor. Biores. Technol. 2016, 210, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, X.; Wang, X.; Sun, Y.; Li, W.; Chen, L.; Zhao, J. Degradation of diclofenac using palladized anaerobic granular sludge: Effects of electron donor, reaction medium and deactivation factors. J. Hazard. Mater. 2018, 365, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.C.V.; Serbent, M.P.; Skoronski, E. Application of immobilised mycelium-based pellets for the removal of organochlorine compounds: A review. Water Sci. Technol. 2021, 83, 1781–1796. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, Y.; Lin, L.; Tian, X.; Ciu, H.; Zhao, F. Degradation of diclofenac by B. subtilis through a cytochrome P450-dependent pathway. Environ. Technol. Innov. 2020, 20, 101160. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, M.; Sanroman, M.A.; Moldes, D. Recent development and applications of immobilized laccase. Biotechnol. Adv. 2013, 31, 1808–1825. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sources | Concentration | References |
|---|---|---|
| Europe | ||
| Soil (Jerez de la Frontera, Spain) | Nd1 –5.06 ng/g | [11] |
| Sediments Ebro Delta region (Catalonia, Spain) | 6.8–7.5 ng/g | [15] |
| Wisła river (Skoczów, Poland) | 74 ng/L | [16] |
| Odra river (Wrocław, Poland) | 0.429 μg/L | [16] |
| Warta river (Częstochowa, Poland) | 0.277 μg/L | [16] |
| Danube river (Budapest, Hungary) | 7–90 ng/L | [12] |
| Aabach river (Switzerland) | 11–310 ng/L | [12] |
| Swiss lakes (Switzerland) | 1–12 ng/L | [12] |
| Vltava river (Prague, The Czech Republic) | 0.104 μg/L | [16] |
| Tejo estuary (Portugal) | 51.8 ng/L | [17] |
| Seawater (Portugal) | 30.6 ng/L | [18] |
| Isar River (Germany) | 9–13 ng/L | [19] |
| Wörthsee lake (Germany) | 10–15 ng/L | [19] |
| Asia | ||
| Beiyun River (China) | 1.8–1300 ng/L | [20] |
| Huangpu River (China) | 13.6 ng/L | [21] |
| Malir River (Karachi, Pakistan) | 0.08–0.3 μg/L | [13] |
| Korang River (Rawalpindi-Islamabad, Pakistan) | 28 μg/L | [22] |
| Sawan River (Rawalpindi-Islamabad, Pakistan) | 62 μg/L | [22] |
| Gumrah Kas (Rawalpindi-Islamabad, Pakistan) | 14 μg/L | [22] |
| Ling Stream (Rawalpindi-Islamabad, Pakistan) | 23 μ/l | [22] |
| Kaveri river (India) | 103 ng/L | [23] |
| Africa | ||
| Mbokodweni river (KwaZulu-Natal, South Africa) | 0.9–5.3 μg/L | [24] |
| Umgeni River (KwaZulu-Natal, South Africa) | 10 μg/L | [25] |
| Red Sea (Saudi Arabia) | 26.9 ng/L | [14] |
| Antarctica | ||
| Stream (Fildes Peninsula, Antarctica) | 84 ng/L | [26] |
| Stream (Seymour/Marambio Island, Antarctica) | 77 ng/L | [26] |
| North America | ||
| Groundwater survey (Montana, USA) | 46 ng/L | [27] |
| Mississippi river (Louisiana, USA) | 22–107 ng/L | [12] |
| South America | ||
| Natural waters (Rio de Janeiro, Brazil) | 0.01–0.06 mg/L | [12] |
| Organism | Exposition Time | Concentration mg/L | Effect | References |
|---|---|---|---|---|
| Danio rerio | 96 h | 0.48 ± 0.05 | Mortality-LC50 | [33] |
| 0.09 ± 0.02 | Teratogenicity-EC50 | |||
| 90 min | 0.00003 | Decreased level of lipid peroxidation in zebrafish embryo | ||
| 96 h | 0.001 | Reduced viability of gill cells | [34] | |
| 48 h | 0.01 | Reduced viability of digestive cells | ||
| 48 h | 0.001 | Reduced viability of haemocytes | ||
| Danio magna | 21 days | 2.0 | Mortality-LC50 | |
| 0.5 | Reduction in egg production | |||
| Gammarus pulex | 24 h | 216 | Mortality-LC50 | [28] |
| Hyalella azteca | 24 h | 175 | Mortality-LC50 | |
| Oncorhynchus mykiss | - | 0.001 | Cytological alterations in the liver, kidney, and gills | |
| Gasterosteus aculeatus | 28 days | 0.0046 | Renal hematopoietic hyperplasia, jaw lesions | [35] |
| 21 days | 0.271 | Mortality-LOEC | ||
| Salmo trutta f. fario | 25 days | 0.1 | Irregularly shaped and vesiculated hepatocytes with a lack of glycogen storage and degenerating nuclei | [36] |
| Dreissena polymorpha | 6 months | 0.00382 | High mortality rates, effects on immunity, and high genotoxicity | [29] |
| 1 h | 0.25 | Destabilisation of lysosomal membranes | [34] | |
| 0.06 | DNA fragmentation | |||
| Clarias gariepinus | 96 h | 25.12 | Mortality-LC50 | [37] |
| Lithobates catesbeianus | 96 h | 1 | Induction malformations such as axial malformations in the tail and notochord, oedema and stunted growth | [38] |
| Xenopus laevis | 96 h | 1 | Induction malformations such as axial malformations in the tail and notochord, oedema and stunted growth | [38] |
| Lemna minor | 10 days | 0.0001 | Decrease in the content of photosynthetic pigments, increased amount of reactive nitrogen and oxygen species in roots, increased lipid peroxidation, disturbation in membrane integrity | [39] |
| Mytillus galloprovincialis | 15 days | 0.25 | Induction of superoxide dismutase and glutathione reductase in the gills, high catalase activity and lipid peroxidation levels in the digestive gland | [40] |
| Oryzias latipes | 4 days | 0.001 | Induction of p53 gene expression | [41] |
| Cirrhinus mrigala | 35 days | 0.001 | Decrease of thyroxine and triiodothyronine levels | [42] |
| Gyps bengalensis | 36–58 h | 0.25/kg | Death from renal failure and visceral gout | [43] |
| Immobilisation Matrix/Technology | Pros and Cons of Matrix/Technology | Microorganism/Enzyme | References |
|---|---|---|---|
| Sodium alginate-silicon dioxide-polyvinyl alcohol | Highly effective in subsequent cycles with an electron mediator | Laccase (Sphingobacterium ksn-11) | [58] |
| Electrospun nanofibers poly(L-lactic acid)-co-poly(ε-caprolactone) | Thin structure, porosity, biocompatibility, a high number of functional groups | Laccase (Trametes versicolor) | [59] |
| Porcine manure biocarbon | High adsorption capacity, effectiveness, high storage stability | Laccase | [60,61] |
| Polyvinylidene chloride membrane modified with multi-wall carbon nanotubes | Resistance to contaminants, specific surface area, mechanical strength, water permeability, selectivity, thermal resistance | Laccase (Trametes hirsuta) | [62] |
| Granulated activated carbon | Large specific surface, high adsorption capacity, porous structure, availability | Laccase | [63] |
| Palladium nanoparticles | Resistance to aggregation | Microorganisms | [64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojcieszyńska, D.; Łagoda, K.; Guzik, U. Diclofenac Biodegradation by Microorganisms and with Immobilised Systems—A Review. Catalysts 2023, 13, 412. https://doi.org/10.3390/catal13020412
Wojcieszyńska D, Łagoda K, Guzik U. Diclofenac Biodegradation by Microorganisms and with Immobilised Systems—A Review. Catalysts. 2023; 13(2):412. https://doi.org/10.3390/catal13020412
Chicago/Turabian StyleWojcieszyńska, Danuta, Karolina Łagoda, and Urszula Guzik. 2023. "Diclofenac Biodegradation by Microorganisms and with Immobilised Systems—A Review" Catalysts 13, no. 2: 412. https://doi.org/10.3390/catal13020412