1. Introduction
The use of enzymes as catalysts in industrial applications is very attractive especially in terms of chemoselectivity, enantioselectivity, and energy demand [
1,
2], while the combination of bio- and chemocatalysis offers additional advantages [
3]. For the fast implementation of such catalytic processes, the cooperation of organic chemists with specialists from neighboring disciplines such as biocatalysis, industrial chemistry, and process analytical technologies are especially helpful.
The use of enzymes in an industrial context often requires their immobilization for easy recycling and simple downstream processing. A variety of materials ranging between natural and synthetic origins are available for immobilization and their nature and properties are shown to influence the interaction with the enzymes as well as their activity, specificity, and performance in the respective applications.
For this reason, a critical parameter that can help understanding the performance of an immobilized enzyme is the spatial distribution of the enzyme and substrate within an immobilization carrier. The enzyme distribution has been obtained experimentally by coupling microscopic techniques with staining [
4,
5], radioactive labeling [
5,
6,
7], or fluorescence labeling [
8,
9,
10]. Methods relying on staining or labelling have the inherent drawback of requiring additional steps during sample preparation and carry a risk of influencing the enzyme distribution, e.g., the solvents used in the labelling process penetrating into the carrier and causing a further diffusion of the enzyme. Infrared (IR) microscopic methods are label-free and are therefore not affected by these problems. IR spectroscopy is also quantitative, allowing conclusions about enzyme concentration to be drawn based on the absorbance of light.
In previous studies by Sinigoi et al. [
11] and Mei et al. [
12], synchrotron-coupled IR microscopes were used to investigate Synbeads and immobilized enzymes, respectively. While synchrotron light is well suited for this application, access to such specialized infrastructure is often highly restricted and requires special training and careful planning. In comparison, the benchtop infrared microscope employed for this work uses a conventional IR light source, making it far more accessible and easier to operate.
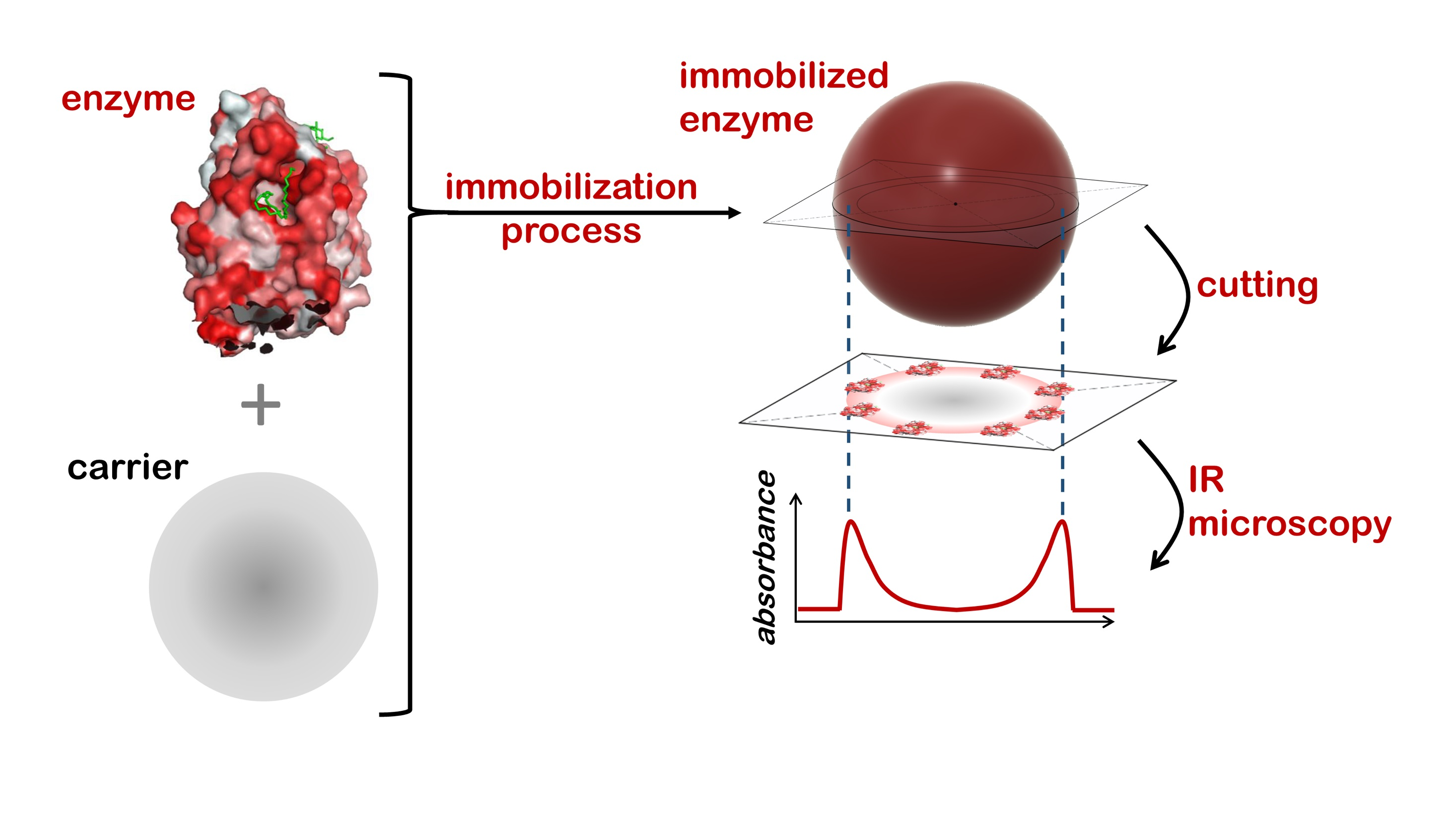
In this paper, sample preparation was streamlined, enabling the time-efficient processing of a batch of 5–10 samples through the use of novel, disposable, 3D-printed microscope slides. The immobilized enzyme beads were embedded in commercial histology-grade paraffin wax, cut into thin slices (sections) using a rotary microtome, and the sections mounted on the aforementioned slides. The paraffin wax used in this study offers considerable advantages over epoxy polymers [
11] or glycerin-based embedding media [
13] used in other studies, which require special slicing equipment and cryogenic temperatures, respectively. The 3D-printed slides are considerably (approx. a factor of 3000) cheaper than the barium fluoride disks conventionally used for sample mounting in transmission IR microscopy [
12].
Chen et al. (2008) demonstrated in a previous paper, that
Candida antarctica lipase B (CalB) was immobilized on the external surface of the resin Lewatit
® VP OC1600, with an enzyme layer thickness of 80–100 µm on beads with a particle size of about 650 µm [
14].
Nielsen et al. (2014) examined the activity and spatial distribution of CalB on polystyrene (PS) and polymethylmethacrylate (PMMA) carriers using fluorescence microscopy. They observed an enzyme layer thickness of 50–85 µm on PMMA beads and 10–20 µm on PS beads with particle sizes between 500–850 µm [
13].
In the present paper, we studied the effect of interaction of lipase CalB on two different resins by means of FT-IR microscopy. In one case the resin was a hydrophobic methacrylate-divinyl benzene resin (ECR1030M), and in the other case a hydrophilic methacrylic resin (ECR8204M).
The copolymer of ECR1030M contains aromatic rings that favor van der Waals interaction. ECR8204M resin contains only methacrylate functionalized with epoxide groups and thus favors covalent bonds with the -NH2 groups present on the enzyme as well as hydrogen bonds. The target of the study was to develop a streamline sample preparation used in combination with the FT-IR to verify the interaction of the enzyme with the polymeric structure in the case of predominantly hydrophobic interactions and in the case of predominantly hydrophilic and covalent interactions.
3. Discussion
The study showed that when CalB was immobilized on a hydrophobic resin as ECR1030M, we encountered a behavior similar to what was observed by Chen et al. (2008) for Novozyme 435 [
14], but a different behavior was observed for the hydrophilic resin ECR8204M. In this work, we also showed that on ECR1030M, the enzyme was immobilized only in the external shell of the polymer, in a section of about 50–70 µm. A similar behavior could be observed with Lewatit
® VP OC 1600, which has a similar chemical structure to ECR1030M [
15]. However, CalB fully penetrated the resin beads when immobilized on the hydrophilic resin ECR8204M. This contrasts with the observations from Chen et al., where the enzyme distribution on Amberzyme PMMA (similar hydrophilicity, same functional groups to ECR8204M) was not homogeneous, with the enzyme only present on the surface of the matrix [
14]. The pore size of Amberzyme (220 Å) was half of the pore size in ECR8204M (519 Å), which may explain the differences.
The main differences between the two enzyme distribution patterns found in this work is related to the hydrophobicity of the beads and the interaction that occurred between the enzyme and the resin. The interaction between CalB and ECR1030M is mainly hydrophobic and occurs rapidly in the first 15–60 min from the immobilization, and the loading front is narrower than in ECR8204M. As suggested by others [
14,
17], epoxy-activated resins would have a two-step immobilization mechanism where firstly the enzyme is adsorbed onto the bead surface and then the covalent bond is formed. In this case, the interaction takes more time, and this allows the enzyme to diffuse deeper inside the resin. Another hypothesis from Gross’s group [
12,
14,
18] points out that there could be repulsive interactions between the immobilized and free CalB forms, and this repulsion would be stronger with hydrophobic resins given the hydrophobic nature of CalB [
12] and would contribute to the binding profile differences between hydrophobic and hydrophilic resins.
The very interesting result is the measurement of the enzyme interaction with the resin during the immobilization process. We analyzed the enzyme diffusion in the beads over a period of 24 h, by analyzing after 0.25 h, 0.5 h, 1 h, 2 h, 4 h, 6 h, and 24 h of immobilization.
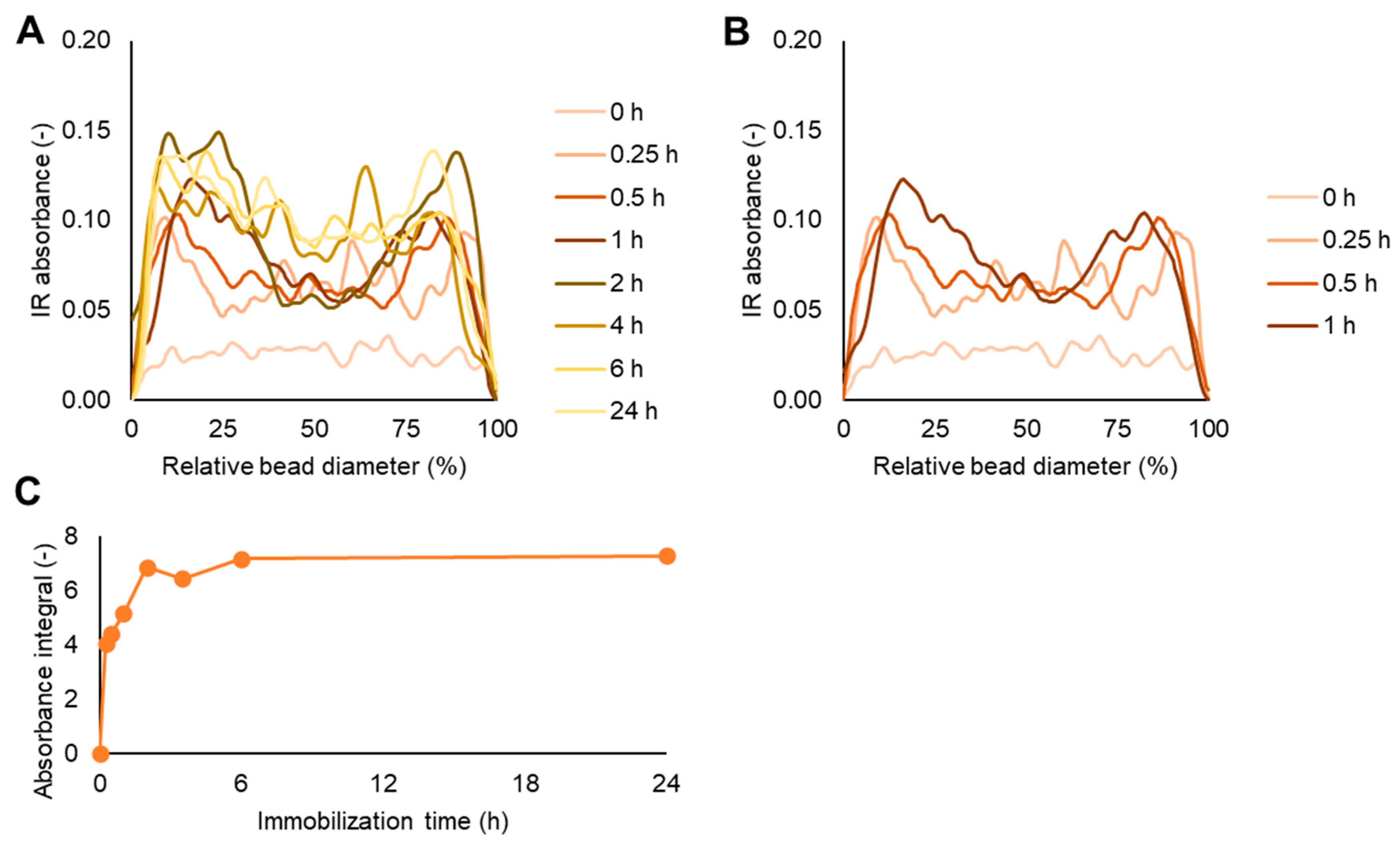
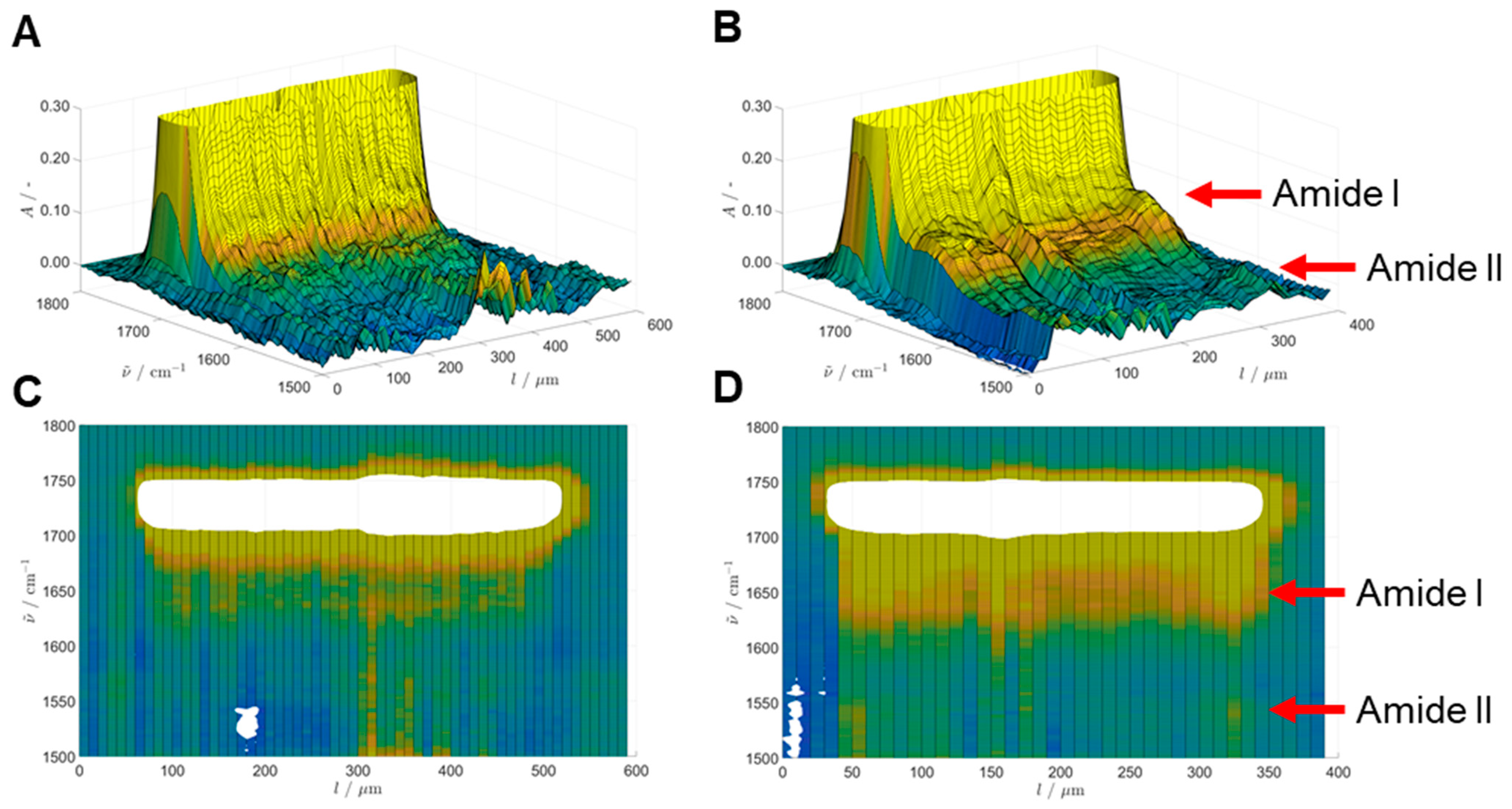
When CalB was immobilized on a hydrophilic resin such as ECR8204M (
Figure 3) the diffusion started on the external part of the bead and the majority of the enzyme was concentrated after 1 h immobilization in the 30 µm range. Over time, the enzyme diffused towards the center of the bead and 24 h were required to complete the immobilization and reach the center of the beads (
Figure 3).
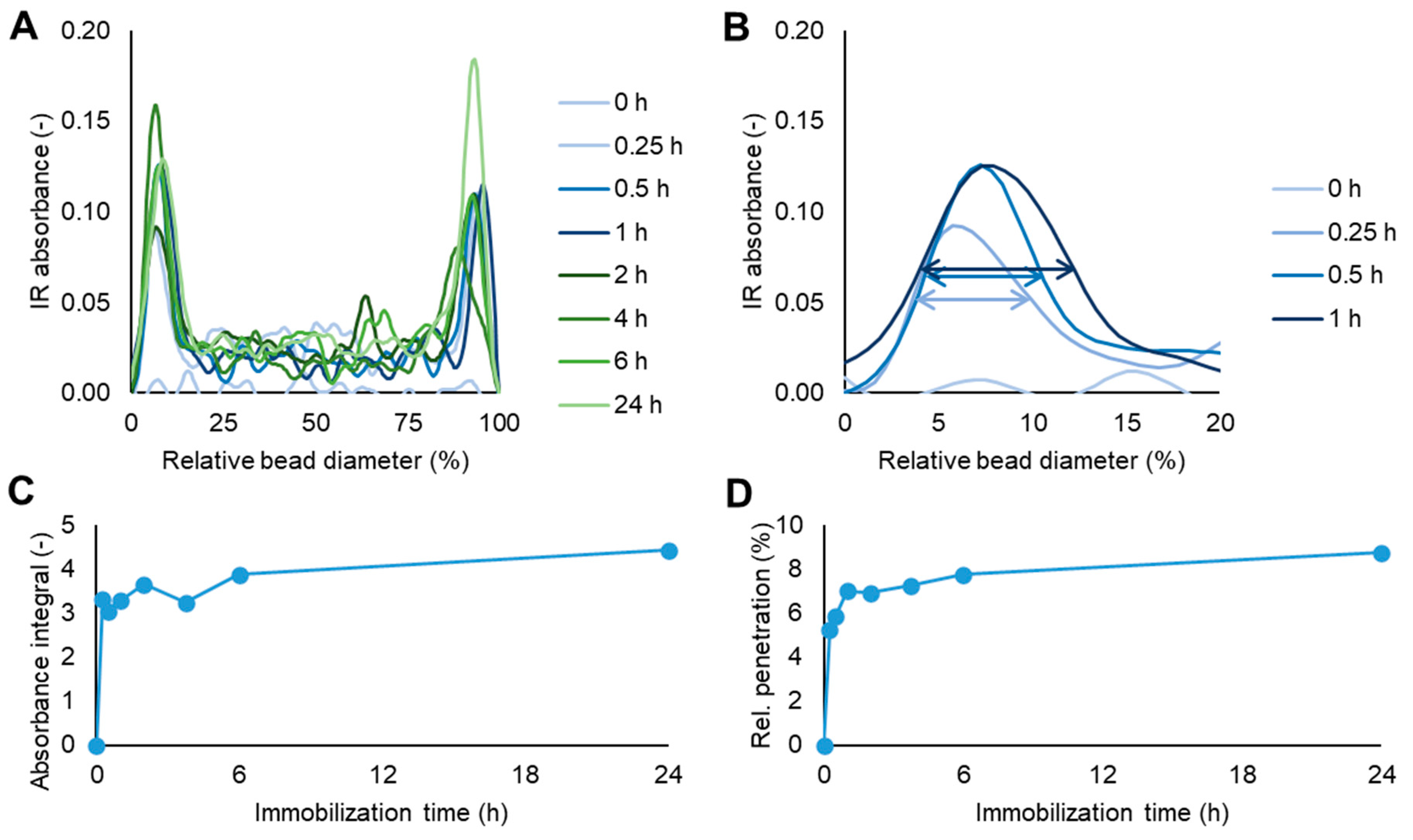
When CalB was immobilized on the hydrophobic resin ECR1030M the interaction proceeded in a completely different pattern. Although there was no linear correlation between the absolute or relative CalB penetration in ECR1030M depending on the bead size, the data obtained showed that the enzyme penetration within the first hour of immobilization was the most efficient, reaching approx. 30 µm (7%) from the surface of the beads, the additional immobilization time up to 6 h only increased the CalB migration inside the beads by another 10 µm, and the extended immobilization time to 24 h was only increasing the enzyme penetration to approx. 50 µm from the bead surface (
Figure 1). These FT-IR microscope values are a confirmation of the fast hydrophobic interaction between CalB and the enzyme carrier ECR1030M.
The findings of the IR measurements were correlated with the measurements of enzyme activity over time.
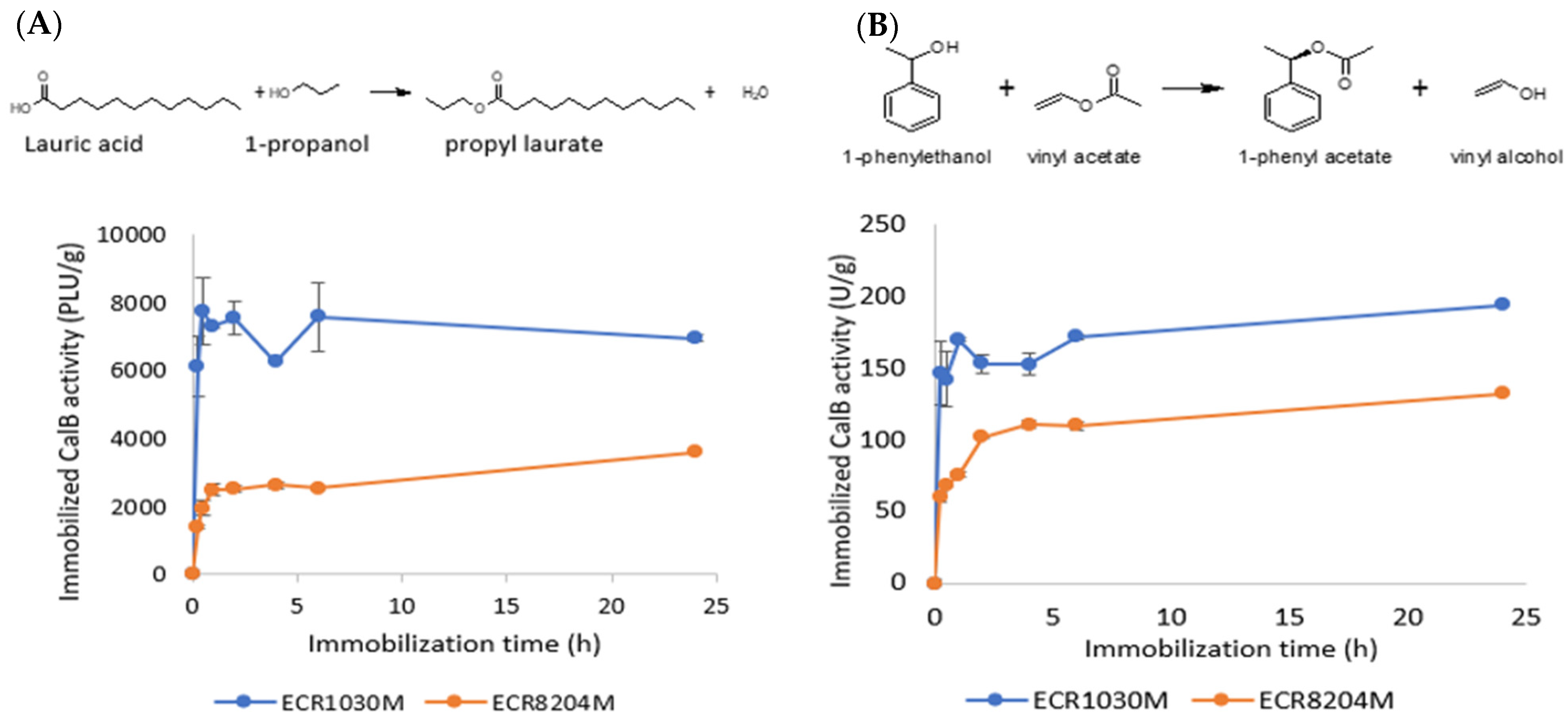
The enzyme activities on ECR1030M were always higher than on ECR8204M (3.2 ± 0.8 times higher in the PLU test and 1.8 ± 0.4 times higher in the 1-phenylethyl acetate test) for all immobilization times and activity test methods (
Figure 5). These results can be explained by analyzing the surface of the lipase CalB. In the paper from Basso et al. (2007) [
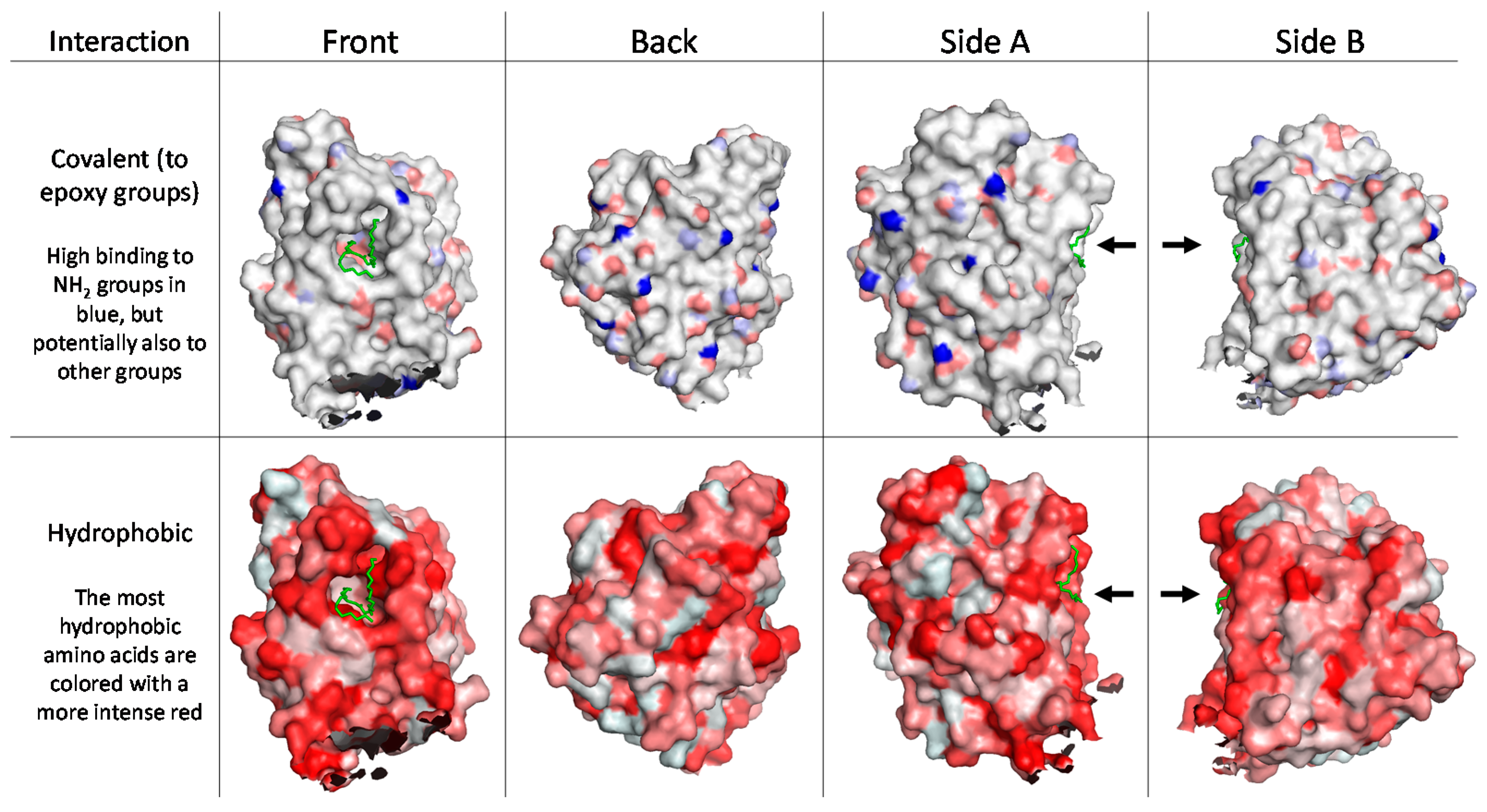
16] the computational analysis of the surface of CalB showed that although the interfacial activation through the “lid” opening was absent, the enzyme evolved to act at the lipid/water interface. This is clearly demonstrated by the distribution of the hydrophilic and hydrophobic regions on the protein surface (
Figure 8). In the case of CalB, it is interesting to note how a broad hydrophilic area is present opposite to the active site. This suggests that the orientation and even the conformation of CalB can change significantly when adsorbed on carriers that differ in hydrophilicity, confirming such behavior previously reported [
19,
20].
Moreover, the FT-IR and activity data supported the hypothesis that most of the activity came from the enzyme immobilized closer to the surface. For example, the loading front advanced in ECR8204M over the 24 h immobilization when the entire bead was almost saturated by the enzyme (
Figure 3A,C). However, the enzyme activities were quite constant from 1–2 h up to 24 h (
Figure 5) for CalB immobilized on both ECR1030M and ECR8204M. Moreover, despite the fact that CalB had a more uniform distribution throughout ECR8204M compared to ECR1030M, the enzyme activity was much higher on ECR1030M, indicating that the substrate and product diffusion through the diameter of the ECR8204M was not fast enough to give optimal activity. Additionally, when studying the potential binding sites at the surface of the enzyme towards the epoxy groups of the ECR8204M resin, it is likely that most of the enzyme is in a well-oriented form with many binding sites opposite to the catalytic center (
Figure 8). [
21] Nonetheless, the enzyme is covalently immobilized on ECR8204M, which renders the enzyme more rigid and could contribute to a lower activity. On the other hand, looking at the density of hydrophobic areas at the enzyme surface (
Figure 8), it is likely that the active center of CalB, when immobilized on ECR1030M, is not exposed towards the reaction media (
Figure 8); however, when CalB is in contact with hydrophobic media, it experiences the so-called interfacial activation [
22], leading to higher activities. Moreover, the hydrophobic immobilization typically leads to a more dynamic enzyme conformation and not as restricted as when covalently bound to the resin. Hence, even though the orientation of the enzyme on ECR8204M might be better than on ECR1030M, the activity is not optimal, regardless of the substrate/products bulkiness or type of reaction, which leads to the conclusion that for CalB, the interfacial activation may lead to a higher activity than a good enzyme orientation.
There was also a good correlation between the IR absorbance at the amide I band across the diameter of the beads (
Figure 1C and
Figure 3C) and the activities obtained (
Figure 5A,B) and it can be concluded that the immobilization on ECR1030M occurred within the first 30 min, while it took up to 2 h on ECR8204M. The apparent dip in enzyme activity at the 4 h timepoint on ECR1030M, seen in
Figure 5A, was also seen as a slightly lower amide I integral in
Figure 1C, suggesting these were not measurement artifacts, but rather a difference caused by the immobilization procedure or workup. These data show that FT-IR can be a meaningful tool not only to study the secondary structure of an immobilized enzyme, as recently seen by Mangiagalli et al. [
22], but also to study the enzyme distribution pattern within a support.
The fact that the CalB activity on ECR1030M was high using both substrates indicates that despite CalB not penetrating to the center of the particles, it is unlikely that the diffusion was hindered by size exclusion. The pore sizes of the particles used were more than 10 times larger than the size of CalB (30 Å × 40 Å × 50 Å) [
23], and this should not have been a limitation for CalB to diffuse throughout the carrier bead. This links to observations made by Mei et al. (2003) [
12], where it was proved that despite the enzyme being located on the surface of the particle, there were larger molecules that could diffuse across the beads.
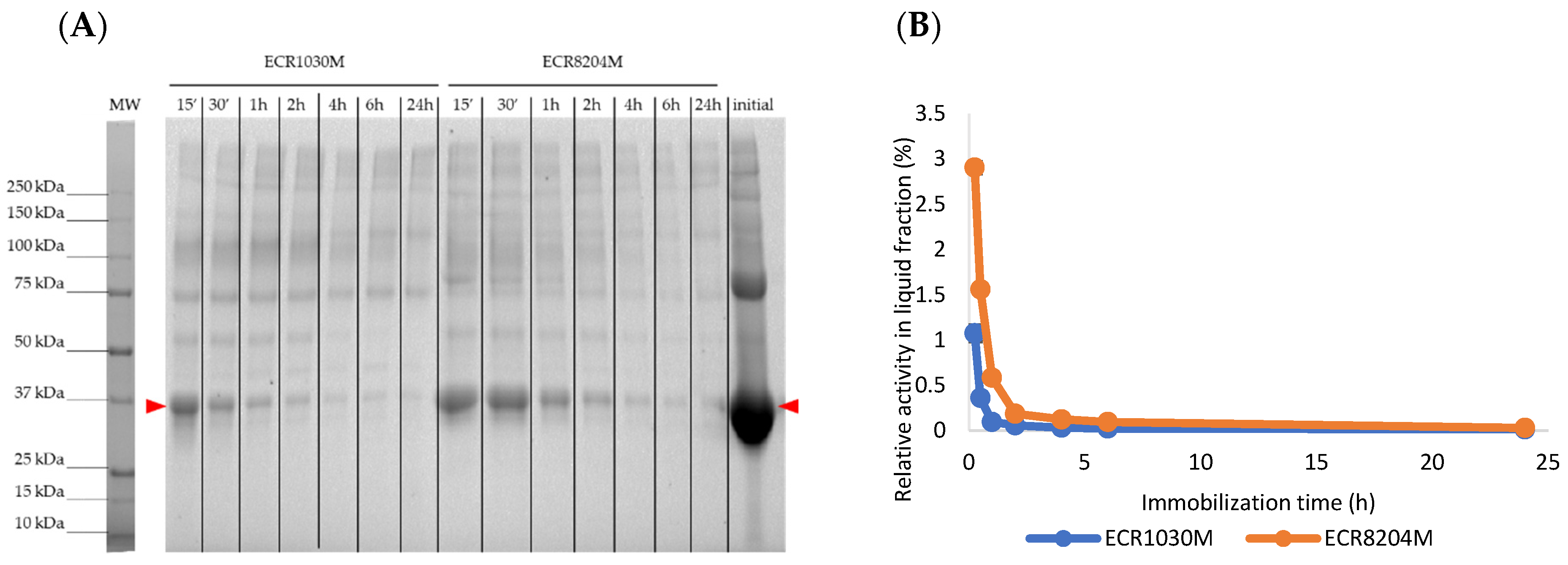
As shown by the SDS-PAGE analysis after the immobilizations (
Figure 6), there was a higher CalB affinity for ECR1030M at initial immobilization times, but after 2 h, a very similar amount of enzyme was immobilized in both resins; this somehow correlates with the rather constant activities after 1–2 h found in
Figure 5.
The results obtained in this work confirm previous reports describing immobilization mechanism and enzyme distribution patterns [
24,
25,
26,
27,
28]. In this way, the enzyme diffuses into the particle forming a loading front with a high enzyme concentration, where most of the times it does not reach the center of the porous material. In order to fully load the particles, long interaction times are needed [
14], or a competing compound could be used [
28,
29]. Generally, if the distribution is nonuniform (U-shaped, with the enzyme concentrated on the surface), the activities are higher than if the enzyme is well distributed along the particle diameter. However, a more uniform distribution is more suitable for reactions that suffer from inhibition kinetics [
26,
27]. Thus, for CalB, if there is no inhibition, a short immobilization time is recommended. This would be approx. 6 h for ECR8204M and 1 h for ECR1030M, as suggested by the enzyme distribution profile (
Figure 1 and
Figure 2) and the activity data (
Figure 5) and SDS-PAGE analysis (
Figure 6).
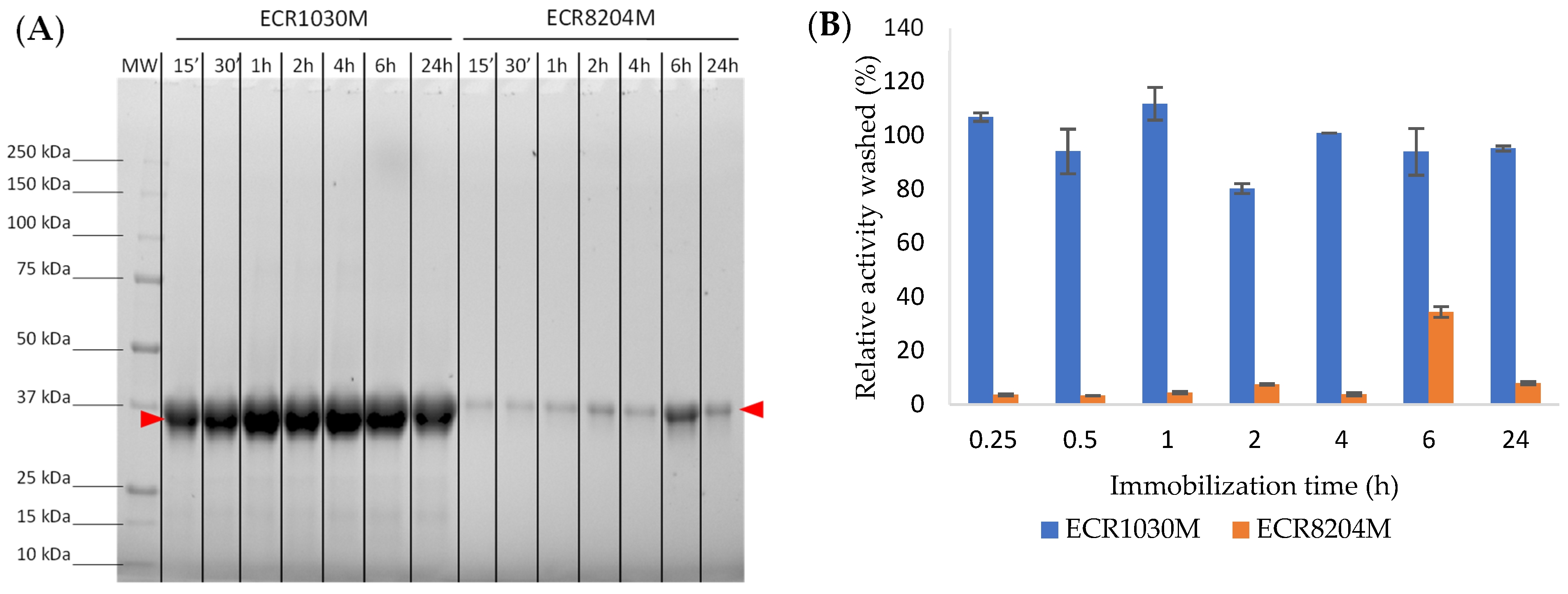
Another important aspect that we wanted to confirm was the type of interaction between the CalB and each enzyme carrier which was done by treating the immobilized enzyme samples with a surfactant (
Figure 7). The leaching of CalB from ECR1030M happened in the presence of the anionic surfactant Triton X-100 that was able to disrupt the hydrophobic interactions between the aromatic groups of the resin and the surface of the enzyme. Since such interaction could lead to a decrease in enzyme activity when used over multiple cycles, we verified the reusability of the preparation. In a separate experiment (data not shown), we observed that CalB immobilized on ECR1030M was very robust and could be recycled for multiple cycles without any loss of enzyme activity (99% recovered activity over 10 cycles using PLU test). Moreover, the commercial preparation of CalB immobilized on ECR1030M (CalB immo Plus) is also used in many industrial applications for multiple cycles and long times, one example being published by Brown et al. [
30].
4. Materials and Methods
All chemicals were laboratory grade and of suitable purity and were purchased from Sigma Aldrich (Merck), UK. Enzyme lipase CalB was provided in liquid form by c-Lecta GmbH (Leipzig, Germany). Embedding and microtomy supplies were purchased from Biosystems AG, Switzerland.
4.1. Characterization of Enzyme Carriers
The enzyme carriers ECR1030M and ECR8204M were of synthetic backbone, produced and provided by Purolite Llantrisant, UK and are commercially available. The pore diameter and porosity were measured by mercury intrusion using Thermo Scientific Pascal 140 and Thermo Scientific Pascal 240 instruments, Waltham, MA, USA. Particle sizes were determined by dynamic image analysis using QICPIC instrument by Sympatec GmbH, Ramlingen, Germany. The total moisture of the enzyme carriers and immobilized enzymes was measure using an IR balance set at 105 °C (Sartorius).
4.2. Enzyme Immobilization in Batch Mode
Two enzyme immobilization techniques were used as described below. For all preparations, the initial enzyme loading was at 2500 pNPB units per gram of dry resin. After immobilization, the supernatants were collected by filtration for protein and activity analysis. The immobilization procedures are described at 1 g scale, but the same procedures were used to make tens of grams of immobilized products by proportionally scaling up. All immobilizations were performed at room temperature (RT), under rotary mixing (20 rpm) (Stuart rotator). The immobilization time varied from 15 min to 24 h.
In the covalent immobilization on an epoxy functionalized resin, the final enzyme solution containing 0.5 M potassium phosphate, a pH 7.0 buffer was mixed in a ratio of 1.5:1 with the ECR8204M enzyme carrier for specified periods of time.
In the hydrophobic immobilization on nonfunctionalized resin, the final enzyme solution containing 0.02M potassium phosphate, a pH 7.0 buffer was mixed in a ratio of 1:1 with the ECR1030M enzyme carrier for specified periods of time.
The liquid obtained after each immobilization was analyzed by SDS-PAGE to qualitatively identify the presence or absence of CalB and compared to the initial CalB solution used for immobilization. For this purpose, a Mini-PROTEAN vertical electrophoresis instrument from Bio-Rad, Watford, UK was used following the method described by Laemmli (1970) [
31].
Following the immobilization, the carriers were washed 4 times with demineralized water in a ratio resin/water of 1:4 (wt:wt). The immobilized enzyme was then filtered to remove excess liquid and stored at 2–8 °C.
4.3. Enzyme Activity Assays
To quantify the activity of the immobilized enzyme, two different test methods were used.
The analysis of the CalB-catalyzed ester formation (PLU activity) was performed using immobilized CalB and following the same method as described by Basso et al. (2016) [
32]. PLU refers to the amount (µmol) of propyl laurate synthesized by the enzyme per minute under the reaction conditions.
The performance of the immobilized CalB (100 mg) was also assessed following the transesterification between 1-phenylethanol (0.6 mL, 4.97 mmol) and vinyl acetate (2.4 mL, 26 mmol) at 25 °C, in the absence of solvents under vigorous mixing over 30 min reaction time. Briefly, 10 µL of the reaction mixture was diluted with 990 µL of acetonitrile and the formation of 1-phenylethyl acetate was quantified by HPLC using a Columbus C18, 5 µm 110A, 250 mm × 4.6 mm column at 25 °C, mobile phase: 60:40 of H2O:CH3CN, 1 mL/min, λ = 260 nm.
The esterase activity of free CalB, from liquid samples before and after the immobilizations and after the Triton X-100 wash, was measured using 2 mM p-nitrophenyl butyrate (pNPB) as a substrate in 0.1% Triton X-100, 50 mM potassium phosphate buffer, pH 8.0. The formation of p-nitrophenol was followed at 405 nm for 1 min. One pNPB unit is defined as micromoles of p-nitrophenol produced per minute at room temperature at pH 8.0.
All activity measurements were done in duplicate.
Following the CalB immobilization, with the purpose to confirm whether the enzyme was attached to the beads covalently or by hydrophobic interactions, 50 mg of immobilized preparations was incubated with 0.5 mL 1% Triton X-100 for 2 h at RT. Then, the liquid fractions were collected and analyzed by SDS-PAGE.
4.4. Paraffin Embedding Procedure
A small quantity of sample was taken out of the original container (stored at 5 °C) and allowed to reach room temperature prior to embedding. The single-use polyethylene molds (15 × 15 × 6 mm, Biosystems AG) used for embedding were preheated to approximately 60 °C in the embedding workbench (Thermo Microm EC 350-1, Thermo Scientific, Germany). Molten paraffin wax (Leica Paraplast Plus, Leica Biosystems, Germany, melting point 58 °C) was poured approx. 2 mm high into a preheated mold. Onto this bed of paraffin, a small amount of sample was dropped with a single-use spatula. The mass was allowed to cool and solidify partially for approximately 30 s, before the embedding cassette (40 × 28 × 6 mm, Biosystems AG) was placed on top of the mold and the mold filled completely with paraffin. The filled molds were left to harden at 5 °C for a minimum of 4 h and kept at this temperature until further processing.
4.5. Microtome Slicing Procedure
The solidified paraffin blocks were allowed to reach room temperature prior to cutting. The blocks were removed from the molds and cut into 10 µm thickness slices using a rotary microtome with section transfer system (Carl Zeiss Hyrax M40 with Epredia STS, Carl Zeiss, Germany). The section transfer system is filled with ultrapure water at 35 °C and enables a gentle collection of paraffin slices through a type of water slide. The 3D-printed microscope slides were dipped into the collection pool of the section transfer system to mount the paraffin sections floating therein. The hydrophobic nature of the paraffin and the slides led to a quick and secure attachment. The adhering water was left to evaporate, and the so-prepared samples were stored at room temperature.
4.6. Spectra Collection
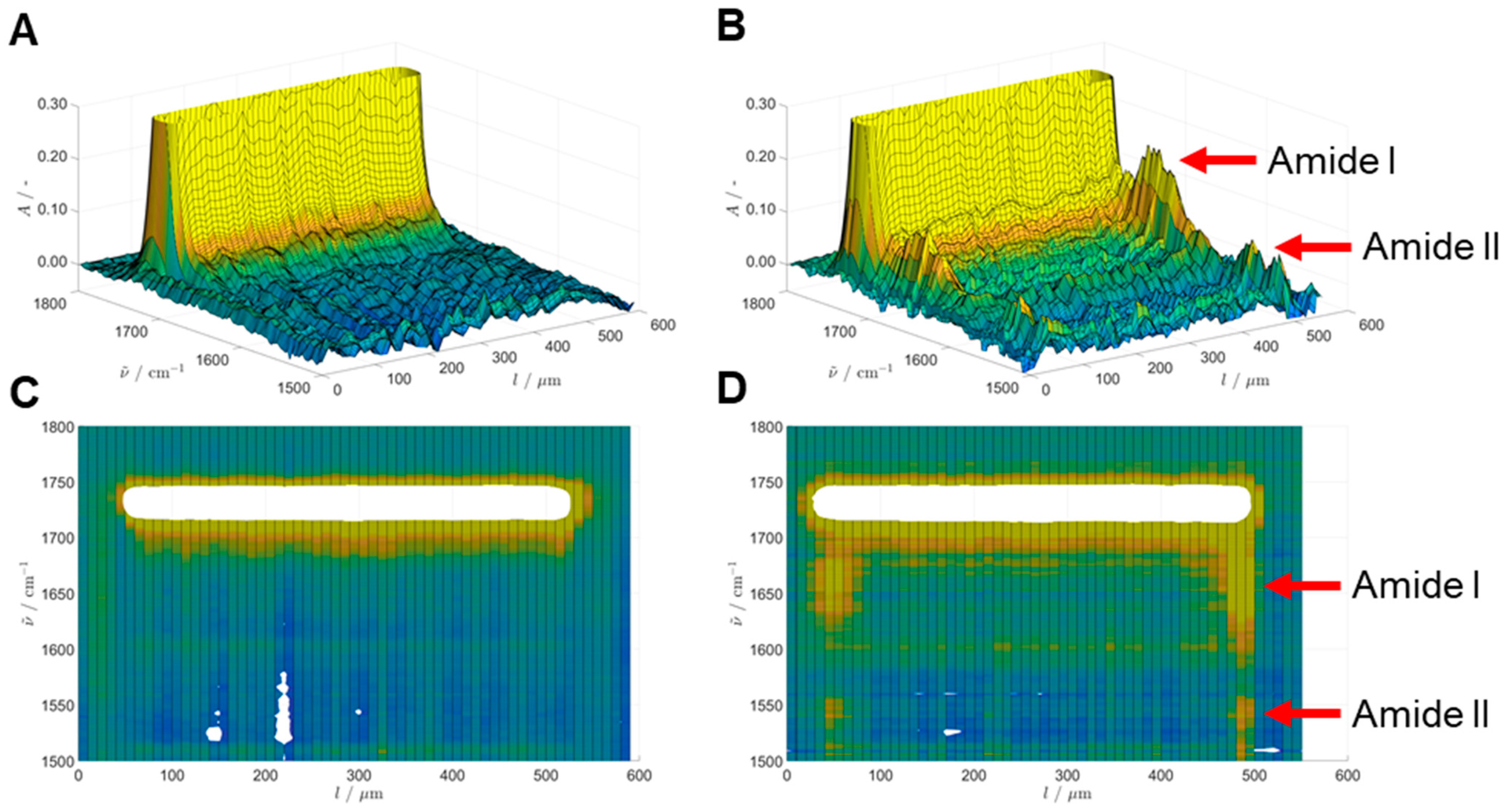
The samples were analyzed using a PerkinElmer Spotlight 200 FT-IR microscope (PerkinElmer, Waltham, MA, USA). The microscope was operated in transmission mode with the rectangular aperture set to 10 µm × 10 µm. The spectral range was set to 4000–600 cm−1 with a resolution of 4 cm−1. A total of 256 integrations were acquired for all measurements. The measurements were programmed using PerkinElmer Spectrum (Version 10.5.1.581, PerkinElmer, Waltham, MA, USA, 2015), using the “Line Scan” feature to draw a line of adjacent squares across the diameter of the particle section as observed in visible-light mode. The spectra of these 10 µm × 10 µm squares was then automatically acquired in sequence, using the motorized high-precision sample positioning table of the microscope. An air background was measured prior to every line scan sequence.
4.7. Data Analysis
The raw data files (x- and y-location, wavenumber, absorbance) were processed using RStudio (Version 2021.09.1, RStudio, Boston, MA, USA, 2021), MATLAB (Version R2020a, The MathWorks, Natick, MA, USA, 2020), and Microsoft Excel (Version 2108, Microsoft, Redmond, WA, USA). The presence of enzyme was monitored through observation of the amide I band at 1658 cm
−1, while a single point baseline was taken at 1800 cm
−1. Information regarding the calculation can be found in the
supporting information (SI).
4.8. Three-Dimensional Printing of Microscope Slides
The microscope slides were designed by O.P. and printed in-house with a commercial Ultimaker S5 (Ultimaker, Utrecht, the Netherlands) fused deposition modelling (FDM) 3D printer using clear polylactide (PLA) filament purchased from Ultimaker as feedstock. The slides had the same outside dimensions as commercial glass microscope slides. Three evenly spaced holes with a diameter of 5 mm allowed the unhindered passage of IR radiation while sufficiently supporting the paraffin sections. The 3D model is available in the
SI.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}