
Novel Transaminase and Laccase from Streptomyces spp. Using Combined Identification Approaches

 , , and
, , and
Abstract
:1. Introduction
2. Results
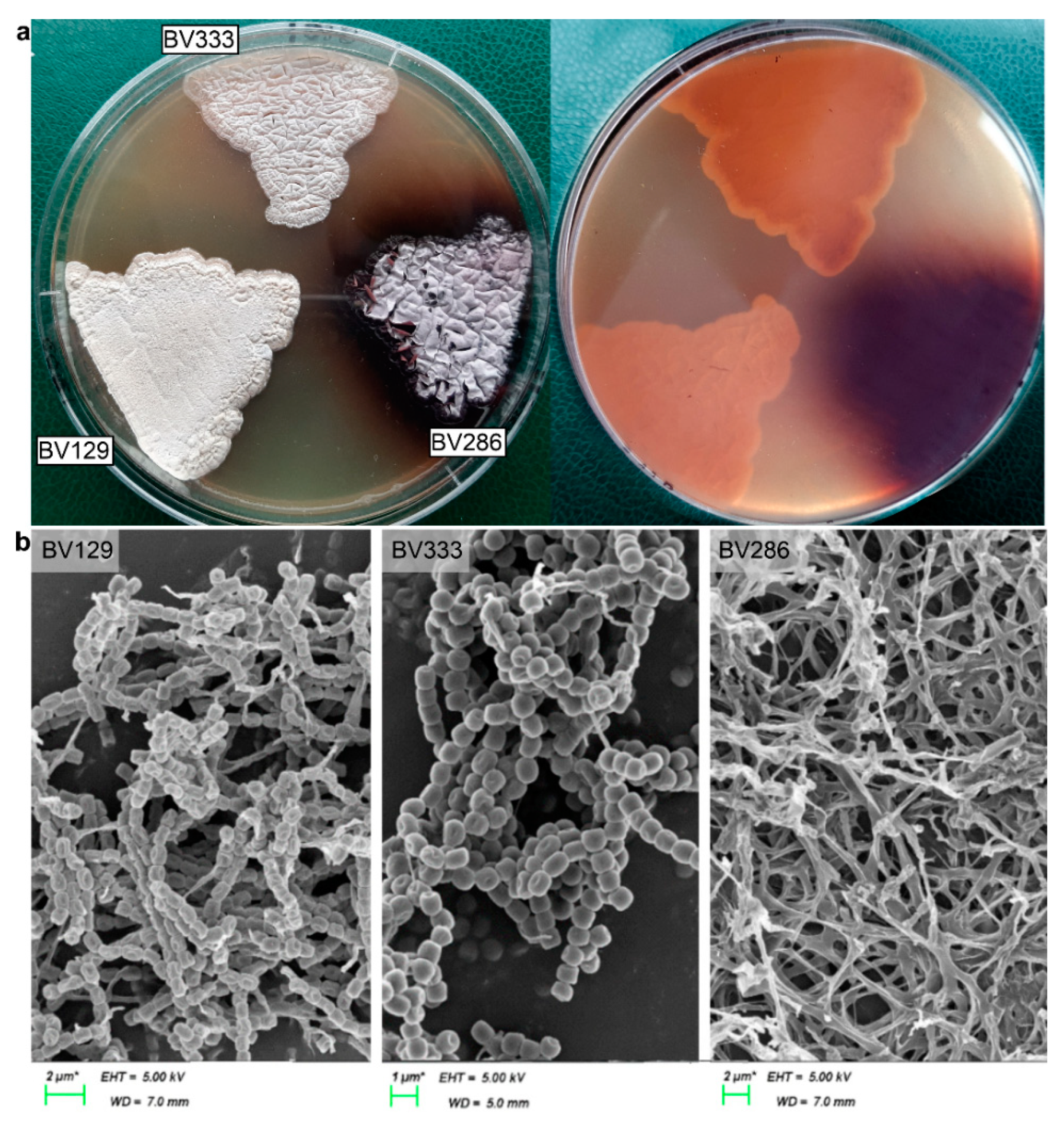
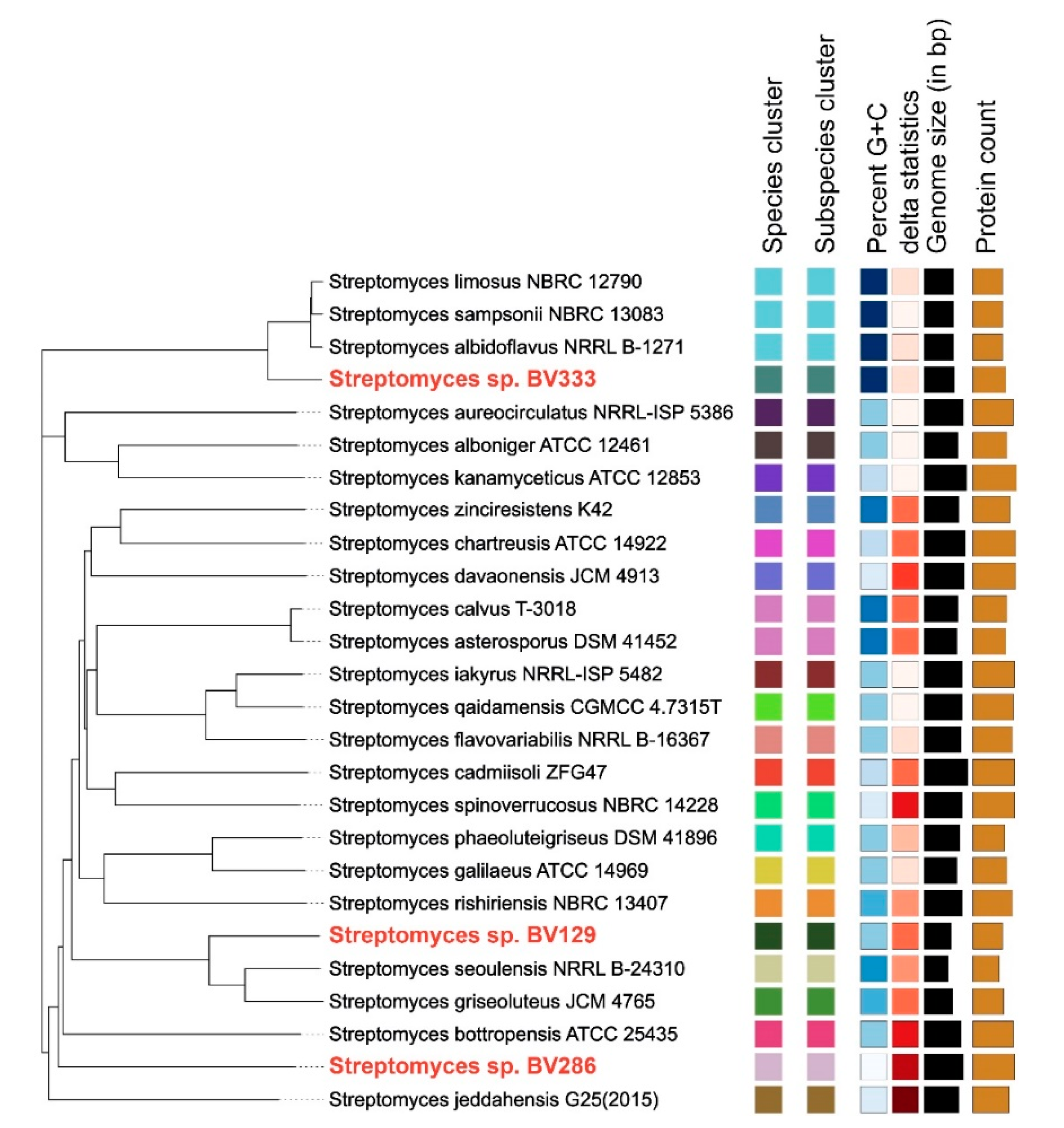
2.1. Genome Sequencing and Analysis
2.2. Bioinformatics Analysis of Transaminases and Laccases
2.3. Cloning, Expression and Functional Analysis of Novel Transaminase and Laccase
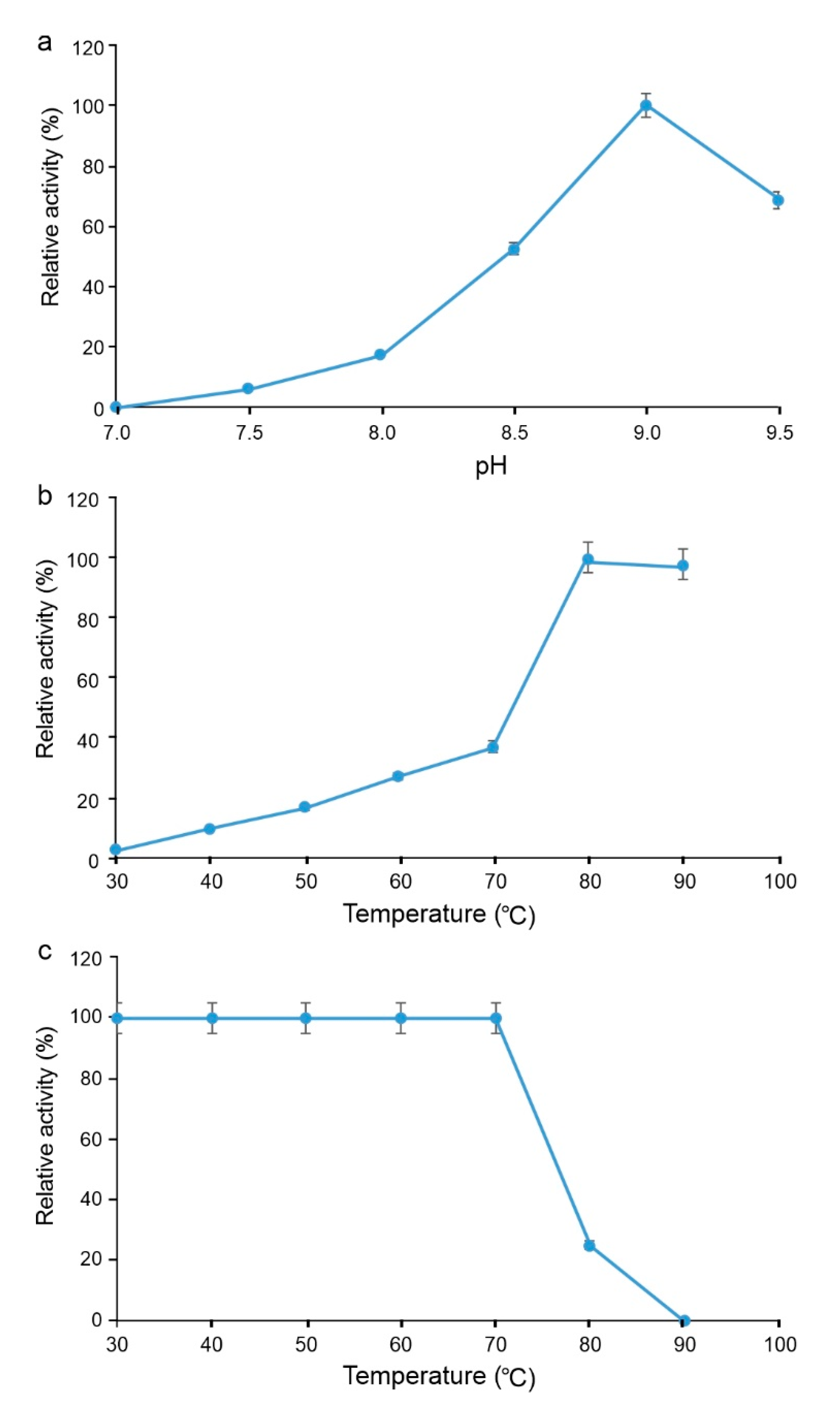
2.3.1. Functional Analysis of Sbv333-TA
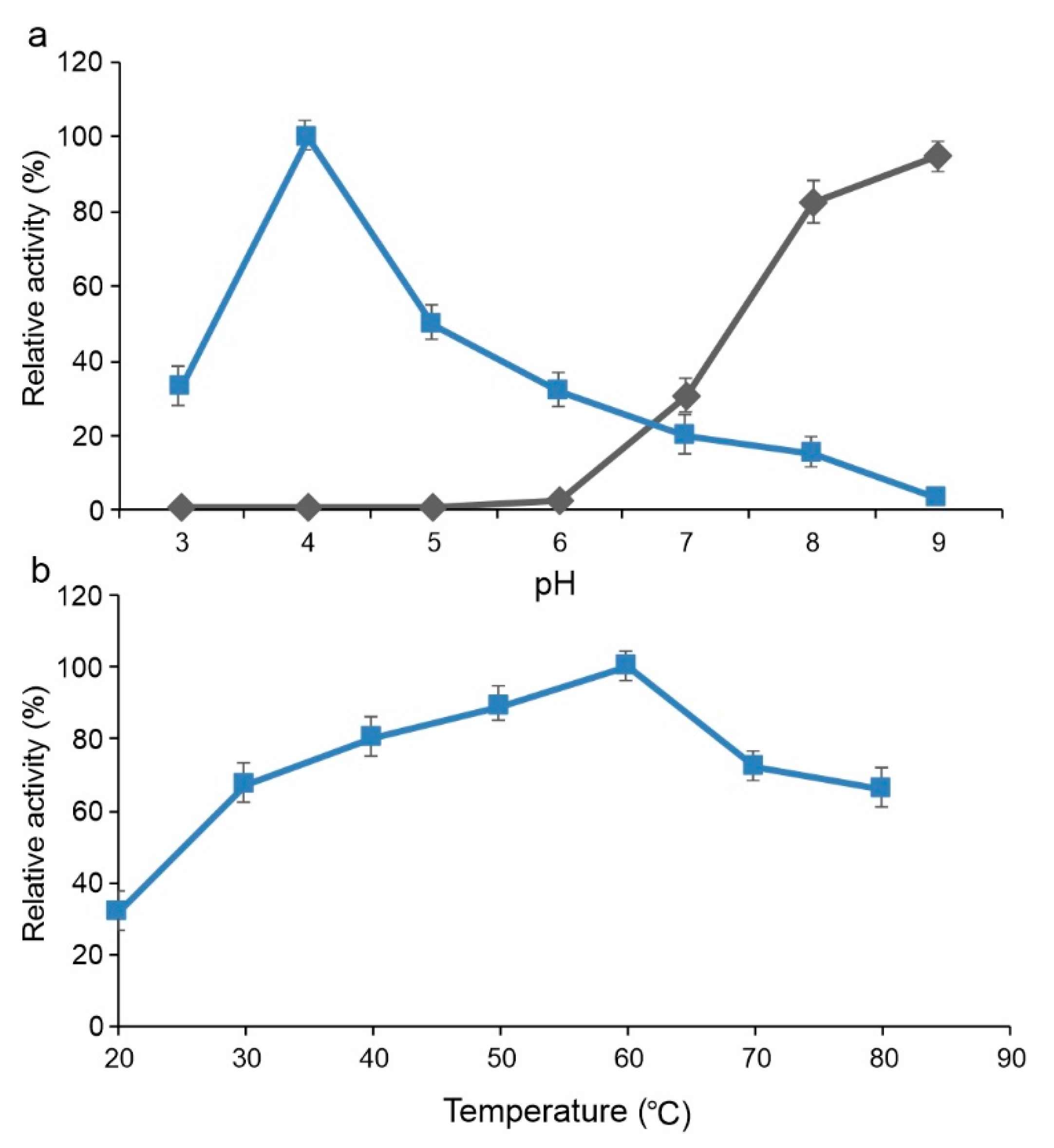
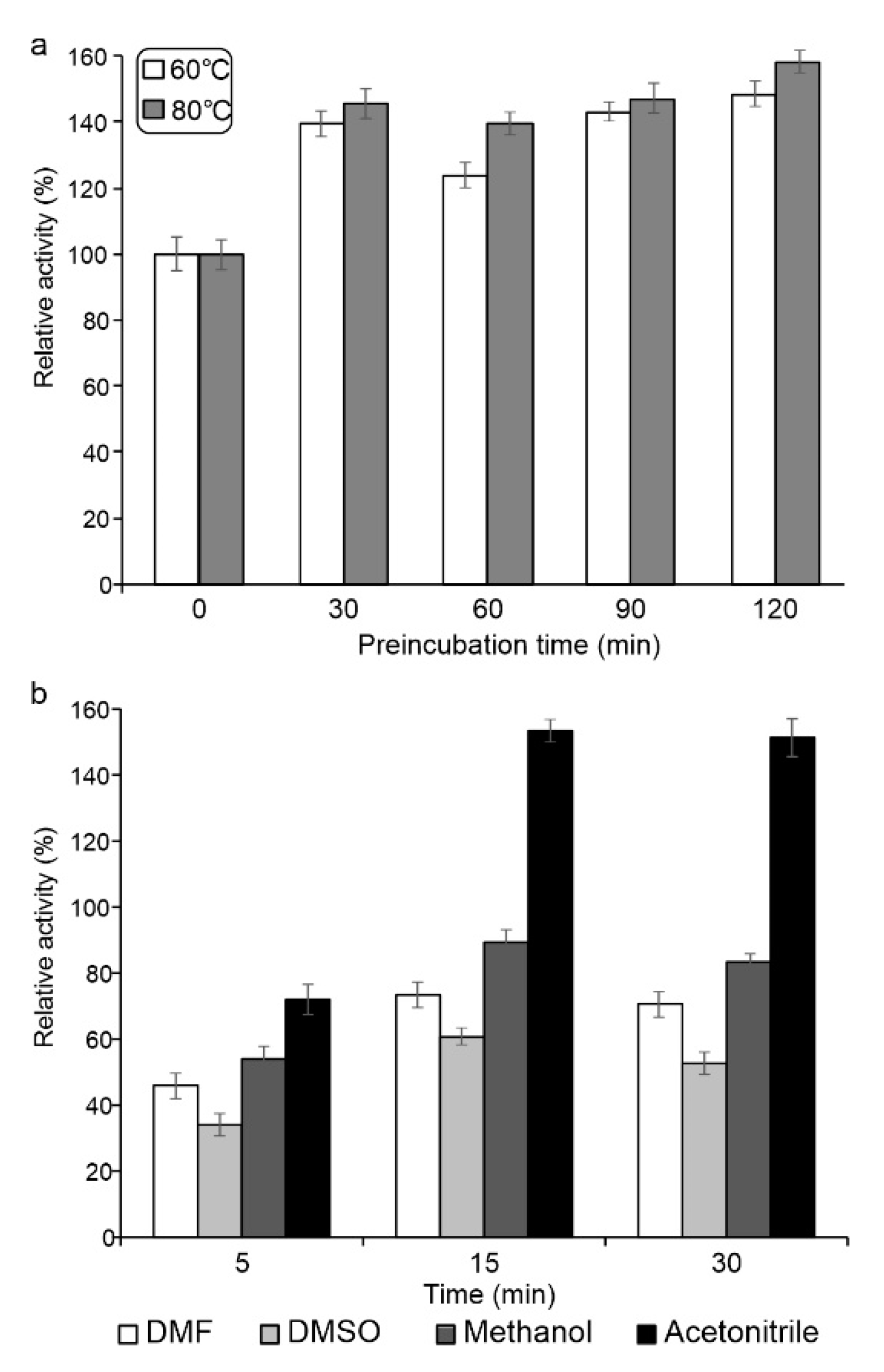
2.3.2. Functional Analysis of Sbv286-LAC
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Analytical Methods
4.3. Streptomyces sp. BV129, BV286, BV333 Genome Sequencing, Annotation, and Analysis
4.4. In Silico Screening for Novel Transaminases and Laccases
4.5. Gene Cloning and Recombinant Strains
4.5.1. Sbv333-TA Expression and Purification
4.5.2. Sbv286-LAC Expression and Purification
4.6. Functional Characterization of Sbv333-TA and Sbv286-LAC
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Spasic, J.; Mandic, M.; Djokic, L.; Nikodinovic-Runic, J. Streptomyces spp. in the biocatalysis toolbox. Appl. Microbiol. Biotechnol. 2018, 102, 3513–3536. [Google Scholar] [CrossRef]
- Spasic, J.; Mandic, M.; Radivojevic, J.; Jeremic, S.; Vasiljevic, B.; Nikodinovic-Runic, J.; Djokic, L. Biocatalytic potential of Streptomyces spp. isolates from rhizosphere of plants and mycorrhizosphere of fungi. Biotechnol. Appl. Biochem. 2018, 65, 822–833. [Google Scholar] [CrossRef]
- Lopes, A.M.; Ferreira Filho, E.X.; Moreira, L.R.S. An update on enzymatic cocktails for lignocellulose breakdown. J. Appl. Microbiol. 2018, 125, 632–645. [Google Scholar] [CrossRef] [Green Version]
- Pellis, A.; Cantone, S.; Ebert, C.; Gardossi, L. Evolving biocatalysis to meet bioeconomy challenges and opportunities. New Biotechnol. 2018, 40, 154–169. [Google Scholar] [CrossRef]
- Markets&Markets. Biocatalysis & Biocatalysts Market by Type, Application, and Geography-Forecast to 2019. 2015. Available online: https://www.marketsandmarkets.com/Market-Reports/biocatalysis-biocatalyst-market-71612080.html (accessed on 12 May 2021).
- Kelly, S.A.; Magill, D.J.; Megaw, J.; Skvortsov, T.; Allers, T.; McGrath, J.W.; Allen, C.C.R.; Moody, T.S.; Gilmore, B.F. Characterisation of a solvent-tolerant haloarchaeal (R)-selective transaminase isolated from a Triassic period salt mine. Appl. Microbiol. Biotechnol. 2019, 103, 5727–5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Li, Y.; Tian, Z.; Qian, Y.; Zhang, H.; Wang, L. A novel thermostable chitinolytic machinery of Streptomyces sp. F-3 consisting of chitinases with different action modes. Biotechnol. Biofuels 2019, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Prakash, D.; Nawani, N.; Prakash, M.; Bodas, M.; Mandal, A.; Khetmalas, M.; Kapadnis, B. Actinomycetes: A repertory of green catalysts with a potential revenue resource. BioMed Res. Int. 2013, 2013, 264020. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.S.; Park, D.J.; Simkhada, J.R.; Hong, J.H.; Sohng, J.K.; Lee, O.H.; Yoo, J.C. A neutral lipase applicable in biodiesel production from a newly isolated Streptomyces sp. CS326. Bioprocess Biosyst. Eng. 2012, 35, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Ferrandi, E.E.; Monti, D. Amine transaminases in chiral amines synthesis: Recent advances and challenges. World J. Microbiol. Biotechnol. 2017, 34, 13. [Google Scholar] [CrossRef]
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleitz, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Yonaha, K.; Suzuki, K.; Toyama, S. Streptomyces beta-alanine: Alpha-ketoglutarate aminotransferase, a novel omega-amino acid transaminase. Purification, crystallization, and enzymologic properties. J. Biol. Chem. 1985, 260, 3265–3268. [Google Scholar] [CrossRef]
- Yonaha, K.; Suzuki, K.; Toyama, S. 4-Aminobutyrate:2-oxoglutarate aminotransferase of Streptomyces griseus: Purification and properties. Eur. J. Biochem. 1985, 146, 101–106. [Google Scholar] [CrossRef]
- Burgie, E.S.; Thoden, J.B.; Holden, H.M. Molecular architecture of DesV from Streptomyces venezuelae: A PLP-dependent transaminase involved in the biosynthesis of the unusual sugar desosamine. Protein Sci. 2007, 16, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassanini, I.; Ferrandi, E.E.; Riva, S.; Monti, D. Biocatalysis with laccases: An updated overview. Catalysts 2021, 11, 26. [Google Scholar] [CrossRef]
- Zerva, A.; Simić, S.; Topakas, E.; Nikodinovic-Runic, J. Applications of microbial laccases: Patent review of the past decade (2009–2019). Catalysts 2019, 9, 1023. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.-B.; Luo, Q.; Wang, H.-R.; Chen, Y.; Liao, X.-R. Bacterial laccases: Promising biological green tools for industrial applications. Cell. Molec. Life Sci. 2018, 75, 3569–3592. [Google Scholar] [CrossRef]
- Ece, S.; Lambertz, C.; Fischer, R.; Commandeur, U. Heterologous expression of a Streptomyces cyaneus laccase for biomass modification applications. AMB Express 2017, 7, 86. [Google Scholar] [CrossRef]
- Sherif, M.; Waung, D.; Korbeci, B.; Mavisakalyan, V.; Flick, R.; Brown, G.; Abou-Zaid, M.; Yakunin, A.F.; Master, E.R. Biochemical studies of the multicopper oxidase (small laccase) from Streptomyces coelicolor using bioactive phytochemicals and site-directed mutagenesis. Microbial Biotechnol. 2013, 6, 588–597. [Google Scholar] [CrossRef]
- Gunne, M.; Urlacher, V.B. Characterization of the alkaline laccase Ssl1 from Streptomyces sviceus with unusual properties discovered by genome mining. PLoS ONE 2012, 7, e52360. [Google Scholar] [CrossRef] [PubMed]
- Gunne, M.; Höppner, A.; Hagedoorn, P.L.; Urlacher, V.B. Structural and redox properties of the small laccase Ssl1 from Streptomyces sviceus. FEBS J. 2014, 281, 4307–4318. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Comm. 2019, 10, 2182. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Steffen-Munsberg, F.; Vickers, C.; Kohls, H.; Land, H.; Mallin, H.; Nobili, A.; Skalden, L.; van den Bergh, T.; Joosten, H.J.; Berglund, P.; et al. Bioinformatic analysis of a PLP-dependent enzyme superfamily suitable for biocatalytic applications. Biotechnol. Advan. 2015, 33, 566–604. [Google Scholar] [CrossRef] [PubMed]
- Mathew, S.; Deepankumar, K.; Shin, G.; Hong, E.Y.; Kim, B.-G.; Chung, T.; Yun, H. Identification of novel thermostable ω-transaminase and its application for enzymatic synthesis of chiral amines at high temperature. RSC Adv. 2016, 6, 69257–69260. [Google Scholar] [CrossRef]
- Ferrandi, E.E.; Previdi, A.; Bassanini, I.; Riva, S.; Peng, X.; Monti, D. Novel thermostable amine transferases from hot spring metagenomes. Appl. Microbiol. Biotechnol. 2017, 101, 4963–4979. [Google Scholar] [CrossRef] [PubMed]
- Höhne, M.; Schätzle, S.; Jochens, H.; Robins, K.; Bornscheuer, U.T. Rational assignment of key motifs for function guides in silico enzyme identification. Nat. Chem. Biol. 2010, 6, 807. [Google Scholar] [CrossRef] [PubMed]
- Sayer, C.; Martinez-Torres, R.J.; Richter, N.; Isupov, M.N.; Hailes, H.C.; Littlechild, J.A.; Ward, J.M. The substrate specificity, enantioselectivity and structure of the (R)-selective amine: Pyruvate transaminase from Nectria haematococca. FEBS J. 2014, 281, 2240–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihssen, J.; Reiss, R.; Luchsinger, R.; Thöny-Meyer, L.; Richter, M. Biochemical properties and yields of diverse bacterial laccase-like multicopper oxidases expressed in Escherichia coli. Sci. Rep. 2015, 5, 10465. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.-B.; Song, C.-M.; Zhang, N.; Zhou, W.; Xu, C.-W.; Zhou, L.-X.; Zhao, H.; Cai, Y.-J.; Liao, X.-R. Overexpression, characterization, and dye-decolorizing ability of a thermostable, pH-stable, and organic solvent-tolerant laccase from Bacillus pumilus W3. J. Mol. Catal. B Enzym. 2014, 101, 1–6. [Google Scholar] [CrossRef]
- Tonin, F.; Melis, R.; Cordes, A.; Sanchez-Amat, A.; Pollegioni, L.; Rosini, E. Comparison of different microbial laccases as tools for industrial uses. New Biotechnol. 2016, 33, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-H.; Lin, M.-C.; Lee, C.-C.; Yu, S.-M.; Wang, A.H.J.; Ho, T.-H.D. Enhancement of laccase activity by pre-incubation with organic solvents. Sci. Rep. 2019, 9, 9754. [Google Scholar] [CrossRef] [PubMed]
- Buß, O.; Voss, M.; Delavault, A.; Gorenflo, P.; Syldatk, C.; Bornscheuer, U.; Rudat, J. β-Phenylalanine ester synthesis from stable β-keto ester substrate using engineered ω-transaminases. Molecules 2018, 23, 1211. [Google Scholar] [CrossRef] [Green Version]
- Midelfort, K.S.; Kumar, R.; Han, S.; Karmilowicz, M.J.; McConnell, K.; Gehlhaar, D.K.; Mistry, A.; Chang, J.S.; Anderson, M.; Villalobos, A.; et al. Redesigning and characterizing the substrate specificity and activity of Vibrio fluvialis aminotransferase for the synthesis of imagabalin. Protein Eng. Des. Sel. 2012, 26, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Nikodinovic, J.; Barrow, K.D.; Chuck, J.A. High yield preparation of genomic DNA from Streptomyces. BioTechniques 2003, 35, 932–934. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [Green Version]
- Farris, J.S. Estimating phylogenetic trees from distance matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]
- Kielbasa, S.M.; Wan, R.; Sato, K.; Horton, P.; Frith, M.C. Adaptive seeds tame genomic sequence comparison. Genome Res. 2011, 21, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Streptomyces sp. BV129 | Streptomyces sp. BV286 | Streptomyces sp. BV333 |
|---|---|---|---|
| Genome size | 6,475,242 | 9,426,047 | 7,323,588 |
| Genome coverage | 83.50 | 59.02 | 72.73 |
| Number of contigs | 82 | 212 | 699 |
| Longest contig | 704,247 | 515,226 | 151,113 |
| N50 * | 144,188 | 114,510 | 28,611 |
| L50 ** | 15 | 26 | 80 |
| GC content (%) | 71.86 | 70.77 | 73.2 |
| Predicted genes | 5933 | 8326 | 6373 |
| Predicted tRNAs | 84 | 92 | 83 |
| Predicted CDS | 5848 | 8233 | 6289 |
| Predicted tmRNA | 1 | 1 | 1 |
| EC no. | Streptomyces sp. BV129 | Streptomyces sp. BV286 | Streptomyces sp. BV333 | |
|---|---|---|---|---|
| lipase | 3.1.1.3 | 7 | 8 | 4 |
| cutinase | 3.1.1.74 | 0 | 1 | 0 |
| cellulase | 3.2.1.4 | 3 | 11 | 3 |
| protease | 3.4.-.- | 124 | 160 | 136 |
| laccase | 1.10.3.1 | 2 | 1 | 1 |
| tyrosinase | 1.14.18.1 | 1 | 1 | 0 |
| aminotransferase (transaminase) | 2.6.1.- | 32 | 35 | 38 |
| Predicted genes with EC number | 2049 | 2763 | 2114 |
| Amino Acceptor | Conversion (%) | |
|---|---|---|
 | α-ketoglutarate | - |
 | Pyruvate | 54 |
 | 2-Oxobutyrate | 46 |
 | Glioxylate | 88 |
 | Methylacetoacetate | 52 |
 | Ethylacetoacetate | 35 |
 | Ethylbenzoylacetate | 47 |
 | Methyl isobutyl ketone | 7 |
 | Propionaldehyde | 100 |
 | Phenylpropionaldehyde | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrandi, E.E.; Spasic, J.; Djokic, L.; Vainshtein, Y.; Senthamaraikannan, R.; Vojnovic, S.; Grumaz, C.; Monti, D.; Nikodinovic-Runic, J. Novel Transaminase and Laccase from Streptomyces spp. Using Combined Identification Approaches. Catalysts 2021, 11, 919. https://doi.org/10.3390/catal11080919
Ferrandi EE, Spasic J, Djokic L, Vainshtein Y, Senthamaraikannan R, Vojnovic S, Grumaz C, Monti D, Nikodinovic-Runic J. Novel Transaminase and Laccase from Streptomyces spp. Using Combined Identification Approaches. Catalysts. 2021; 11(8):919. https://doi.org/10.3390/catal11080919
Chicago/Turabian StyleFerrandi, Erica E., Jelena Spasic, Lidija Djokic, Yevhen Vainshtein, Ramsankar Senthamaraikannan, Sandra Vojnovic, Christian Grumaz, Daniela Monti, and Jasmina Nikodinovic-Runic. 2021. "Novel Transaminase and Laccase from Streptomyces spp. Using Combined Identification Approaches" Catalysts 11, no. 8: 919. https://doi.org/10.3390/catal11080919