

A Novel Chalcone Derivative Regulates the Expression and Phosphorylation of ERK1/2 by Inhibiting Fli-1 Promoter Activity for Preventing the Malignant Progression of Erythroleukemia
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of ZH-254 on the Growth of Leukemia Cells
ZH-254 Inhibited Proliferation and Induced Apoptosis in Erythroleukemia Cells
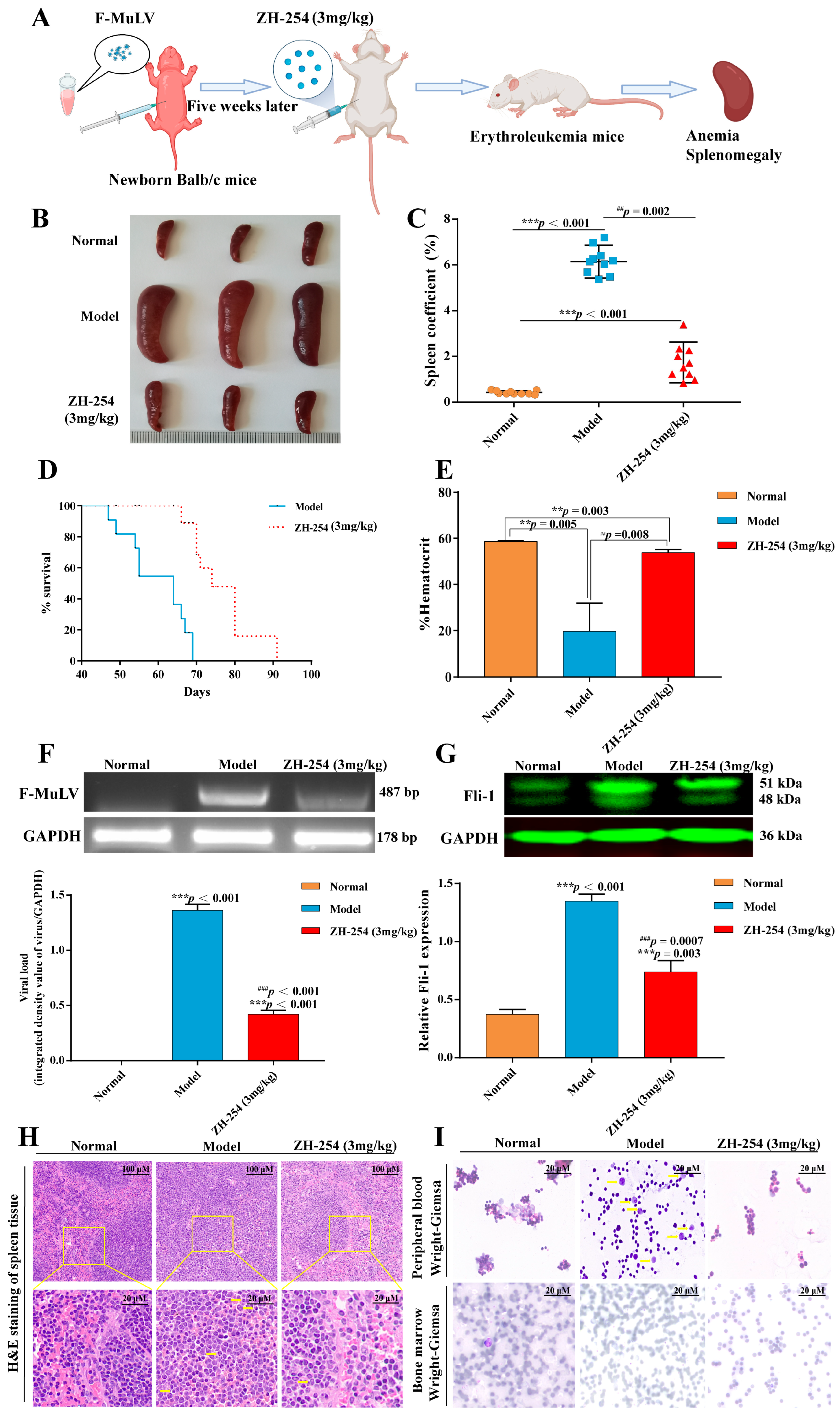
2.2. ZH-254 Could Effectively Prevent the Malignant Development of F-MuLV-Induced Erythroleukemia in Mouse
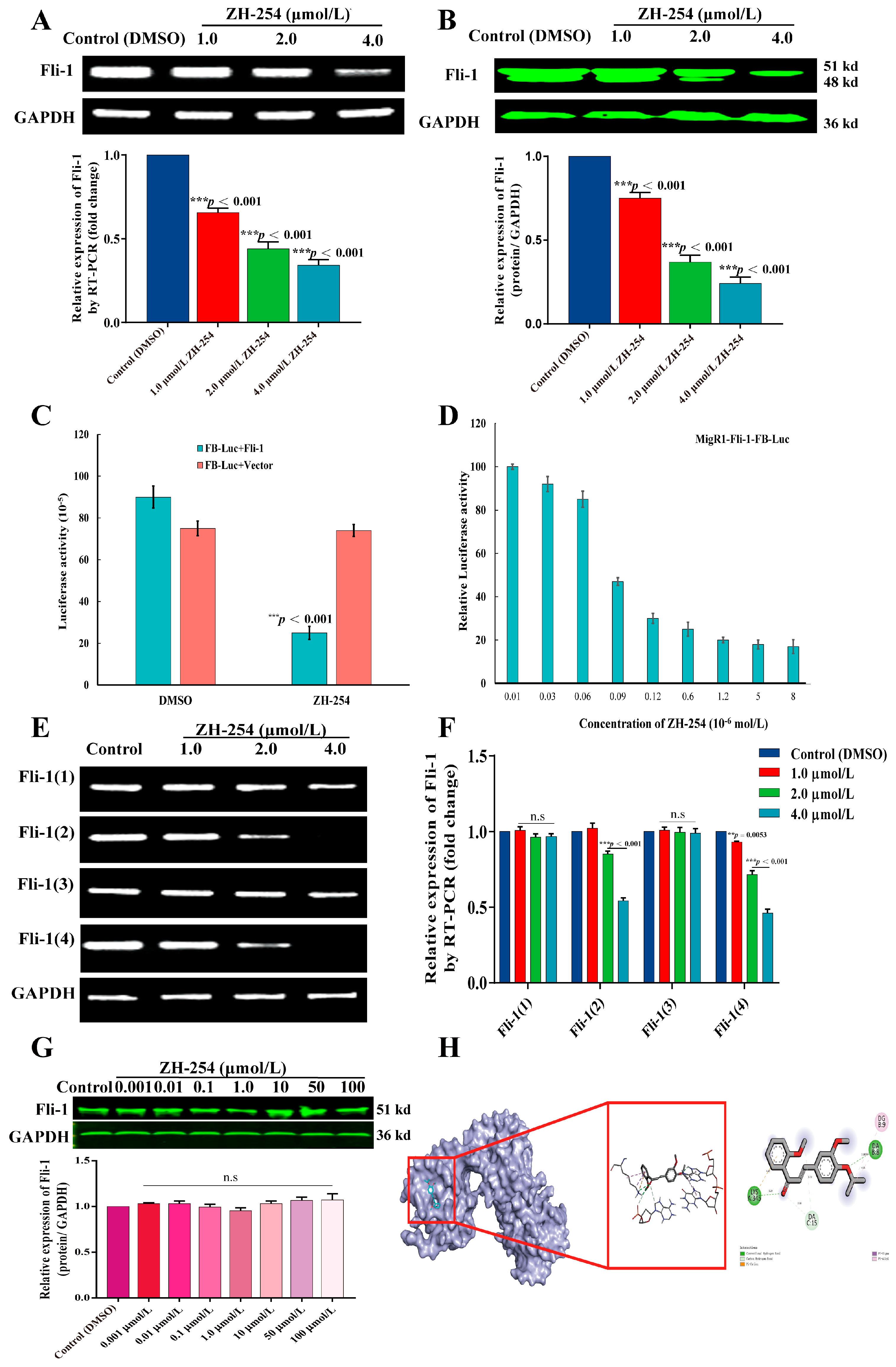
2.3. ZH-254 Downregulated Fli-1 Expression in HEL Cells by Inhibiting Promoter Activity
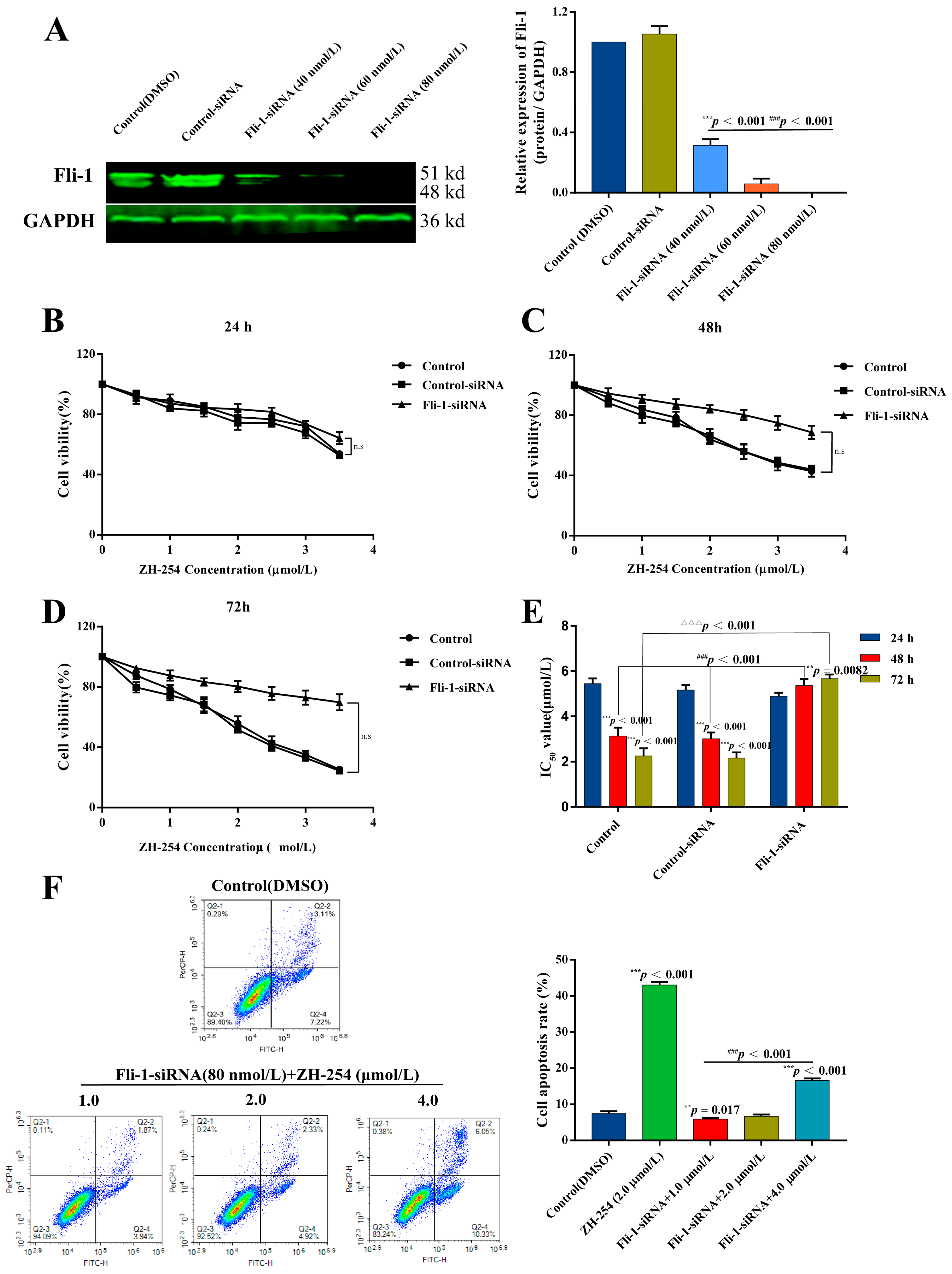
2.4. Validation of ZH-254 Targeting Fli-1 against HEL Cells
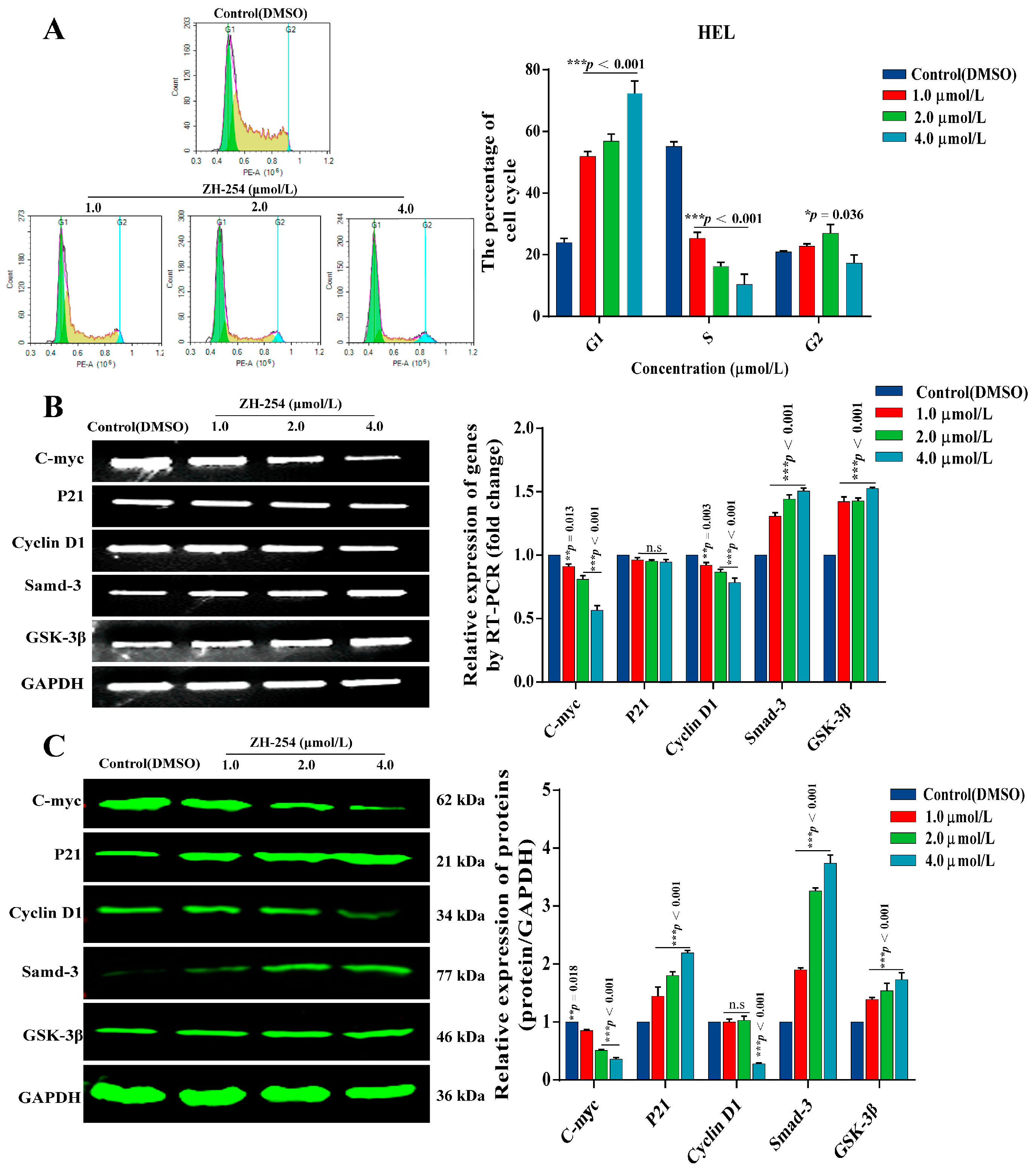
2.5. Effects of ZH-254 on the HEL Cell Cycle and Cycle-Related Genes
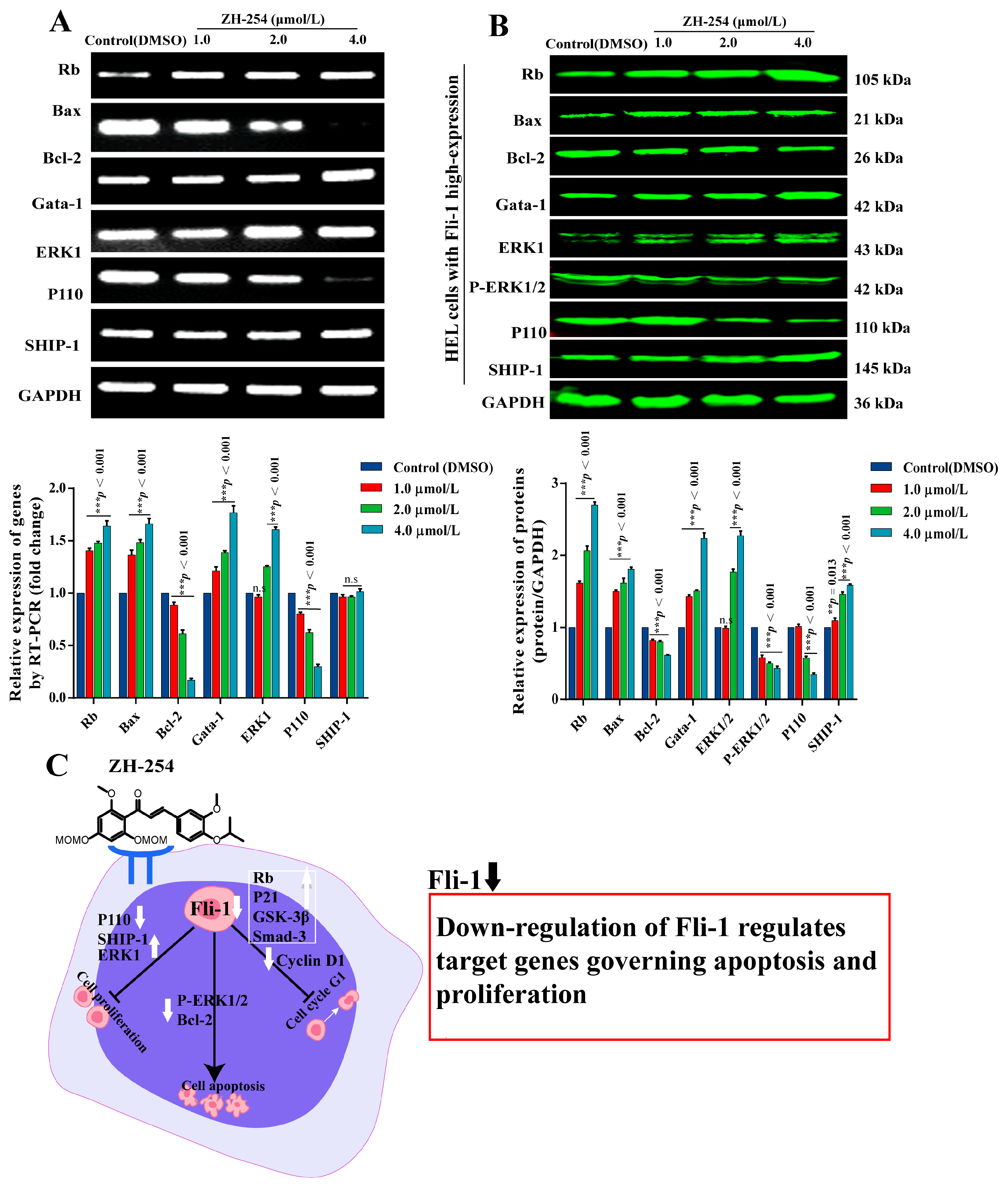
2.6. Regulation of Fli-1 Target Gene Expression in Erythroleukemia Cells by ZH-254
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents, and Instruments
4.2. Antibodies
4.3. Cell Culture and Animal Feeding
4.4. Compound Treatment
4.5. Cell Growth Assay
4.5.1. Cell Proliferation Assay
4.5.2. Cell Apoptosis Assay
4.5.3. Cell Cycle Assay
4.5.4. Hoechst 33258 Staining
4.5.5. TUNEL Staining Assay
4.5.6. Mitochondrial Membrane Potential (MMP) Assay
4.6. Luciferase Reporter Assay
4.7. ZH-254 In Vivo Activity Assay
4.7.1. In Vivo Assay
4.7.2. Hematoxylin and Eosin (H&E) Staining
4.7.3. Wright–Giemsa Staining
4.8. Validation Assay
4.8.1. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.8.2. Western Blotting Assay
4.8.3. Molecular Docking
4.8.4. Cellular Thermal Shift Assay (CETSA)
4.8.5. Gene Silencing Assay
4.9. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Animal Care
References
- Fagnan, A.; Bagger, F.O.; Piqué-Borràs, M.R.; Ignacimouttou, C.; Caulier, A.; Lopez, C.K.; Robert, E.; Uzan, B.; Gelsi-Boyer, V.; Aid, Z.; et al. Human erythroleukemia genetics and transcriptomes identify master transcription factors as functional disease drivers. Blood 2020, 136, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Cao, S.; Xu, R. Cancer incidence, mortality, and burden in China: A time-trend analysis and comparison with the United States and United Kingdom based on the global epidemiological data released in 2020. Cancer Commun. 2021, 41, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, G.; Koutros, S.; Hofmann, J.N.; Sandler, D.P.; Lubin, J.H.; Lynch, C.F.; Lerro, C.C.; De Roos, A.J.; Parks, C.G.; Alavanja, M.C.; et al. Glyphosate Use and Cancer Incidence in the Agricultural Health Study. J. Natl. Cancer Inst. 2018, 110, 509–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, J.; Yoo, H. Residential radon exposure and leukemia: A meta-analysis and dose-response meta-analyses for ecological, case-control, and cohort studies. Environ. Res. 2021, 202, 111714. [Google Scholar] [CrossRef]
- Ko, P.S.; Liu, Y.C.; Yeh, C.M.; Gau, J.P.; Yu, Y.B.; Hsiao, L.T.; Tzeng, C.H.; Chen, P.M.; Chiou, T.J.; Liu, C.J.; et al. The uniqueness of morphological features of pure erythroid leukemia in myeloid neoplasm with erythroid predominance: A reassessment using criteria revised in the 2016 World Health Organization classification. PLoS ONE 2017, 12, e0172029. [Google Scholar] [CrossRef]
- Ben-David, Y.; Giddens, E.B.; Bernstein, A. Identification and mapping of a common proviral integration site Fli-1 in erythroleukemia cells induced by Friend murine leukemia virus. Proc. Natl. Acad. Sci. USA 1990, 87, 1332–1336. [Google Scholar] [CrossRef] [Green Version]
- Lakhanpal, G.K.; Vecchiarelli-Federico, L.M.; Li, Y.J.; Cui, J.W.; Bailey, M.L.; Spaner, D.E.; Dumont, D.J.; Barber, D.L.; Ben-David, Y. The inositol phosphatase SHIP-1 is negatively regulated by Fli-1 and its loss accelerates leukemogenesis. Blood 2010, 116, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Charfi, C.; Voisin, V.; Levros, L.C., Jr.; Edouard, E.; Rassart, E. Gene profiling of Graffi murine leukemia virus-induced lymphoid leukemias: Identification of leukemia markers and Fmn2 as a potential oncogene. Blood 2011, 117, 1899–1910. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, D.; Poliquin, L.; Kozak, C.A.; Rassart, E. Identification of a common viral integration region in Cas-Br-E murine leukemia virus-induced non-T-, non-B-cell lymphomas. J. Virol. 1991, 65, 7–15. [Google Scholar] [CrossRef]
- Rodenburg, M.; Fischer, M.; Engelmann, A.; Harbers, S.O.; Ziegler, M.; Lohler, J.; Stocking, C. Importance of receptor usage, Fli1 activation, and mouse strain for the stem cell specificity of 10A1 murine leukemia virus leukemogenicity. J. Virol. 2007, 81, 732–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly, R.A.; Bosselut, R.; Zucman, J.; Cormier, F.; Delattre, O.; Roussel, M.; Thomas, G.; Ghysdael, J. DNA-binding and transcriptional activation properties of the EWS-FLI-1 fusion protein resulting from the t(11;22) translocation in Ewing sarcoma. Mol. Cell Biol. 1994, 14, 3230–3241. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Thapa, P.; Upadhyay, S.P.; Suo, W.Z.; Singh, V.; Gurung, P.; Lee, E.S.; Sharma, R.; Sharma, M. Chalcone and its analogs: Therapeutic and diagnostic applications in Alzheimer’s disease. Bioorg. Chem. 2021, 108, 104681. [Google Scholar] [CrossRef]
- Wen, Z.; Zhang, Y.; Wang, X.; Zeng, X.; Hu, Z.; Liu, Y.; Xie, Y.; Liang, G.; Zhu, J.; Luo, H.; et al. Novel 3′,5′-diprenylated chalcones inhibited the proliferation of cancer cells in vitro by inducing cell apoptosis and arresting cell cycle phase. Eur. J. Med. Chem. 2017, 133, 227–239. [Google Scholar] [CrossRef]
- Cui, J.W.; Vecchiarelli-Federico, L.M.; Li, Y.J.; Wang, G.J.; Ben-David, Y. Continuous Fli-1 expression plays an essential role in the proliferation and survival of F-MuLV-induced erythroleukemia and human erythroleukemia. Leukemia 2009, 23, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Qiu, X.; Xu, R.Z.; Zhao, X.Y. Berbamine inhibits proliferation and induces apoptosis of KU812 cells by increasing Smad3 activity. J. Zhejiang Univ. Sci. B 2011, 12, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.J.; Cheng, A.C.; Lee, M.F.; Hsu, Y.C. Simvastatin induces G1 arrest by up-regulating GSK3beta and down-regulating CDK4/cyclin D1 and CDK2/cyclin E1 in human primary colorectal cancer cells. J. Cell Physiol. 2018, 233, 4618–4625. [Google Scholar] [CrossRef]
- Seth, A.; Watson, D.K. ETS transcription factors and their emerging roles in human cancer. Eur. J. Cancer 2005, 41, 2462–2478. [Google Scholar] [CrossRef]
- Mhawech-Fauceglia, P.; Herrmann, F.R.; Bshara, W.; Odunsi, K.; Terracciano, L.; Sauter, G.; Cheney, R.T.; Groth, J.; Penetrante, R.; Mhawech-Fauceglia, P. Friend leukaemia integration-1 expression in malignant and benign tumours: A multiple tumour tissue microarray analysis using polyclonal antibody. J. Clin. Pathol. 2007, 60, 694–700. [Google Scholar] [CrossRef]
- Yan, X.; Yu, Y.; Li, L.; Chen, N.; Song, W.; He, H.; Dong, J.; Liu, X.; Cui, J. Friend leukemia virus integration 1 is a predictor of poor prognosis of breast cancer and promotes metastasis and cancer stem cell properties of breast cancer cells. Cancer Med. 2018, 7, 3548–3560. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhai, Y.; La, J.; Lui, J.W.; Moore, S.P.G.; Little, E.C.; Xiao, S.; Haresi, A.J.; Brem, C.; Bhawan, J.; et al. Targeting Pan-ETS Factors Inhibits Melanoma Progression. Cancer Res. 2021, 81, 2071–2085. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, Y.; Hamlett, E.D.; Goodwin, A.J.; Halushka, P.V.; Carroll, S.L.; Liu, M.; Fan, H. Suppression of Fli-1 protects against pericyte loss and cognitive deficits in Alzheimer’s disease. Mol. Ther. J. Am. Soc. Gene Ther. 2022, 30, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, S.; Starck, J.; Gonnet, C.; Doubeikovski, A.; Melet, F.; Morle, F. Negative and translation termination-dependent positive control of FLI-1 protein synthesis by conserved overlapping 5′ upstream open reading frames in Fli-1 mRNA. Mol. Cell Biol. 2000, 20, 2959–2969. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, Y.; Gajendran, B.; Sample, K.M.; Zacksenhaus, E. Current insights into the role of Fli-1 in hematopoiesis and malignant transformation. Cell. Mol. Life Sci. CMLS 2022, 79, 163. [Google Scholar] [CrossRef]
- Liu, T.; Xia, L.; Yao, Y.; Yan, C.; Fan, Y.; Gajendran, B.; Yang, J.; Li, Y.J.; Chen, J.; Filmus, J.; et al. Identification of diterpenoid compounds that interfere with Fli-1 DNA binding to suppress leukemogenesis. Cell Death Dis. 2019, 10, 117. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Fan, Y.; Yuan, C.M.; Song, J.; Yao, Y.; Liu, W.; Gajendran, B.; Zacksenhaus, E.; Li, Y.; Liu, J.; et al. Selective ERK1/2 agonists isolated from Melia azedarach with potent anti-leukemic activity. BMC Cancer 2019, 19, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yu, J.; Cheng, S.; Peng, Z.; Luo, H. Transcription factor Fli-1 as a new target for antitumor drug development. Int. J. Biol. Macromol. 2022, 209, 1155–1168. [Google Scholar] [CrossRef]
- Howard, J.C.; Li, Q.; Chu, W.; Zochodne, B.; Kapoor, M.; Ung, Y.; Rosen, K.; Ben-David, Y. Bcl-2 expression in F-MuLV-induced erythroleukemias: A role for the anti-apoptotic action of Bcl-2 during tumor progression. Oncogene 2001, 20, 2291–2300. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, B.; Yu, J.; Huang, L.; Zeng, X.; Shen, X.; Ren, C.; Ben-David, Y.; Luo, H. Fli-1 Activation through Targeted Promoter Activity Regulation Using a Novel 3′, 5′-diprenylated Chalcone Inhibits Growth and Metastasis of Prostate Cancer Cells. Int. J. Mol. Sci. 2020, 21, 2216. [Google Scholar] [CrossRef]
- Lesault, I.; Quang, C.T.; Frampton, J.; Ghysdael, J. Direct regulation of BCL-2 by FLI-1 is involved in the survival of FLI-1-transformed erythroblasts. EMBO J. 2002, 21, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Yuan, C.; Yang, J.; Liu, T.; Yao, Y.; Xiao, X.; Gajendran, B.; Xu, D.; Li, Y.J.; Wang, C.; et al. Novel flavagline-like compounds with potent Fli-1 inhibitory activity suppress diverse types of leukemia. FEBS J. 2018, 285, 4631–4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Q.; Wen, Z.H.; Wan, K.; Yuan, D.; Zeng, X.; Liang, G.; Zhu, J.; Xu, B.; Luo, H. A novel synthesized 3′, 5′-diprenylated chalcone mediates the proliferation of human leukemia cells by regulating apoptosis and autophagy pathways. Biomed. Pharmacother. 2018, 106, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Wu, X.; Shi, Y.; Guo, W.; Luo, J.; Liu, H.; Zheng, L.; Du, Y.; Wang, P.; Wang, Q.; et al. 20-HETE synthesis inhibition attenuates traumatic brain injury-induced mitochondrial dysfunction and neuronal apoptosis via the SIRT1/PGC-1α pathway: A translational study. Cell Prolif. 2021, 54, e12964. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liang, Y.; Sun, G.; He, Q.; Qu, H.; Gao, P. UBQLN4 is activated by C/EBPbeta and exerts oncogenic effects on colorectal cancer via the Wnt/beta-catenin signaling pathway. Cell Death Discov. 2021, 7, 398. [Google Scholar] [CrossRef]
- Dignam, J.D.; Lebovitz, R.M.; Roeder, R.G. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983, 11, 1475–1489. [Google Scholar] [CrossRef] [Green Version]
- Restall, B.S.; Haven, N.J.M.; Kedarisetti, P.; Martell, M.T.; Cikaluk, B.D.; Silverman, S.; Peiris, L.; Deschenes, J.; Zemp, R.J. Virtual hematoxylin and eosin histopathology using simultaneous photoacoustic remote sensing and scattering microscopy. Opt. Express 2021, 29, 13864–13875. [Google Scholar] [CrossRef]
- Dziekan, J.M.; Yu, H.; Chen, D.; Dai, L.; Wirjanata, G.; Larsson, A.; Prabhu, N.; Sobota, R.M.; Bozdech, Z.; Nordlund, P. Identifying purine nucleoside phosphorylase as the target of quinine using cellular thermal shift assay. Sci. Transl. Med. 2019, 11, eaau3174. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | ||||
|---|---|---|---|---|
| HEL | K562 | Jurkat | HL-60 | |
| Inhibition rate (%) | 77.05 ± 0.28 | 34.57 ± 3.61 | 60.37 ± 0.31 | 53.29 ± 0.41 |
| The Proportion of Marrow Granulocyte (%) | Normal (n = 6) | Model (n = 6) | ZH-254 (3 mg/kg) (n = 6) |
|---|---|---|---|
| mature granulocyte | 23.83 ± 1.47 | 54.00 ± 2.45 ** | 66.17 ± 5.78 **,## |
| Promyelocyte | 15.31 ± 4.49 | 33.17 ± 2.32 ** | 20.18 ± 2.71 *,## |
| Myelocyte and metamyelocyte | 8.20 ± 1.60 | 16.83 ± 3.97 * | 13.17 ± 2.48 *,# |
| Genes | Primer | Primer Sequence (5′→3′) |
|---|---|---|
| Fli-1(total) | Forward | CAGCCCCACAAGATCAACCC |
| Reverse | CACCGGAGACTCCCTGGAT | |
| Fli-1(1) | Forward | ATGGACGGGACTATTAAGGA |
| Reverse | TCATGTCGGCCTTGGGGAGAT | |
| Fli-1(2) | Forward | ACTGCCTCGGGGAGTCCTGAC |
| Reverse | CTTGACGTTGACCCTCACTG | |
| Fli-1(3) | Forward | GTGGACTGCAGCGTTAGCAA |
| Reverse | GGCCATTCTTCTCGTCCATA | |
| Fli-1(4) | Forward | CAATCCTCACGATTGAGTGTC |
| Reverse | GGACTTTTGTTGAGGCCAGA | |
| CDK4 | Forward | ATGGCTACCTCTCGATATGAGC |
| Reverse | CATTGGGGACTCTCACACTCT | |
| C-myc | Forward | TCCTGGCAAAAGGTCAGAGT |
| Reverse | GTTGTGTGTTCGCCTCTTGA | |
| P21 | Forward | GCAGACCAGCATGACAGATTT |
| Reverse | GGATTAGGGCTTCCTCTTGGA | |
| Cyclin D1 | Forward | GCTGCGAAGTGGAAACCATC |
| Reverse | CCTCCTTCTGCACACATTTGAA | |
| Smad-3 | Forward | TGGACGCAGGTTCTCCAAAC |
| Reverse | CCGGCTCGCAGTAGGTAAC | |
| GSK-3β | Forward | ATGGCAGCAAGGTAACCACAG |
| Reverse | TCTCGGTTCTTAAATCGCTTGTC | |
| Rb | Forward | CTCTCGTCAGGCTTGAGTTTG |
| Reverse | GACATCTCATCTAGGTCAACTGC | |
| Bcl-2 | Forward | GGTGGGGTCATGTGTGTGG |
| Reverse | CGGTTCAGGTACTCAGTCATCC | |
| Gata-1 | Forward | CTGTCCCCAATAGTGCTTATGG |
| Reverse | GAATAGGCTGCTGAATTGAGGG | |
| ERK | Forward | TACACCAACCTCTCGTACATCG |
| Reverse | CATGTCTGAAGCGCAGTAAGATT | |
| P110 | Forward | TATTTGGACTTTGCGACAAGACT |
| Reverse | TCGAACGTACGGTCTGGATAG | |
| SHIP-1 | Forward | GCGTGCTGTATCGGAATTGC |
| Reverse | TGGTGAAGAACCTCATGGAGAC | |
| Fr.MuLV env | Forward Reverse | TGGAATCCTGTGGCATCCATGAAC TAAAACGCAGCTCAGTAACAGTCCG |
| GAPDH | Forward | GGAGCGAGATCCCTCCAAAAT |
| Reverse | GGCTGTTGTCATACTTCTCATGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, M.; Ma, Y.; Yu, J.; Ahmad, M.; Wan, X.; Yu, Z.; Xu, B.; Luo, H. A Novel Chalcone Derivative Regulates the Expression and Phosphorylation of ERK1/2 by Inhibiting Fli-1 Promoter Activity for Preventing the Malignant Progression of Erythroleukemia. Catalysts 2023, 13, 84. https://doi.org/10.3390/catal13010084
Mo M, Ma Y, Yu J, Ahmad M, Wan X, Yu Z, Xu B, Luo H. A Novel Chalcone Derivative Regulates the Expression and Phosphorylation of ERK1/2 by Inhibiting Fli-1 Promoter Activity for Preventing the Malignant Progression of Erythroleukemia. Catalysts. 2023; 13(1):84. https://doi.org/10.3390/catal13010084
Chicago/Turabian StyleMo, Min, Youfen Ma, Jia Yu, Mashaal Ahmad, Xinwei Wan, Zijiang Yu, Bixue Xu, and Heng Luo. 2023. "A Novel Chalcone Derivative Regulates the Expression and Phosphorylation of ERK1/2 by Inhibiting Fli-1 Promoter Activity for Preventing the Malignant Progression of Erythroleukemia" Catalysts 13, no. 1: 84. https://doi.org/10.3390/catal13010084