
Immobilization of Recombinant Endoglucanase (CelA) from Clostridium thermocellum on Modified Regenerated Cellulose Membrane


Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of Cellulase Gene in Plasmid
2.2. Recombinant Cellulase Expression under Different Conditions
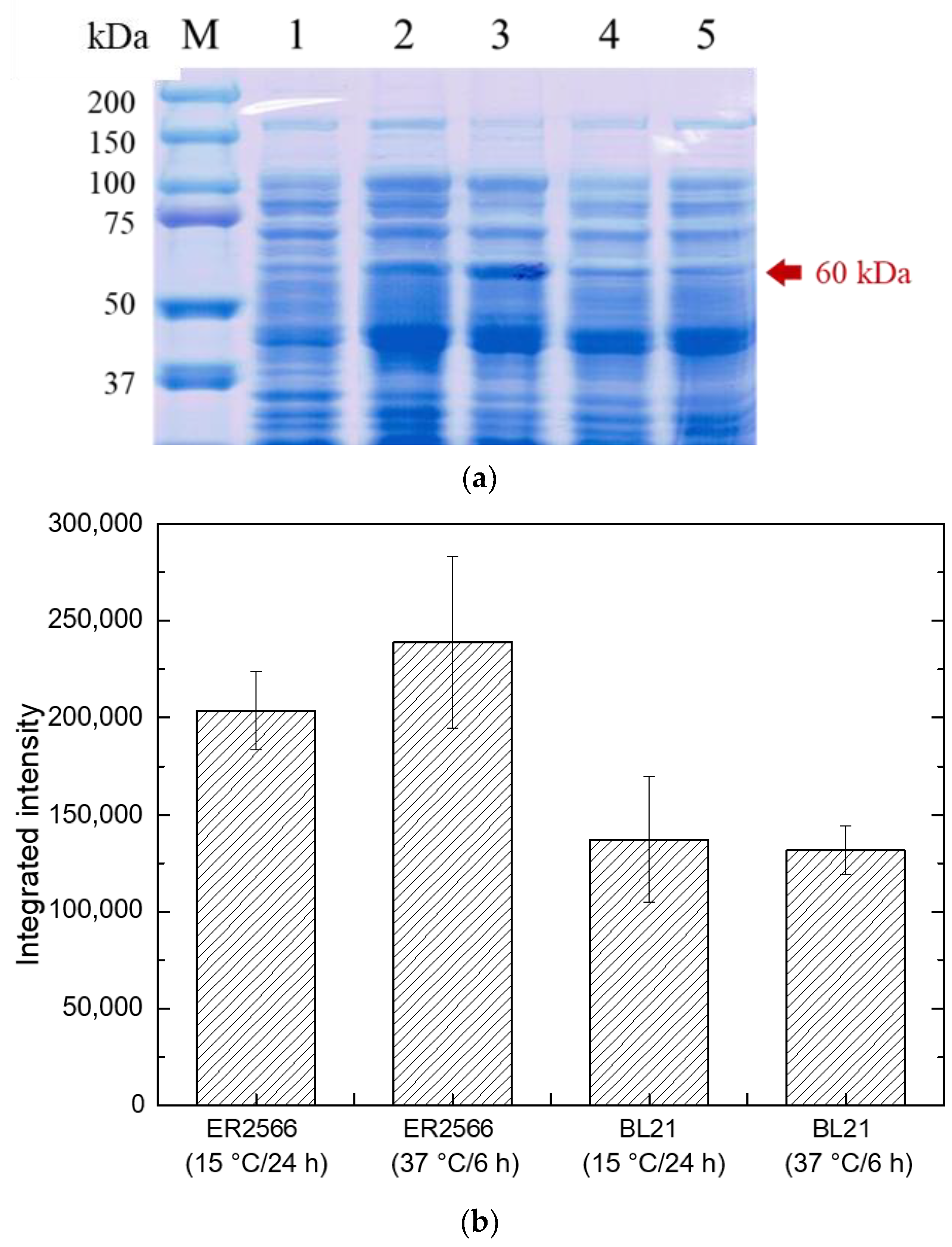
2.2.1. Effect of Different Hosts and Temperatures
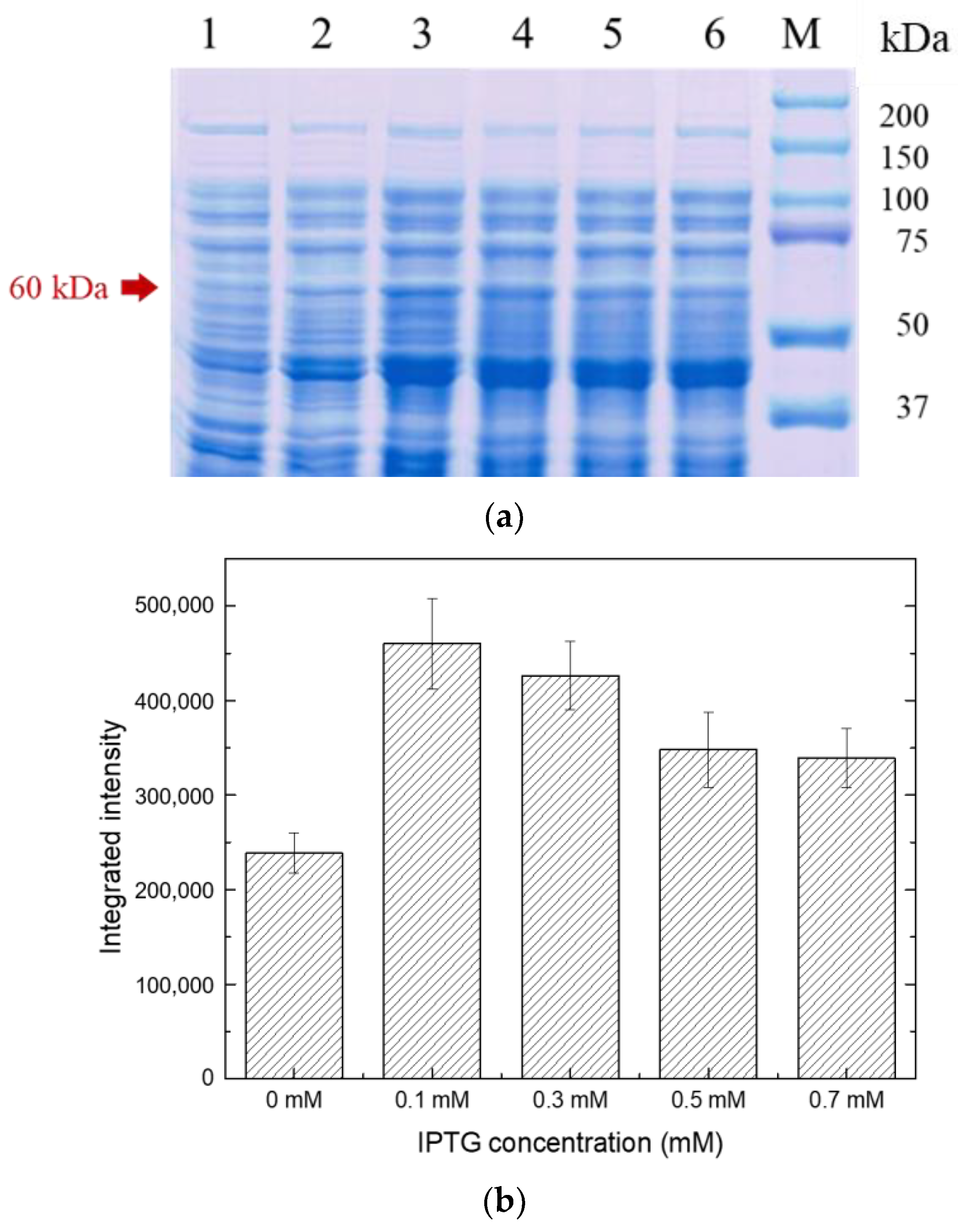
2.2.2. Effect of Different Inducer Concentrations
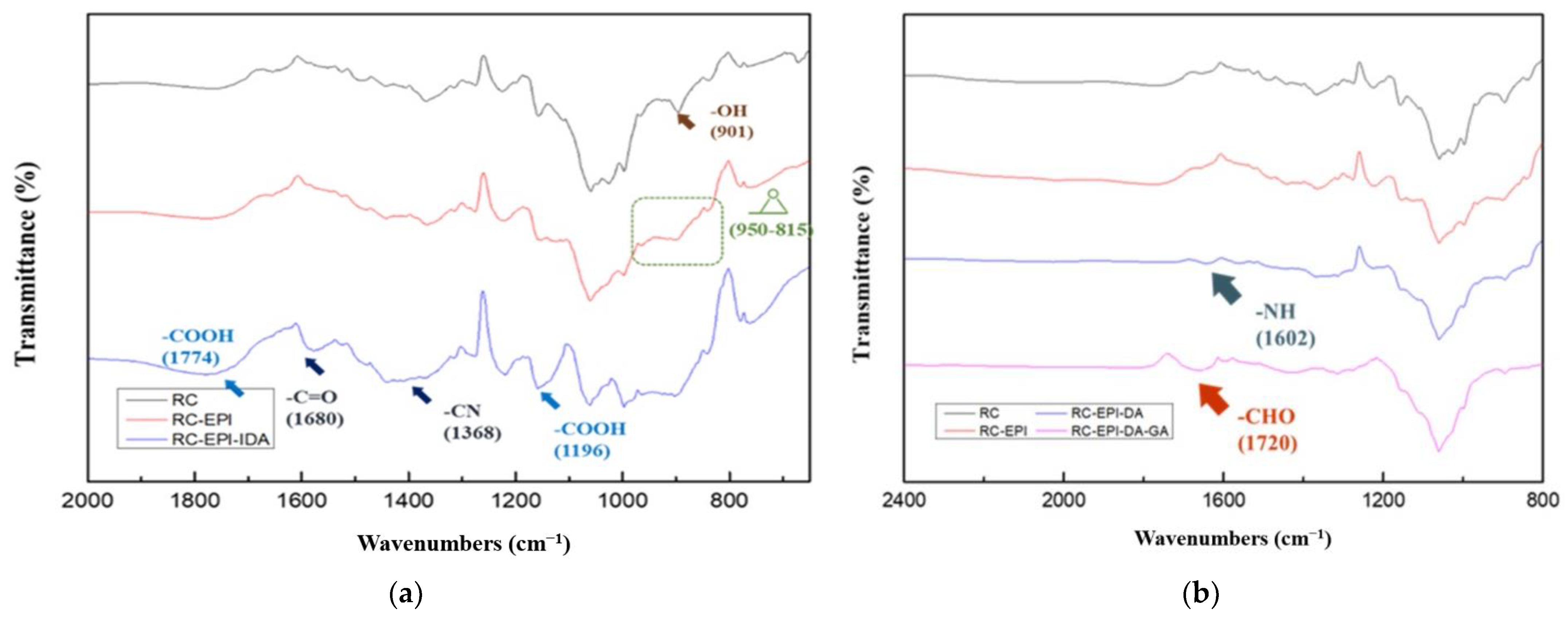
2.3. Characterization of Modified RC Membranes
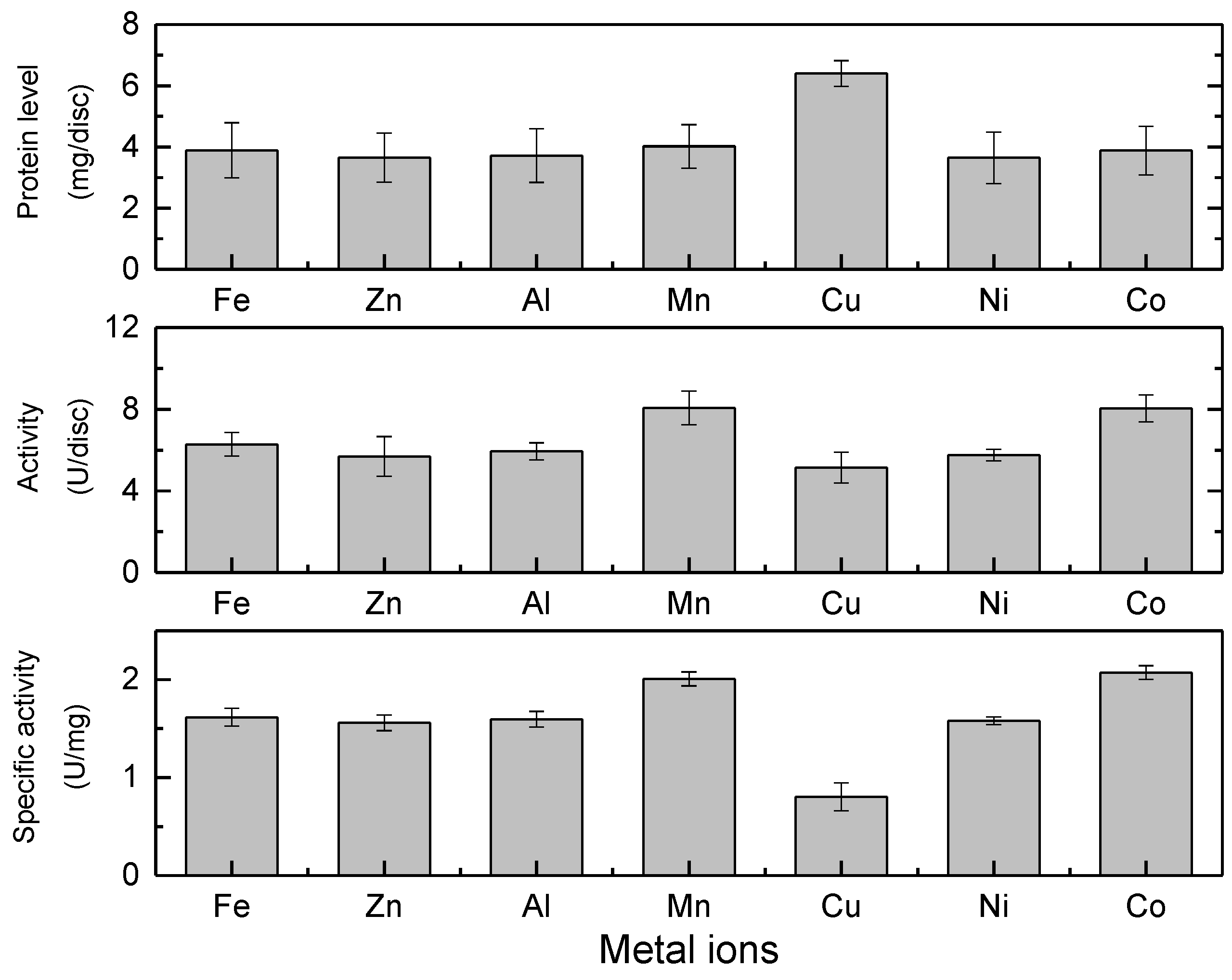
2.4. Metal Ion Selection for IMAM Chelation and Effect on Enzyme Activity
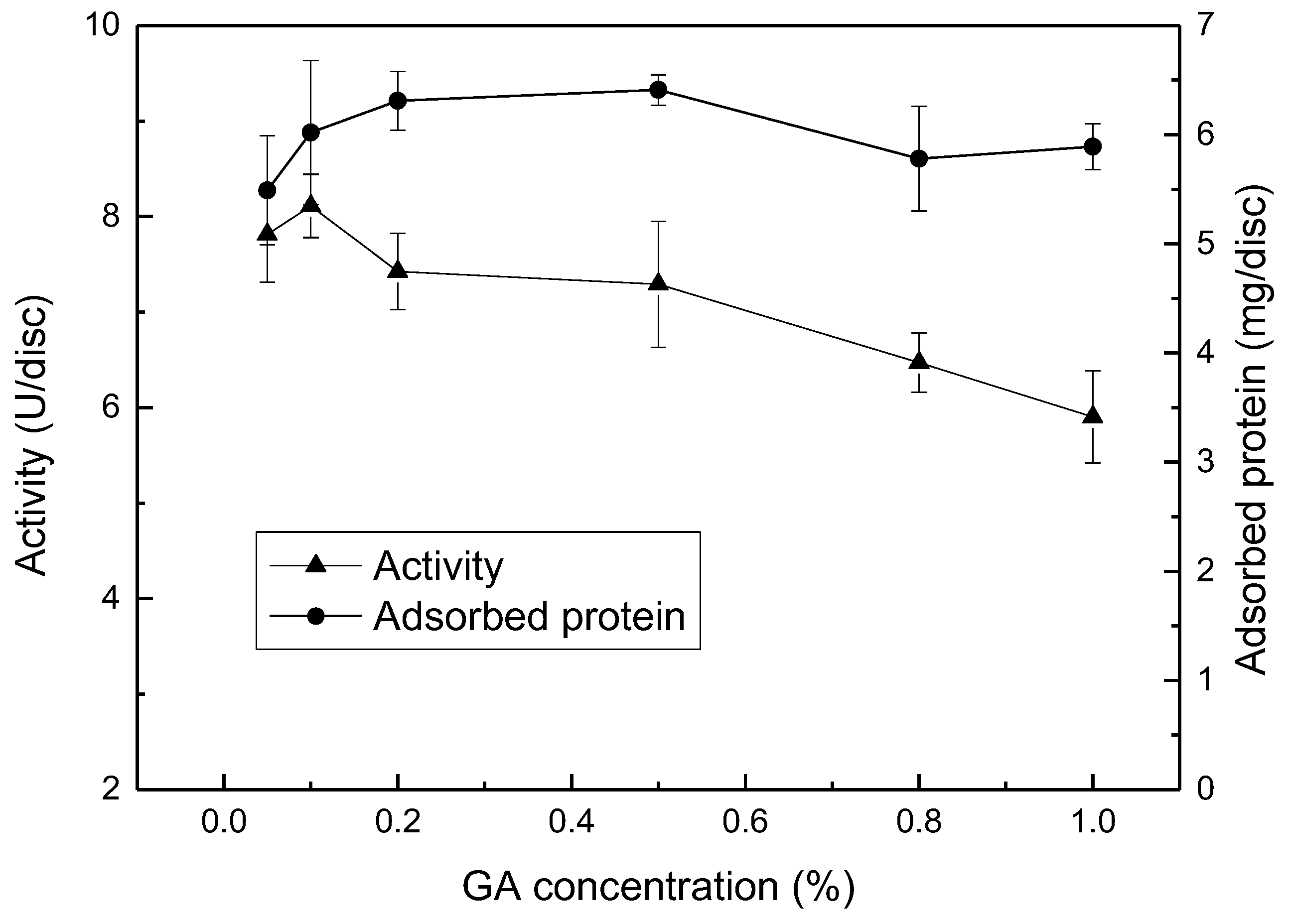
2.5. Effect of GA Concentration and Treatment Time
2.6. Purification Analysis
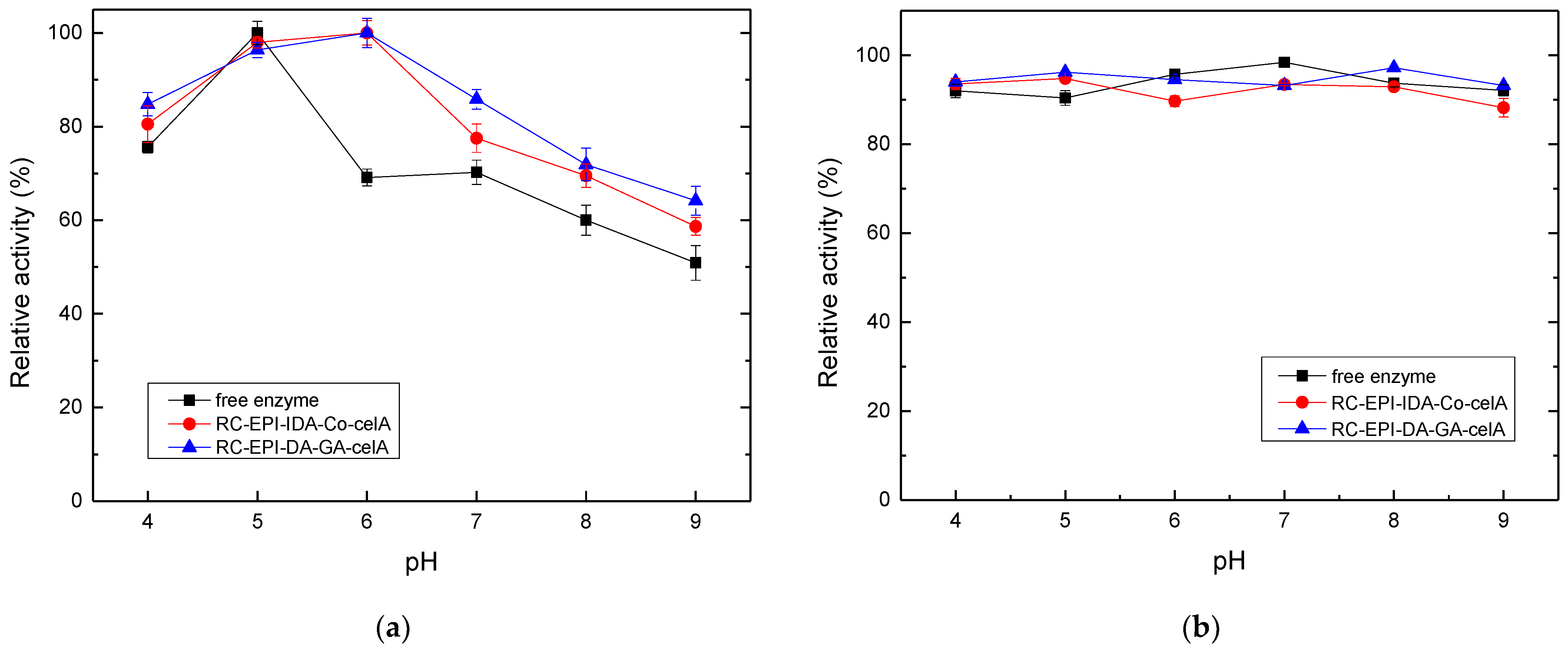
2.7. Optimal pH and Enzyme Stability
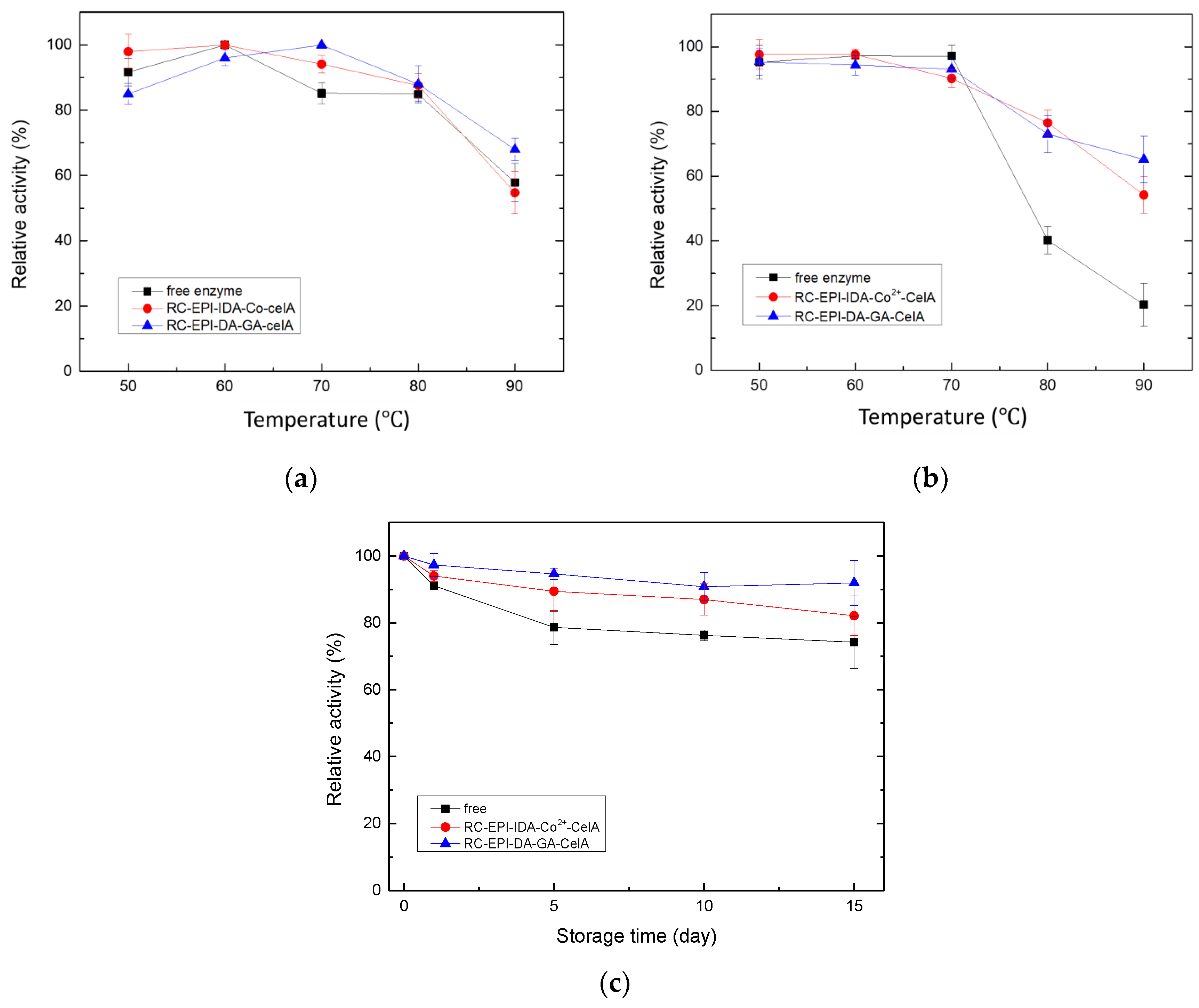
2.8. Optimal Temperature and Thermo-Stability
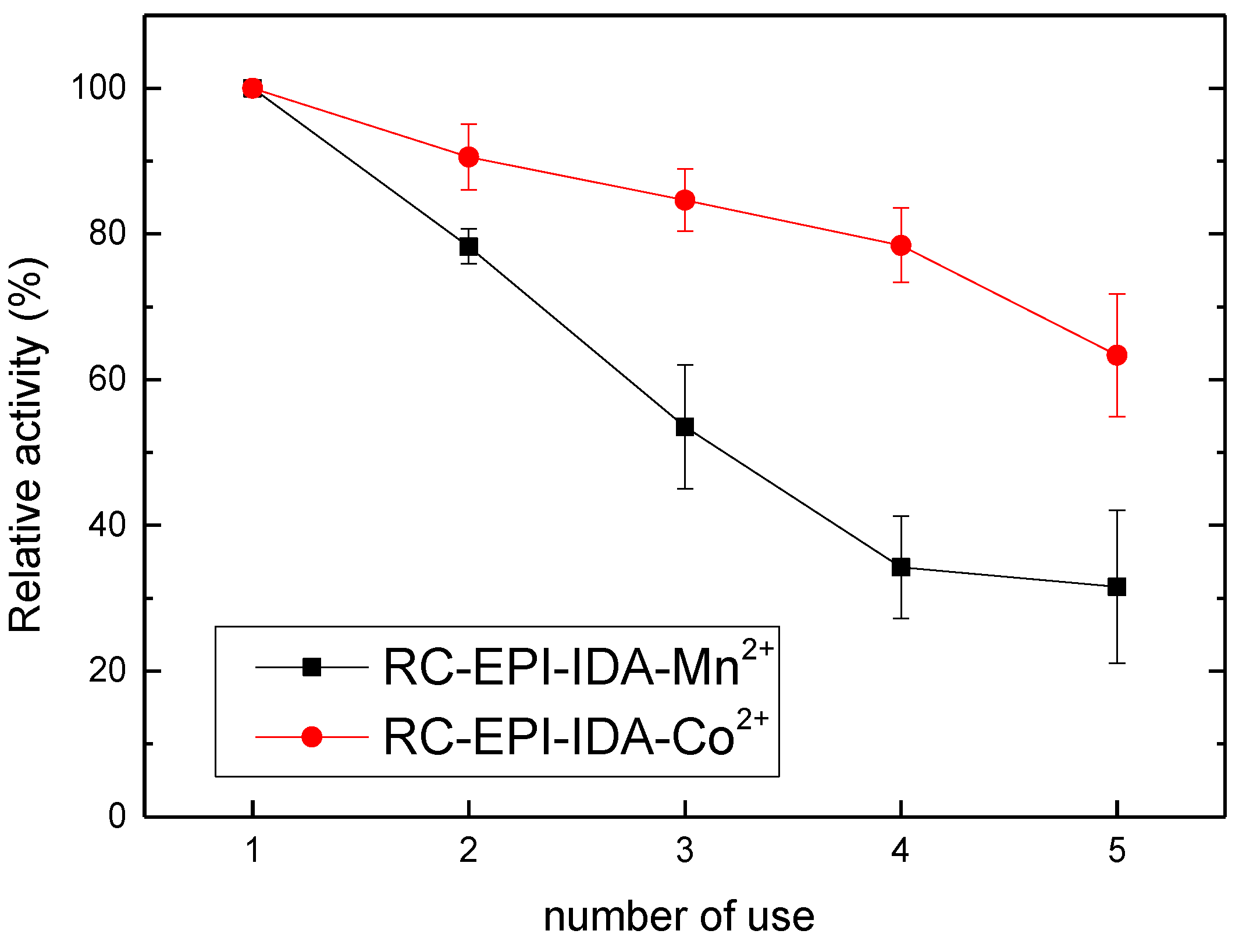
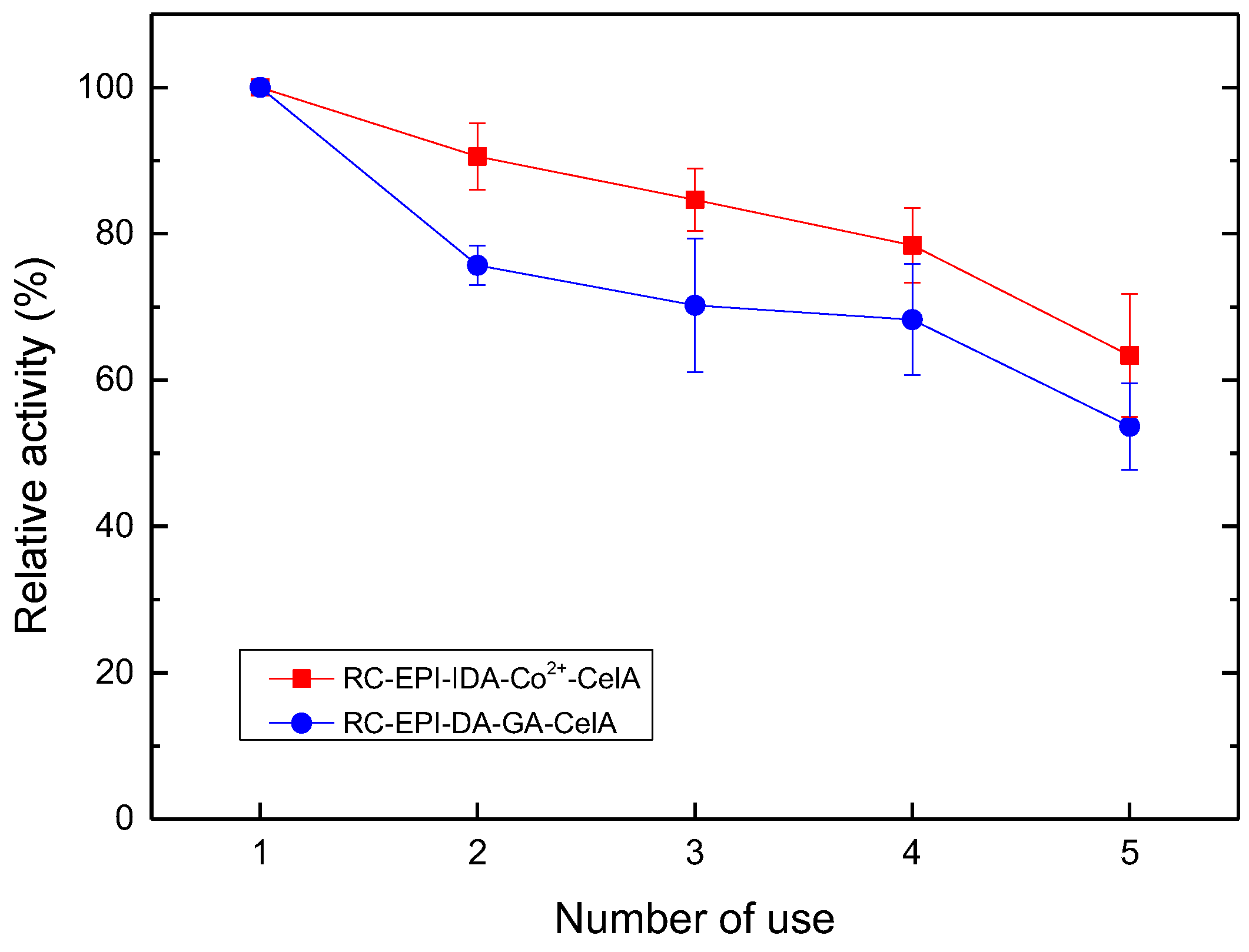
2.9. Enzyme Reusability Analysis
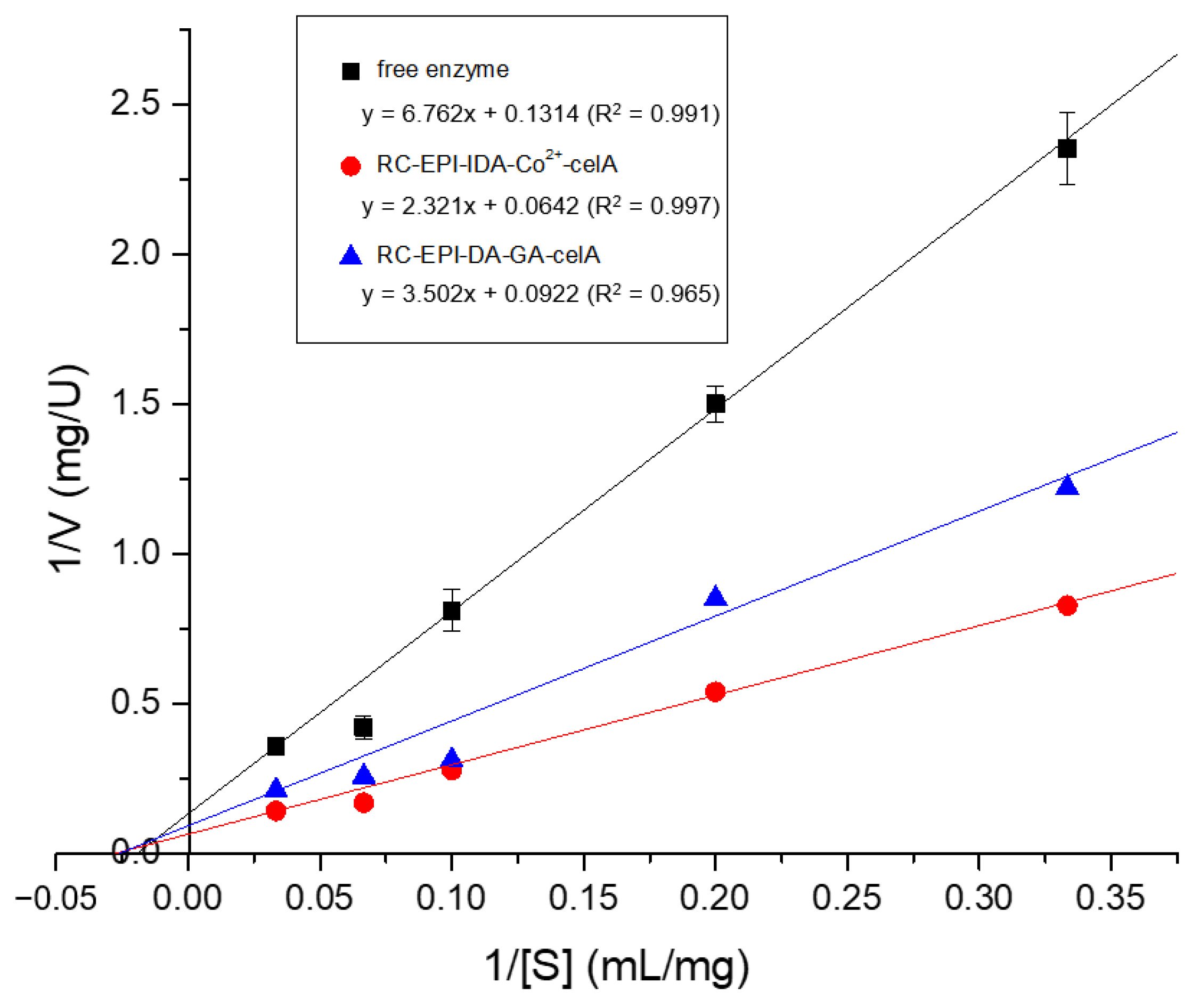
2.10. Kinetics Study of Free and Immobilized Enzymes
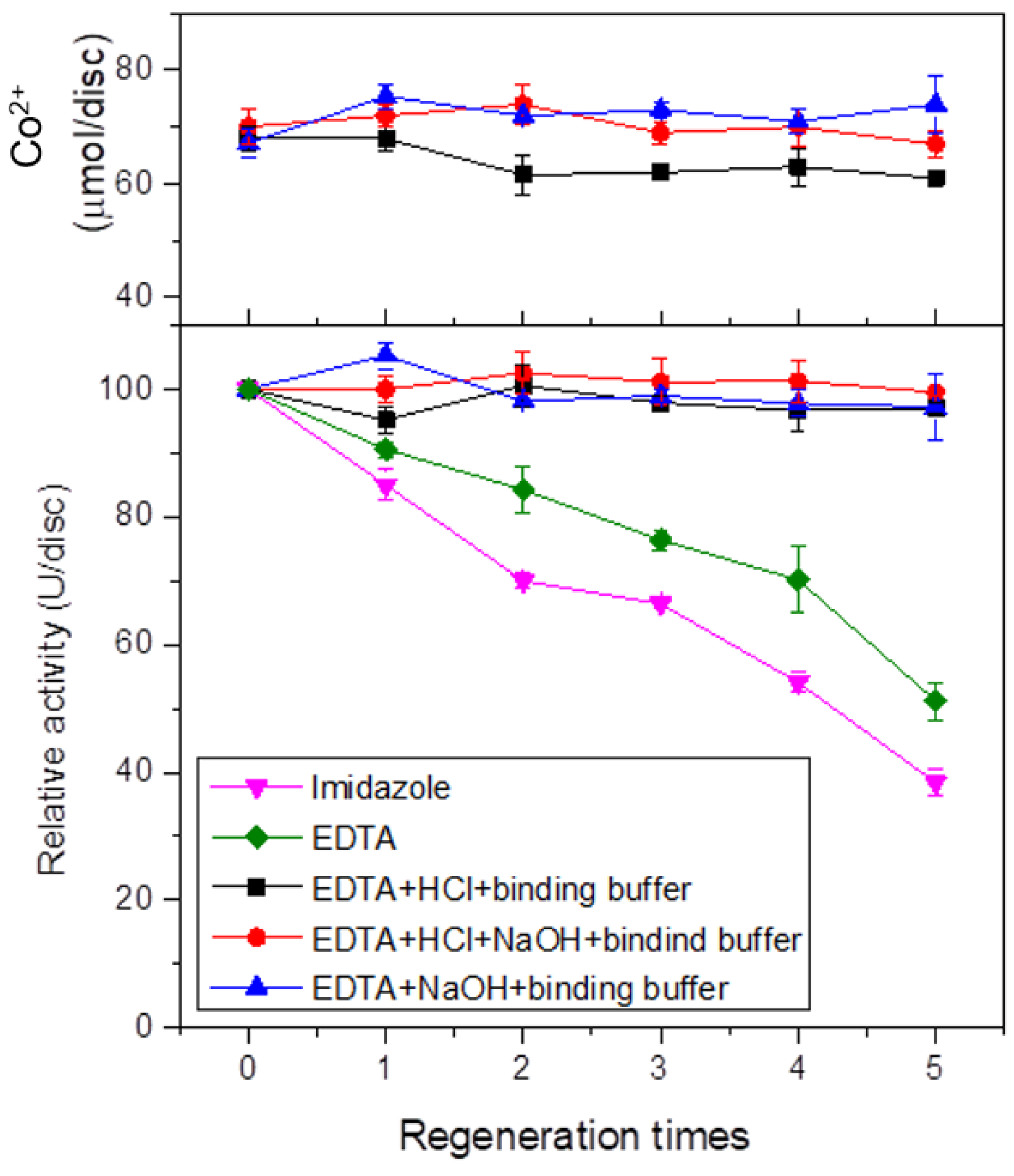
2.11. Regeneration of IMAM
3. Materials and Methods
3.1. Chemicals, E. coli Strains, and Plasmid Sources
3.2. Gene Cloning and E. coli Culture
3.3. Gene Expression and Enzyme Extraction
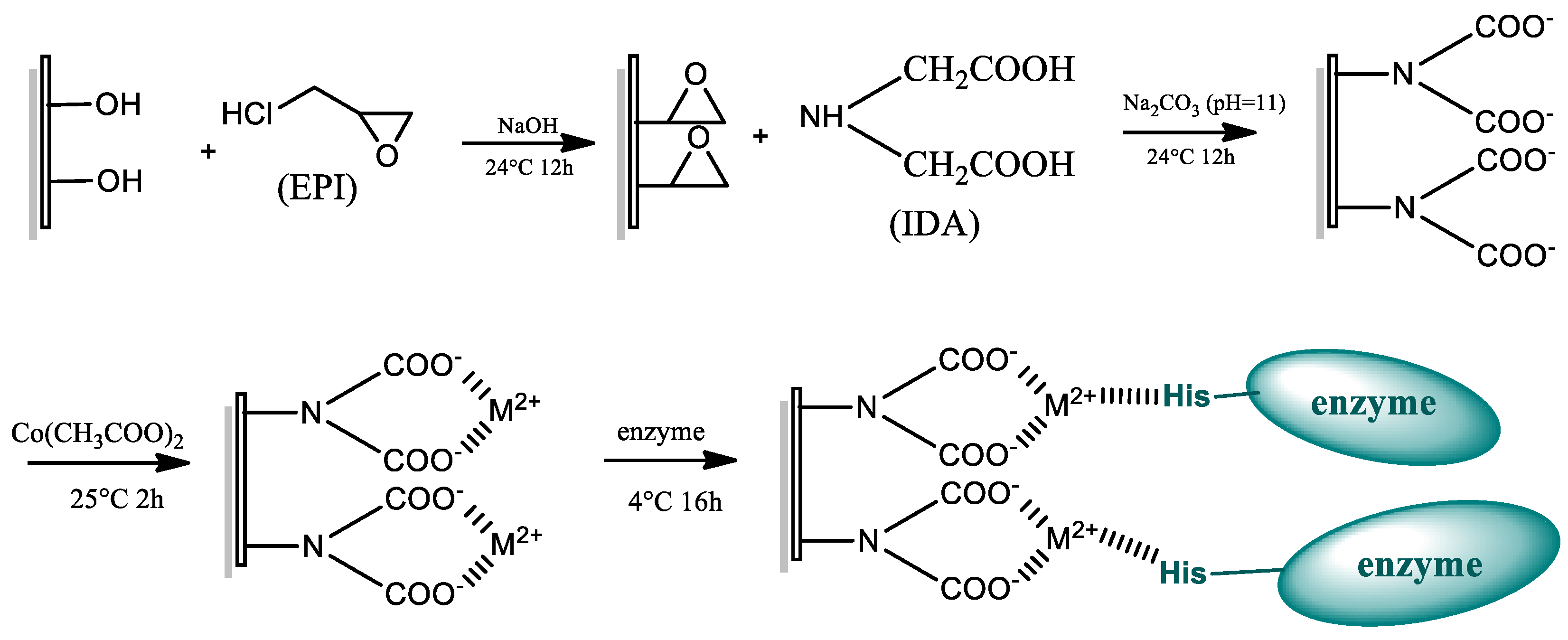
3.4. Preparation of IMAMs and Enzyme Immobilization
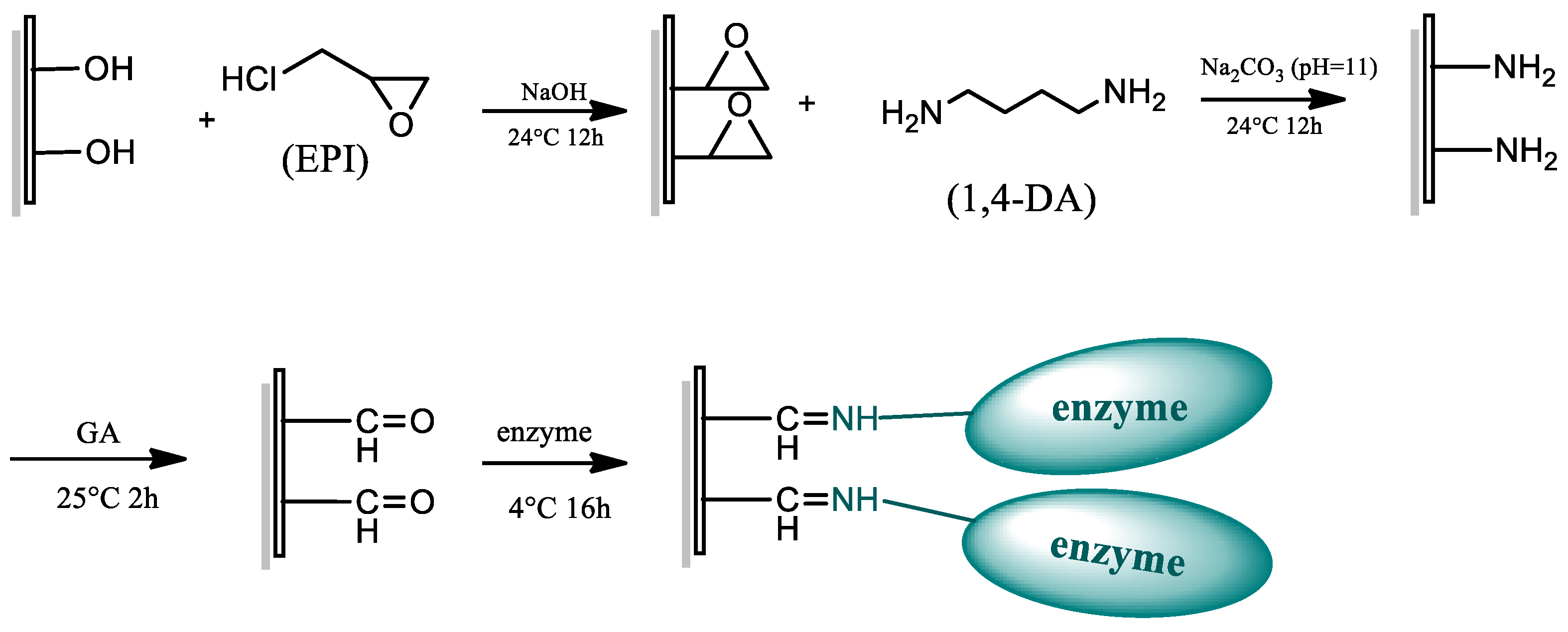
3.5. Preparation of RC-EPI-DA-GA Membranes and Enzyme Immobilization
3.6. Characterization of Thin Films by FTIR Analysis
3.7. Assays
3.8. Effect of pH and Temperature
3.9. Kinetic Analysis
3.10. Reusability Study for Immobilized CelA
3.11. IMAM Regeneration
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sharma, V.; Tsai, M.-L.; Chen, C.-W.; Sun, P.-P.; Patel, A.K.; Singhania, R.R.; Nargotra, P.; Dong, C.-D. Deep eutectic solvents as promising pretreatment agents for sustainable lignocellulosic biorefineries: A review. Bioresour. Technol. 2022, 360, 127631. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.K.; Kalia, V.C.; Joo, J.B.; Kang, Y.C.; Lee, J.K. Biotransformation of methane into methanol by methanotrophs immobilized on coconut coir. Bioresour. Technol. 2020, 297, 122433. [Google Scholar] [CrossRef] [PubMed]
- Nargotra, P.; Sharma, V.; Bajaj, B.K. Consolidated bioprocessing of surfactant-assisted ionic liquid-pretreated Parthenium hysterophorus L. biomass for bioethanol production. Bioresour. Technol. 2019, 289, 121611. [Google Scholar] [CrossRef] [PubMed]
- Amjith, L.; Bavanish, B. A review on biomass and wind as renewable energy for sustainable environment. Chemosphere 2022, 293, 133579. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Nargotra, P.; Sharma, V.; Bangotra, R.; Kaur, M.; Kapoor, N.; Paul, S.; Bajaj, B.K. Nanobiocatalysts for efficacious bioconversion of ionic liquid pretreated sugarcane tops biomass to biofuel. Bioresour. Technol. 2021, 333, 125191. [Google Scholar] [CrossRef] [PubMed]
- Kumari, D.; Singh, R. Pretreatment of lignocellulosic wastes for biofuel production: A critical review. Renew. Sustain. Energy Rev. 2018, 90, 877–891. [Google Scholar] [CrossRef]
- Vasić, K.; Knez, Ž.; Leitgeb, M. Bioethanol production by enzymatic hydrolysis from different lignocellulosic sources. Molecules 2021, 26, 753. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Lee, C.-K. Enhanced enzymatic hydrolysis of sugarcane bagasse by N-methylmorpholine-N-oxide pretreatment. Bioresour. Technol. 2009, 100, 866–871. [Google Scholar] [CrossRef]
- Patel, S.K.; Singh, M.; Kumar, P.; Purohit, H.J.; Kalia, V.C. Exploitation of defined bacterial cultures for production of hydrogen and polyhydroxybutyrate from pea-shells. Biomass Bioenergy 2012, 36, 218–225. [Google Scholar] [CrossRef]
- Vadala, B.S.; Deshpande, S.; Apte-Deshpande, A. Soluble expression of recombinant active cellulase in E. coli using B. subtilis (natto strain) cellulase gene. J. Genet. Eng. Biotechnol. 2021, 19, 1–7. [Google Scholar] [CrossRef]
- Hosseini, S.H.; Hosseini, S.A.; Zohreh, N.; Yaghoubi, M.; Pourjavadi, A. Covalent immobilization of cellulase using magnetic poly (ionic liquid) support: Improvement of the enzyme activity and stability. J. Agric. Food Chem. 2018, 66, 789–798. [Google Scholar] [CrossRef]
- Gennari, A.; Führ, A.J.; Volpato, G.; de Souza, C.F.V. Magnetic cellulose: Versatile support for enzyme immobilization—A review. Carbohydr. Polym. 2020, 246, 116646. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Patel, S.K.; Gupta, R.K.; Otari, S.V.; Gao, H.; Lee, J.K.; Zhang, L. Enhanced saccharification and fermentation of rice straw by reducing the concentration of phenolic compounds using an immobilized enzyme cocktail. Biotechnol. J. 2019, 14, 1800468. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, W.; Yang, Z.; Yang, X.; Wang, N.; Yu, X. Novel magnetic cross-linked cellulase aggregates with a potential application in lignocellulosic biomass bioconversion. Molecules 2017, 22, 269. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-H.; Liu, Y.-C.; Chang, C.-M.J.; Chen, J.-H.; Chang, C.; Shieh, C.-J. Optimum conditions for lipase immobilization on chitosan-coated Fe3O4 nanoparticles. Carbohydr. Polym. 2012, 87, 2538–2545. [Google Scholar] [CrossRef]
- Patel, S.K.; Anwar, M.Z.; Kumar, A.; Otari, S.V.; Pagolu, R.T.; Kim, S.Y.; Kim, I.W.; Lee, J.K. Fe2O3 yolk-shell particle-based laccase biosensor for efficient detection of 2,6-dimethoxyphenol. Biochem. Eng. J. 2018, 132, 1–8. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Chen, G.-J.; Twu, Y.-K.; Liu, Y.-C.; Shieh, C.-J. Optimum lipase immobilized on diamine-grafted PVDF membrane and its characterization. Ind. Eng. Chem. Res. 2012, 51, 5141–5147. [Google Scholar] [CrossRef]
- Singh, R.S.; Chauhan, K.; Kaur, N.; Kumar, N. Inulinase immobilization onto glutaraldehyde activated duolite XAD for the production of fructose from inulin. Biocatal. Agric. Biotechnol. 2020, 27, 101699. [Google Scholar] [CrossRef]
- Patel, S.K.; Choi, S.H.; Kang, Y.C.; Lee, J.K. Large-scale aerosol-assisted synthesis of biofriendly Fe2O3 yolk–shell particles: A promising support for enzyme immobilization. Nanoscale 2016, 8, 6728–6738. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.K.; Choi, H.; Lee, J.K. Multimetal-based inorganic–protein hybrid system for enzyme immobilization. ACS Sustain. Chem. Eng. 2019, 7, 13633–13638. [Google Scholar] [CrossRef]
- Nargotra, P.; Sharma, V.; Sharma, S.; Bangotra, R.; Bajaj, B.K. Purification of an ionic liquid stable cellulase from Aspergillus aculeatus PN14 with potential for biomass refining. Environ. Sustain. 2022, 5, 313–323. [Google Scholar] [CrossRef]
- Wong, H.-L.; Hu, N.-J.; Juang, T.-Y.; Liu, Y.-C. Co-immobilization of xylanase and scaffolding protein onto an immobilized metal ion affinity membrane. Catalysts 2020, 10, 1408. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Wong, J.H.; Ng, T.B. Immobilized metal ion affinity chromatography: A review on its applications. Appl. Microbiol. Biotechnol. 2012, 96, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Xi, C.R.; Di Fazio, A.; Nadvi, N.A.; Patel, K.; Xiang, M.S.W.; Zhang, H.E.; Deshpande, C.; Low, J.K.; Wang, X.T.; Chen, Y. A novel purification procedure for active recombinant human DPP4 and the inability of DPP4 to bind SARS-CoV-2. Molecules 2020, 25, 5392. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-Y.; Suen, S.-Y.; Chen, S.-C.; Tzeng, J.-H. Analysis of protein adsorption on regenerated cellulose-based immobilized copper ion affinity membranes. J. Chromatogr. A 2003, 996, 53–70. [Google Scholar] [CrossRef]
- Liu, Y.-C.; ChangChien, C.-C.; Suen, S.-Y. Purification of penicillin G acylase using immobilized metal affinity membranes. J. Chromatogr. B 2003, 794, 67–76. [Google Scholar] [CrossRef]
- Kim, D.; Ku, S. Bacillus cellulase molecular cloning, expression, and surface display on the outer membrane of Escherichia coli. Molecules 2018, 23, 503. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lu, Z.; Wang, X.; Selvaraj, J.N.; Zhang, G. Genetic engineering modification and fermentation optimization for extracellular production of recombinant proteins using Escherichia coli. Appl. Microbiol. Biotechnol. 2018, 102, 1545–1556. [Google Scholar] [CrossRef]
- Huleani, S.; Roberts, M.R.; Beales, L.; Papaioannou, E.H. Escherichia coli as an antibody expression host for the production of diagnostic proteins: Significance and expression. Crit. Rev. Biotechnol. 2022, 42, 756–773. [Google Scholar] [CrossRef]
- Mühlmann, M.; Forsten, E.; Noack, S.; Büchs, J. Optimizing recombinant protein expression via automated induction profiling in microtiter plates at different temperatures. Microb. Cell Factories 2017, 16, 1–12. [Google Scholar] [CrossRef]
- Lin, T.-N.; Lin, S.-C. Metal chelate-epoxy bifunctional membranes for selective adsorption and covalent immobilization of a His-tagged protein. J. Biosci. Bioeng. 2022, 133, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-J.; Kuo, C.-H.; Chen, C.-I.; Yu, C.-C.; Shieh, C.-J.; Liu, Y.-C. Effect of membranes with various hydrophobic/hydrophilic properties on lipase immobilized activity and stability. J. Biosci. Bioeng. 2012, 113, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Modenez, I.A.; Sastre, D.E.; Moraes, F.C.; Marques Netto, C.G. Influence of glutaraldehyde cross-linking modes on the recyclability of immobilized lipase B from Candida antarctica for transesterification of soy bean oil. Molecules 2018, 23, 2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, C.; Li, W.; Han, W.; Wang, Q.; Bi, P.; Han, C.; Zhu, L. Characterization of a novel thermostable GH7 endoglucanase from Chaetomium thermophilum capable of xylan hydrolysis. Int. J. Biol. Macromol. 2018, 117, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Chaga, G.S. Twenty-five years of immobilized metal ion affinity chromatography: Past, present and future. J. Biochem. Biophys. Methods 2001, 49, 313–334. [Google Scholar] [CrossRef]
- Li, T.; Gong, X.; Yang, G.; Li, Q.; Huang, J.; Zhou, N.; Jia, X. Cross-linked enzyme aggregates (CLEAs) of cellulase with improved catalytic activity, adaptability and reusability. Bioprocess Biosyst Eng. 2022, 45, 865–875. [Google Scholar] [CrossRef]
- Zeng, R.; Jin, B.K.; Yang, Z.H.; Guan, R.; Quan, C. Preparation of a modified crosslinked chitosan/polyvinyl alcohol blended affinity membrane for purification of his-tagged protein. J. Appl. Polym. Sci. 2019, 136, 47347. [Google Scholar] [CrossRef]
- Ahmad, R.; Khare, S.K. Immobilization of Aspergillus niger cellulase on multiwall carbon nanotubes for cellulose hydrolysis. Bioresour. Technol. 2018, 252, 72–75. [Google Scholar] [CrossRef]
- Kaur, P.; Taggar, M.S.; Kalia, A. Characterization of magnetic nanoparticle–immobilized cellulases for enzymatic saccharification of rice straw. Biomass Convers. Biorefin. 2021, 11, 955–969. [Google Scholar] [CrossRef]
- Saleem, M.; Rashid, M.; Jabbar, A.; Perveen, R.; Khalid, A.; Rajoka, M. Kinetic and thermodynamic properties of an immobilized endoglucanase from Arachniotus citrinus. Process Biochem. 2005, 40, 849–855. [Google Scholar] [CrossRef]
- Guo, R.; Zheng, X.; Wang, Y.; Yang, Y.; Ma, Y.; Zou, D.; Liu, Y. Optimization of cellulase immobilization with sodium alginate-polyethylene for enhancement of enzymatic hydrolysis of microcrystalline cellulose using response surface methodology. Appl. Biochem. Biotechnol. 2021, 193, 2043–2060. [Google Scholar] [CrossRef] [PubMed]
- Abbaszadeh, M.; Hejazi, P. Metal affinity immobilization of cellulase on Fe3O4 nanoparticles with copper as ligand for biocatalytic applications. Food Chem. 2019, 290, 47–55. [Google Scholar] [CrossRef]
- Saha, K.; Verma, P.; Sikder, J.; Chakraborty, S.; Curcio, S. Synthesis of chitosan-cellulase nanohybrid and immobilization on alginate beads for hydrolysis of ionic liquid pretreated sugarcane bagasse. Renew. Energy 2019, 133, 66–76. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, C.; Guo, R.; Ma, Y.; Yuan, Y.; Liu, Y. Cellulase immobilized by sodium alginate-polyethylene glycol-chitosan for hydrolysis enhancement of microcrystalline cellulose. Process Biochem. 2021, 107, 38–47. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, P.; Bajaj, B.K. Characterization of a robust serine protease from Bacillus subtilis K-1. J. Basic Microbiol. 2018, 58, 88–98. [Google Scholar] [CrossRef]
- Chien, H.-I.; Tsai, Y.-H.; Wang, H.-M.D.; Dong, C.-D.; Huang, C.-Y.; Kuo, C.-H. Extrusion puffing pretreated cereals for rapid production of high-maltose syrup. Food Chem. X 2022, 15, 100445. [Google Scholar] [CrossRef]
- Tüzmen, N.; Kalburcu, T.; Denizli, A. Immobilization of catalase via adsorption onto metal-chelated affinity cryogels. Process Biochem. 2012, 47, 26–33. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, Y.; Yang, T.; Zheng, D.; Liu, X.; Zhang, J.; Zheng, M. Preparation of immobilized Alcalase based on metal affinity for efficient production of bioactive peptides. LWT 2022, 162, 113505. [Google Scholar] [CrossRef]
- Chen, C.-I.; Ko, Y.-M.; Lien, W.-L.; Lin, Y.-H.; Li, I.-T.; Chen, C.-H.; Shieh, C.-J.; Liu, Y.-C. Development of the reversible PGA immobilization by using the immobilized metal ion affinity membrane. J. Membr. Sci. 2012, 401, 33–39. [Google Scholar] [CrossRef]
- Morrison, D. Transformation and preservation of competent bacterial cells by freezing. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1979; Volume 68, pp. 326–331. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Activity (U/mL or U/disc) | Protein (mg/mL) | Specific Activity (U/mg) | Purification Fold |
|---|---|---|---|---|
| Free CelA a | 1.250 | 1.322 | 0.946 | 1.00 |
| RC-EPI-IDA-Co2+-CelA b | 9.42 | 3.12 | 3.02 | 3.19 |
| RC-EPI-DA-GA-CelA b | 7.24 | 4.98 | 1.45 | 1.54 |
| Enzymes | Vmax (U/mg) | Km (mg/mL) |
|---|---|---|
| Free enzyme | 7.61 | 51.45 |
| RC-EPI-IDA-Co2+-CelA | 15.57 | 36.14 |
| RC-EPI-DA-GA-CelA | 10.85 | 37.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, Z.-H.; Nargotra, P.; Kuo, C.-H.; Liu, Y.-C. Immobilization of Recombinant Endoglucanase (CelA) from Clostridium thermocellum on Modified Regenerated Cellulose Membrane. Catalysts 2022, 12, 1356. https://doi.org/10.3390/catal12111356
Weng Z-H, Nargotra P, Kuo C-H, Liu Y-C. Immobilization of Recombinant Endoglucanase (CelA) from Clostridium thermocellum on Modified Regenerated Cellulose Membrane. Catalysts. 2022; 12(11):1356. https://doi.org/10.3390/catal12111356
Chicago/Turabian StyleWeng, Zi-Han, Parushi Nargotra, Chia-Hung Kuo, and Yung-Chuan Liu. 2022. "Immobilization of Recombinant Endoglucanase (CelA) from Clostridium thermocellum on Modified Regenerated Cellulose Membrane" Catalysts 12, no. 11: 1356. https://doi.org/10.3390/catal12111356