
Synthesis and Laccase-Mediated Oxidation of New Condensed 1,4-Dihydropyridine Derivatives

 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
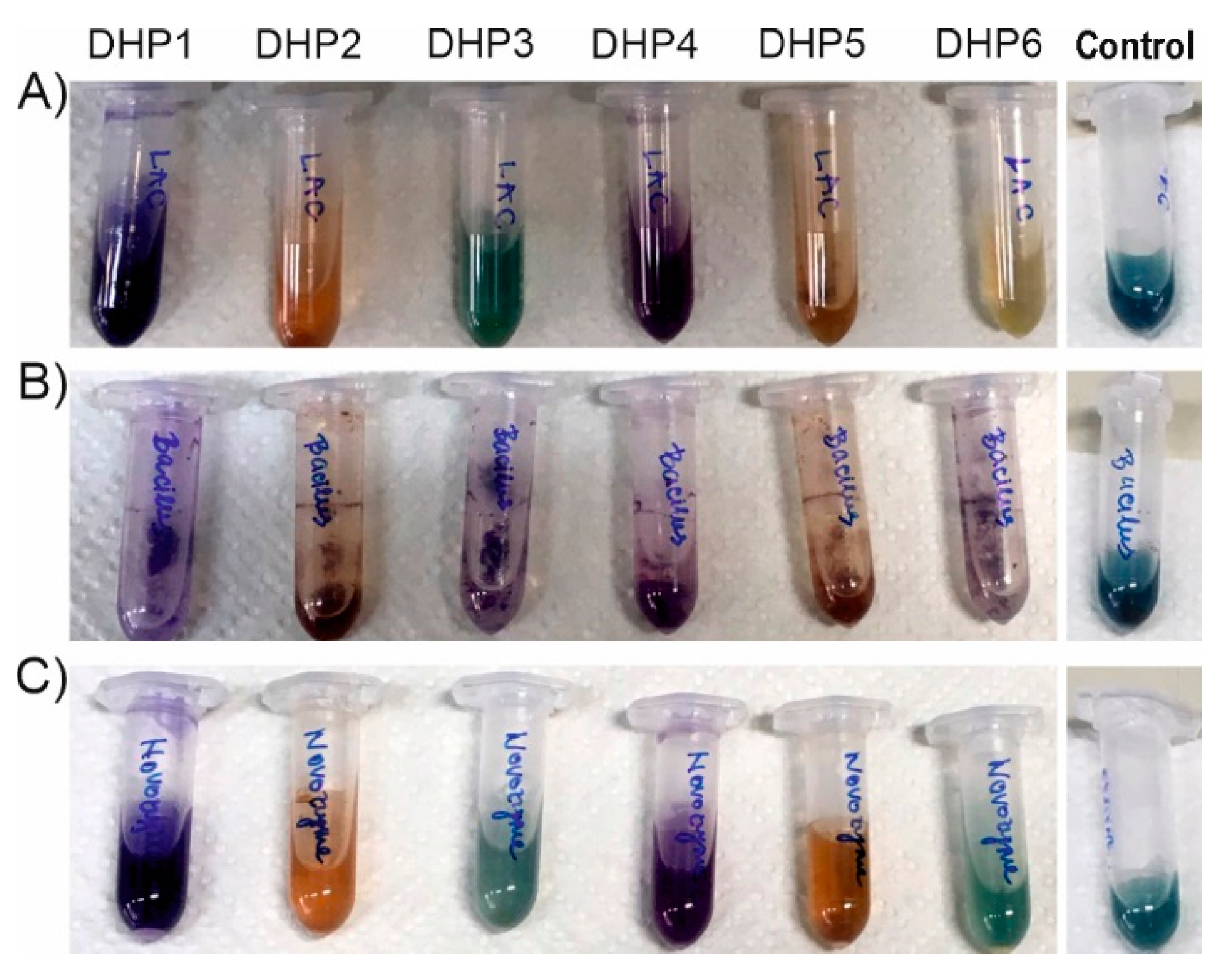
2.1. Oxidation of DHPs by Laccases from Trametes versicolor (TvLacc)
2.2. Oxidation of DHPs by Laccases with Different Redox Potential
3. Materials and Methods
3.1. Reagents and Chemicals
3.2. Analytical Methods
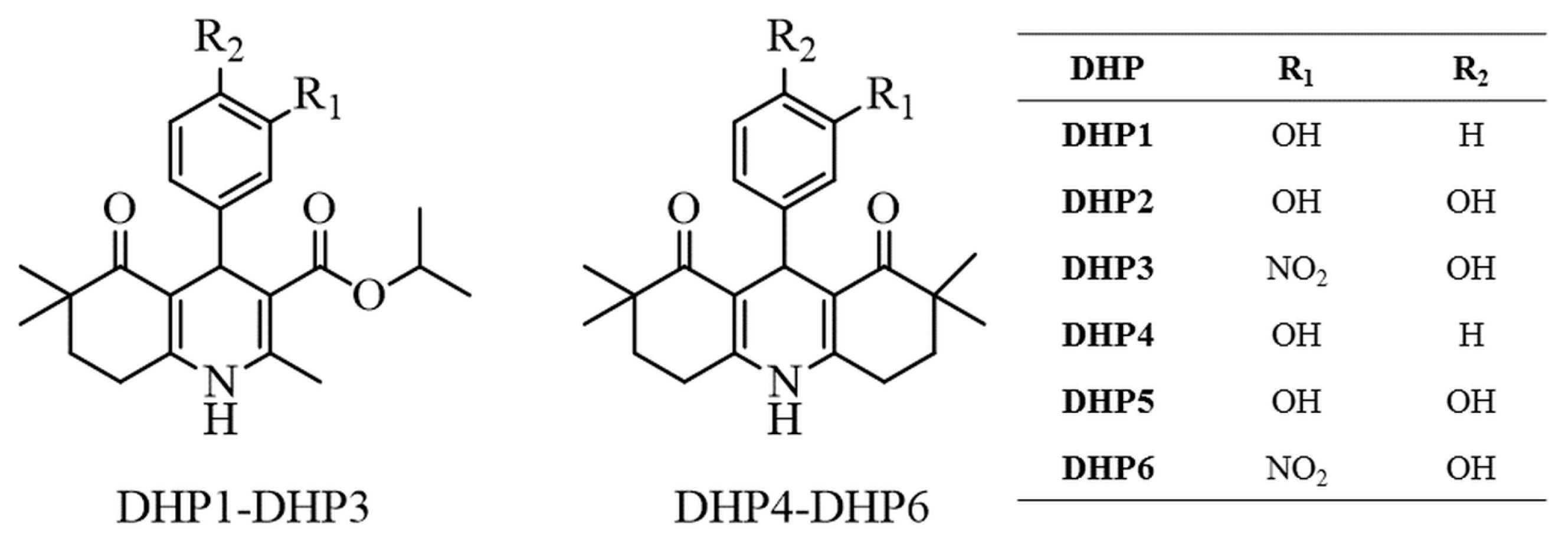
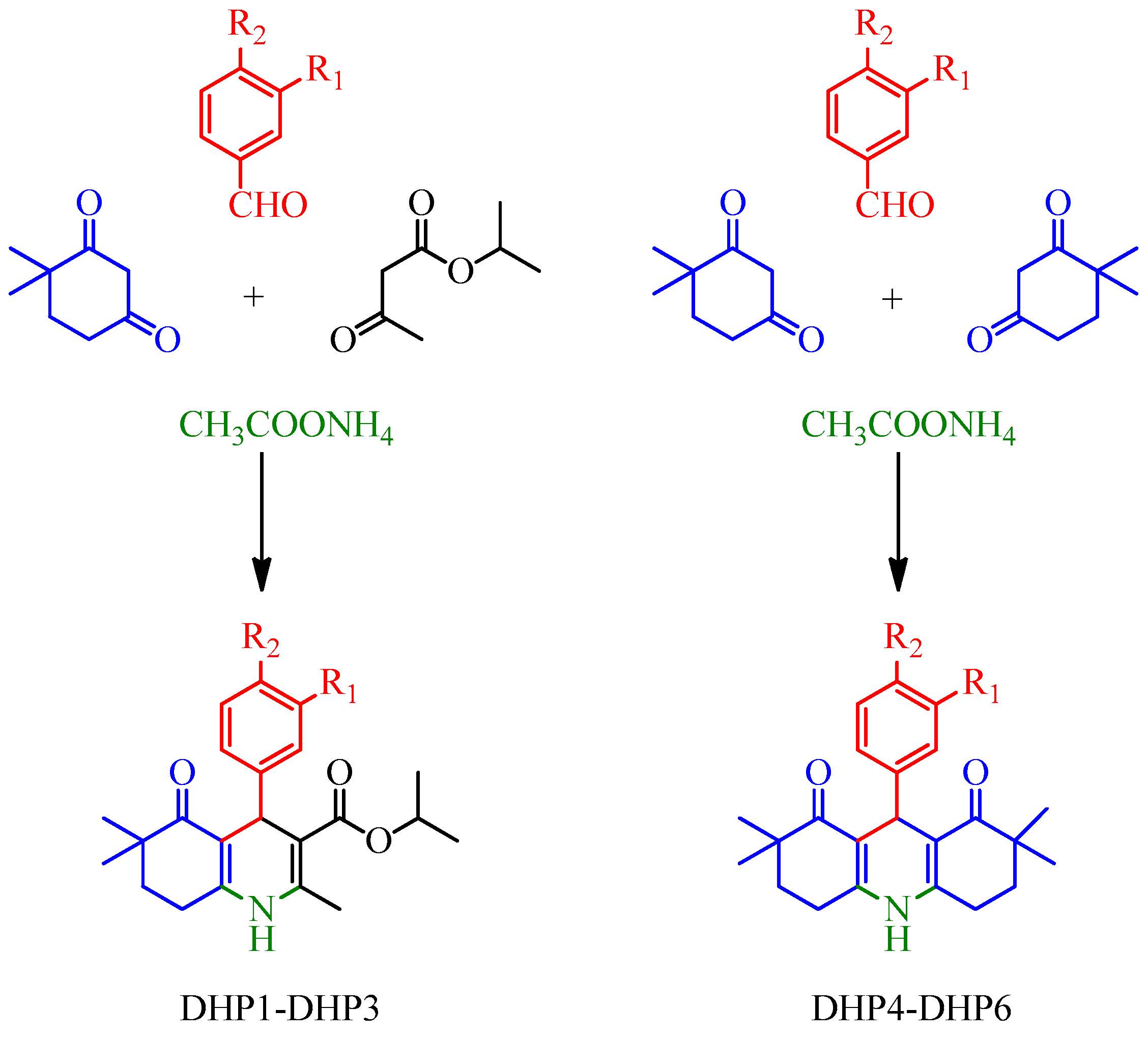
3.3. General Procedure for the Synthesis and Structure Elucidation of DHPs
- Isopropyl 4-(3-hydroxyphenyl)-2,6,6-trimethyl-5-oxo-1,4,5,6,7,8-hexahydroquinoline-3-carboxylate (DHP1): Yield: 34%. M.p. 229–231 °C. 1H-NMR (δ, DMSO-d6): 0.88 (3H; s), 0.95 (3H; s), 1.05 (3H; d; J = 6.4 Hz), 1.15 (3H; d; J = 6.4 Hz), 1.67–1.71 (2H; m), 2.22 (3H; s), 2.44–2.48 (2H; m), 4.74 (1H; s), 4.77–4.83 (1H; m), 6.41–6.93 (4H; m), 8.97 (1H; s), 9.02 (1H; s). ESI-MS (m/z): 392 [M+Na]+. Anal. Calcd. for C22H27NO4; C, 71.52; H, 7.37; N, 3.79. Found: C, 71.25; H, 7.44; N, 3.82.
- Isopropyl 4-(3,4-dihydroxyphenyl)-2,6,6-trimethyl-5-oxo-1,4,5,6,7,8-hexahydro quinoline-3-carboxylate (DHP2): Yield: 32%. M.p. 242–244 °C. 1H-NMR (δ, DMSO-d6): 0.90 (3H; s), 0.97 (3H; s), 1.08 (3H; d; J = 6.4 Hz), 1.17 (3H; d; J = 6.4 Hz), 1.69–1.73 (2H; m), 2.22 (3H; s), 2.45–2.49 (2H; m), 4.66 (1H; s), 4.79–4.85 (1H; m), 6.38 (1H; dd; J = 2.0/8.0 Hz), 6.49 (1H; d; J = 8.0 Hz), 6.54 (1H; d; J = 2.0 Hz), 8.45 (1H; s), 8.53 (1H; s), 8.92 (1H; s). ESI-MS (m/z): 408 [M+Na]+. Anal. Calcd. for C22H27NO5; C, 68.55; H, 7.06; N, 3.63. Found: C, 68.38; H, 7.11; N, 3.62.
- Isopropyl 4-(4-hydroxy-3-nitrophenyl)-2,6,6-trimethyl-5-oxo-1,4,5,6,7,8-hexahydro quinoline-3-carboxylate (DHP3): Yield: 65%. M.p. 256–258 °C. 1H-NMR (δ, DMSO-d6): 0.86 (3H; s), 0.95 (3H; s), 1.02 (3H; d; J = 6.4 Hz), 1.16 (3H; d; J= 6.4 Hz), 1.66–1.72 (2H; m), 2.24 (3H; s), 2.45–2.49 (2H; m), 4.75 (1H; s), 4.77–4.83 (1H; m), 6.95 (1H; d; J = 8.8 Hz), 7.28 (1H; dd; J = 2.0/8.8 Hz), 7.57 (1H; d; J= 2.0 Hz), 9.09 (1H; s). ESI-MS (m/z): 437 [M+Na]+. Anal. Calcd. for C22H26N2O6; C, 63.76; H, 6.32; N, 6.76. Found: C, 63.31; H, 6.29; N, 6.76.
- 9-(3-hydroxyphenyl)-2,2,7,7-tetramethyl-3,4,6,7,9,10-hexahydroacridine-1,8(2H,5H) -dione (DHP4): Yield: 25%. M.p. 294–295 °C. 1H-NMR (δ, DMSO-d6): 0.91 (6H; s), 0.97 (6H; s), 1.71–1.75 (4H; m), 2.49–2.53 (4H; m), 4.79 (1H; s), 6.37–6.90 (4H; m), 9.01 (1H; s), 9.29 (1H; s). ESI-MS (m/z): 388 [M+Na]+. Anal. Calcd. for C23H27NO3; C, 75.59; H, 7.45; N, 3.83. Found: C, 75.43; H, 7.39; N, 3.80.
- 9-(3,4-dihydroxyphenyl)-2,2,7,7-tetramethyl-3,4,6,7,9,10-hexahydroacridine-1,8 (2H,5H)-dione (DHP5): Yield: 39%. M.p. 260–261 °C. 1H-NMR (δ, DMSO-d6): 0.89 (6H; s), 0.94 (6H; s), 1.70 (4H; t), 2.47 (4H; t), 4.68 (1H; s), 6.32 (1H; dd; J = 2.0/8.0 Hz), 6.44 (1H; d; J = 8.0 Hz), 6.52 (1H; d; J = 2.0 Hz), 8.34 (1H; s), 8.50 (1H; s), 9.19 (1H; s). ESI-MS (m/z): 404 [M+Na]+. Anal. Calcd. for C23H27NO4; C, 72.42; H, 7.13; N, 3.67. Found: C, 72.20; H, 7.07; N, 3.69.
- 9-(4-hydroxy-3-nitrophenyl)-2,2,7,7-tetramethyl-3,4,6,7,9,10-hexahydroacridine-1,8 (2H,5H)-dione (DHP6): Yield: 58%. M.p. 267–268 °C. 1H-NMR (δ, DMSO-d6): 0.87 (6H; s), 0.95 (6H; s), 1.69-1.73 (4H; m), 2.47–2.52 (4H; m), 4.77 (1H; s), 6.92 (1H; d; J = 8.4 Hz), 7.30 (1H; dd; J = 2.4/8.4 Hz), 7.52 (1H; d; J = 2.4 Hz), 9.38 (1H; s). ESI-MS (m/z): 433 [M+Na]+. Anal. Calcd. for C23H26N2O5; C, 67.30; H, 6.38; N, 6.82. Found: C, 67.43; H, 6.43; N, 6.89.
3.4. Biocatalytic Reaction Setup and Analytical Procedures
3.4.1. General Procedure for the Laccase-Catalyzed Oxidation of DHPs -Analytical Scale
3.4.2. General Procedure for the Laccase-Catalyzed Oxidation of DHPs -Medium Scale
3.4.3. HPLC Analysis of Reaction Mixtures
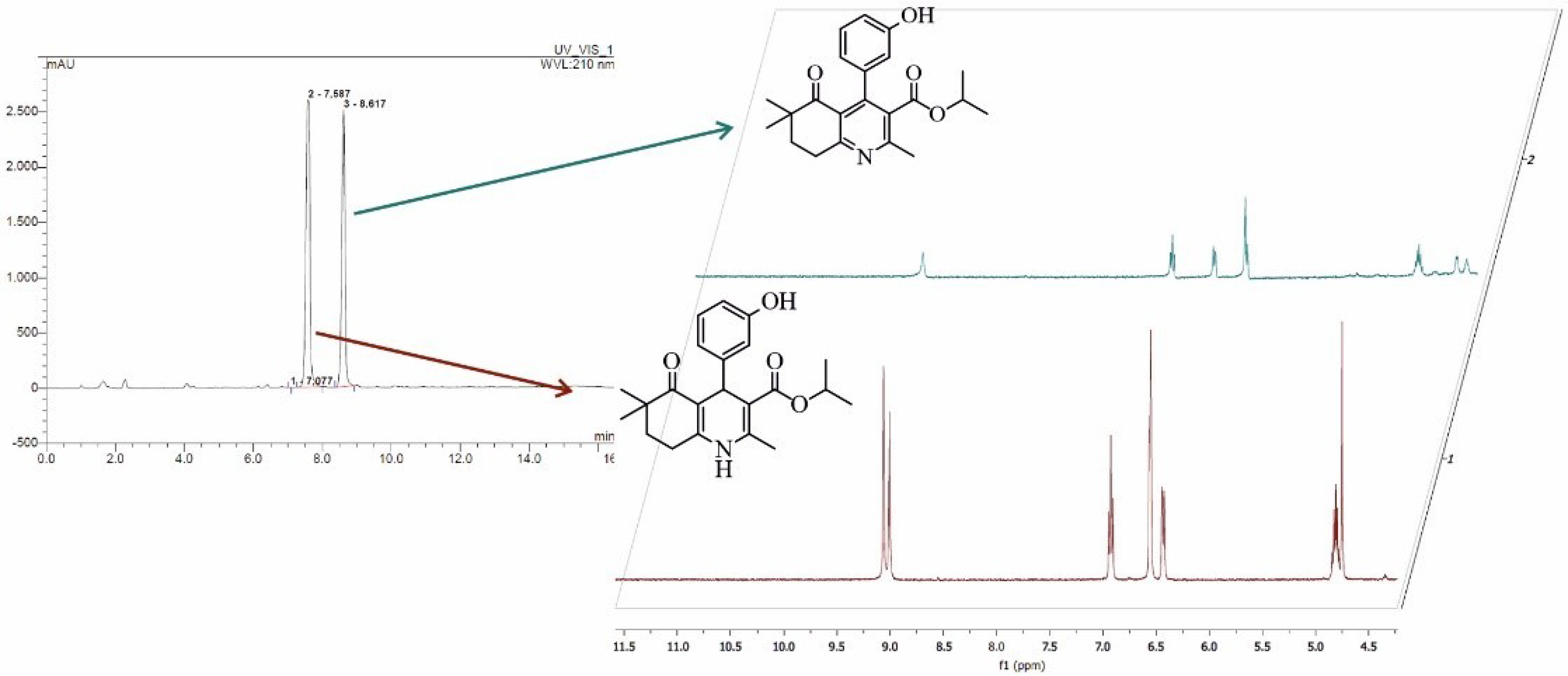
3.4.4. NMR Analysis of Isolated Product from Biocatalytic Oxidation of DHP1
- Isopropyl 4-(3-hydroxyphenyl)-2,6,6-trimethyl-5-oxo-5,6,7,8-tetrahydroquinoline-3-carboxylate: 1H-NMR (δ, DMSO-d6): 9.45 (s, 1H), 7.10 (t, J = 7.7 Hz, 1H), 6.71 (d, J = 7.3 Hz, 1H), 6.40 (d, J = 7.9 Hz, 1H), 4.79 (dd, J = 12.3, 6.1 Hz, 1H), 3.10 (t, J = 6.2 Hz, 2H), 2.45 (s, 3H), 1.94 (t, J = 6.4 Hz, 2H), 1.06 (s, 3H), 0.94 (d, J = 6.2 Hz, 3H). 13C NMR (δ, DMSO-d6): 201.74, 166.81, 163.39, 157.16, 156.86, 148.45, 138.91, 129.94, 129.17, 123.73, 118.69, 115.02, 114.81, 69.19, 42.58, 34.59, 29.51, 24.09, 22.96, 21.38.
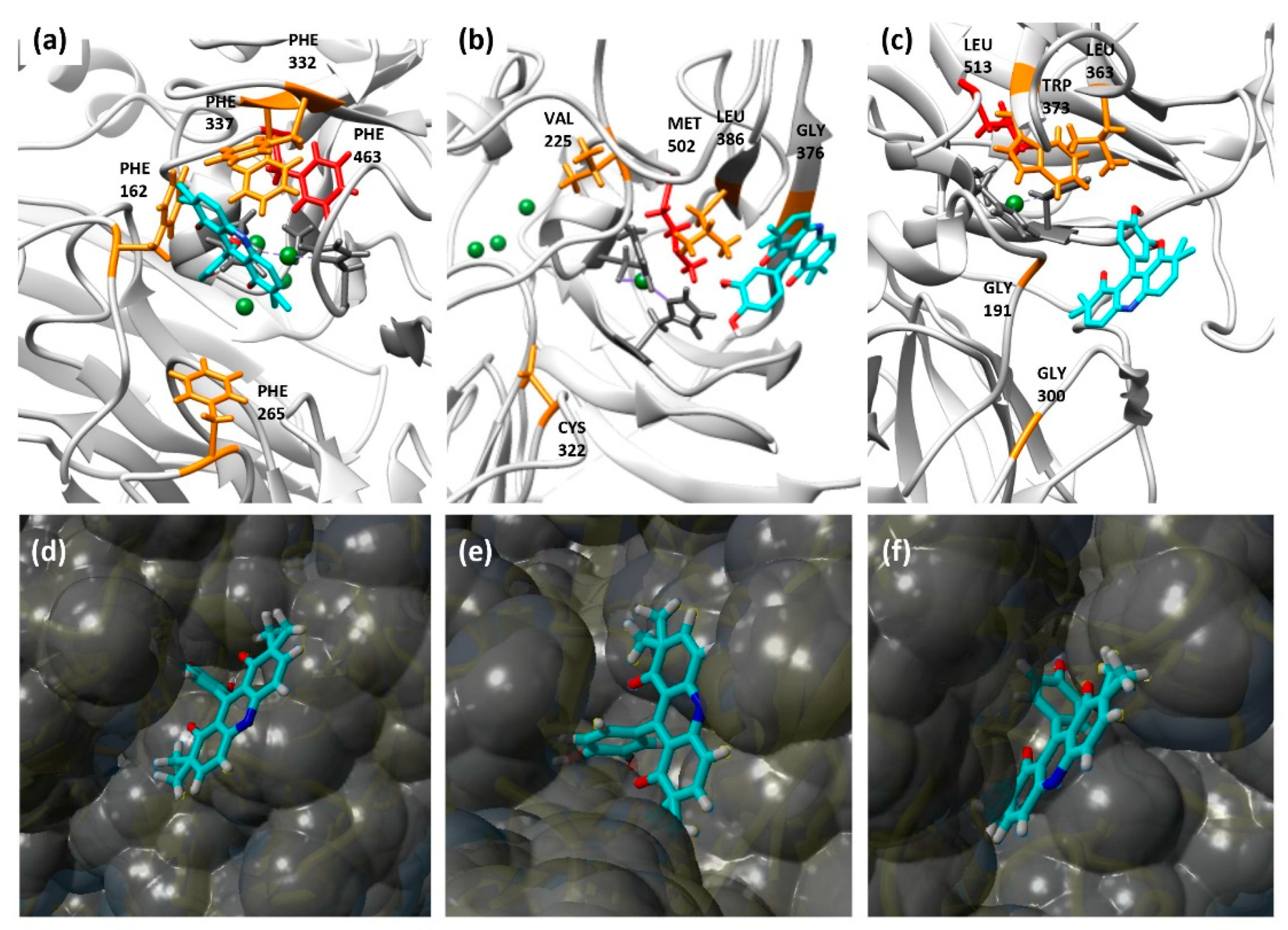
3.5. Molecular Docking Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mishra, A.P.; Bajpai, A.; Rai, A.K. 1,4-Dihydropyridine: A Dependable Heterocyclic Ring with the Promising and the Most Anticipable Therapeutic Effects. Mini Rev. Med. Chem. 2019, 19, 1219–1254. [Google Scholar] [CrossRef] [PubMed]
- Aygün Cevher, H.; Schaller, D.; Gandini, M.A.; Kaplan, O.; Gambeta, E.; Zhang, F.X.; Çelebier, M.; Tahir, M.N.; Zamponi, G.W.; Wolber, G.; et al. Discovery of Michael acceptor containing 1,4-dihydropyridines as first covalent inhibitors of L-/T-type calcium channels. Bioorganic Chem. 2019, 91, 103187. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, S.; Edraki, N.; Firuzi, O.; Khoshneviszadeh, M.; Miri, R. 5-Oxo-hexahydroquinoline: An attractive scaffold with diverse biological activities. Mol. Divers. 2019, 23, 471–508. [Google Scholar] [CrossRef]
- Sahiba, N.; Sethiya, A.; Soni, J.; Agarwal, S. Acridine-1,8-diones: Synthesis and Biological Applications. ChemistrySelect 2021, 6, 2210–2251. [Google Scholar] [CrossRef]
- Bai, C.-B.; Wang, N.-X.; Wang, Y.-J.; Lan, X.-W.; Xing, Y.; Wen, J.-L. A new oxidation system for the oxidation of Hantzsch-1,4-dihydropyridines and polyhydroquinoline derivatives under mild conditions. RSC Adv. 2015, 5, 100531–100534. [Google Scholar] [CrossRef]
- Abdel-Mohsen, H.T.; Conrad, J.; Beifuss, U. Laccase-catalyzed oxidation of Hantzsch 1,4-dihydropyridines to pyridines and a new one pot synthesis of pyridines. Green Chem. 2012, 14, 2686–2690. [Google Scholar] [CrossRef]
- Triggle, D.J. Calcium channel antagonists: Clinical uses—Past, present and future. Biochem. Pharmacol. 2007, 74, 1–9. [Google Scholar] [CrossRef]
- Wang, B.; Hu, Y.; Hu, H. The Aromatization of Hantzsch 1,4-Dihydropyridines by Tetrakis-Pyridine Cobalt (II) Dichromate (TPCD). Synth. Commun. 1999, 29, 4193–4199. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, A. An Expeditious Oxidative Aromatization of Hantzsch 1,4-Dihydropyridines to Pyridines Using Cetyltrimethylammonium Peroxodisulfate: A Phase Transferring Oxidant. Bull. Korean Chem. Soc. 2010, 31, 2299–2303. [Google Scholar] [CrossRef] [Green Version]
- Mirza-Aghayan, M.; Boukherroub, R.; Nemati, M.; Rahimifard, M. Graphite oxide mediated oxidative aromatization of 1,4-dihydropyridines into pyridine derivatives. Tetrahedron Lett. 2012, 53, 2473–2475. [Google Scholar] [CrossRef]
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Frasconi, M.; Favero, G.; Boer, H.; Koivula, A.; Mazzei, F. Kinetic and biochemical properties of high and low redox potential laccases from fungal and plant origin. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Cambria, M.T.; Minniti, Z.; Librando, V.; Cambria, A. Degradation of Polycyclic Aromatic Hydrocarbons by Rigidoporus lignosus and its Laccase in the Presence of Redox Mediators. Appl. Biochem. Biotechnol. 2008, 149, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shariati, M.; Imanzadeh, G.; Rostami, A.; Ghoreishy, N.; Kheirjou, S. Application of laccase/DDQ as a new bioinspired catalyst system for the aerobic oxidation of tetrahydroquinazolines and Hantzsch 1,4-dihydropyridines. Comptes Rendus Chim. 2019, 22, 337–346. [Google Scholar] [CrossRef]
- Khaledian, D.; Rostami, A.; Zarei, S.A.; Mohammadi, B. Aerobic oxidative aromatization of Hantzsch 1,4-dihydropyridines via an anomeric-based oxidation in the presence of Laccase enzyme/4-Phenyl urazole as a cooperative catalytic oxidation system. J. Iran. Chem. Soc. 2019, 16, 1871–1878. [Google Scholar] [CrossRef]
- Simić, S.; Jeremic, S.; Djokic, L.; Božić, N.; Vujčić, Z.; Lončar, N.; Senthamaraikannan, R.; Babu, R.; Opsenica, I.M.; Nikodinovic-Runic, J. Development of an efficient biocatalytic system based on bacterial laccase for the oxidation of selected 1,4-dihydropyridines. Enzym. Microb. Technol. 2020, 132, 109411. [Google Scholar] [CrossRef]
- Kurniawati, S.; Nicell, J.A. Efficacy of mediators for enhancing the laccase-catalyzed oxidation of aqueous phenol. Enzym. Microb. Technol. 2007, 41, 353–361. [Google Scholar] [CrossRef]
- Piontek, K.; Antorini, M.; Choinowski, T. Crystal structure of a laccase from the fungus Trametes versicolor at 1.90-A resolution containing a full complement of coppers. J. Biol. Chem. 2002, 277, 37663–37669. [Google Scholar] [CrossRef] [Green Version]
- Tadesse, M.A.; D’Annibale, A.; Galli, C.; Gentili, P.; Sergi, F. An assessment of the relative contributions of redox and steric issues to laccase specificity towards putative substrates. Org. Biomol. Chem. 2008, 6, 868–878. [Google Scholar] [CrossRef]
- Durão, P.; Chen, Z.; Fernandes, A.T.; Hildebrandt, P.; Murgida, D.H.; Todorovic, S.; Pereira, M.M.; Melo, E.P.; Martins, L.O. Copper incorporation into recombinant CotA laccase from Bacillus subtilis: Characterization of fully copper loaded enzymes. J. Biol. Inorg. Chem. JBIC Publ. Soc. Biol. Inorg. Chem. 2008, 13, 183–193. [Google Scholar] [CrossRef]
- Zouraris, D.; Kiafi, S.; Zerva, A.; Topakas, E.; Karantonis, A. FTacV study of electroactive immobilized enzyme/free substrate reactions: Enzymatic catalysis of epinephrine by a multicopper oxidase from Thermothelomyces thermophila. Bioelectrochemistry 2020, 134, 107538. [Google Scholar] [CrossRef] [PubMed]
- Gramss, G. Reappraising a Controversy: Formation and Role of the Azodication (ABTS2+) in the Laccase-ABTS Catalyzed Breakdown of Lignin. Fermentation 2017, 3, 27. [Google Scholar] [CrossRef]
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase-mediator systems and their applications: A review. Appl. Biochem. Microbiol. 2007, 43, 523–535. [Google Scholar] [CrossRef]
- Hilgers, R.; Vincken, J.-P.; Gruppen, H.; Kabel, M.A. Laccase/Mediator Systems: Their Reactivity toward Phenolic Lignin Structures. ACS Sustain. Chem. Eng. 2018, 6, 2037–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerva, A.; Koutroufini, E.; Kostopoulou, I.; Detsi, A.; Topakas, E. A novel thermophilic laccase-like multicopper oxidase from Thermothelomyces thermophila and its application in the oxidative cyclization of 2,3,4-trihydroxychalcone. New Biotechnol. 2019, 49, 10–18. [Google Scholar] [CrossRef]
- Johannes, C.; Majcherczyk, A. Natural Mediators in the Oxidation of Polycyclic Aromatic Hydrocarbons by Laccase Mediator Systems. J. Appl. Environ. Microbiol. 2000, 66, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Galli, C.; Gentili, P.; Jolivalt, C.; Madzak, C.; Vadalà, R. How is the reactivity of laccase affected by single-point mutations? Engineering laccase for improved activity towards sterically demanding substrates. Appl. Microbiol. Biotechnol. 2011, 91, 123–131. [Google Scholar] [CrossRef]
- Sitarz, A.K.; Mikkelsen, J.D.; Meyer, A.S. Structure, functionality and tuning up of laccases for lignocellulose and other industrial applications. Crit. Rev. Biotechnol. 2016, 36, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Subrahmanyam, V.V.; Kolachana, P.; Smith, M.T. Metabolism of hydroquinone by human myeloperoxidase: Mechanisms of stimulation by other phenolic compounds. Arch. Biochem. Biophys. 1991, 286, 76–84. [Google Scholar] [CrossRef]
- Xu, F.; Shin, W.; Brown, S.H.; Wahleithner, J.A.; Sundaram, U.M.; Solomon, E.I. A study of a series of recombinant fungal laccases and bilirubin oxidase that exhibit significant differences in redox potential, substrate specificity, and stability. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1996, 1292, 303–311. [Google Scholar] [CrossRef]
- Mehra, R.; Muschiol, J.; Meyer, A.S.; Kepp, K.P. A structural-chemical explanation of fungal laccase activity. Sci. Rep. 2018, 8, 17285. [Google Scholar] [CrossRef] [PubMed]
- Glazunova, O.A.; Trushkin, N.A.; Moiseenko, K.V.; Filimonov, I.S.; Fedorova, T.V. Catalytic Efficiency of Basidiomycete Laccases: Redox Potential versus Substrate-Binding Pocket Structure. Catalysts 2018, 8, 152. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.-N.; Zhao, M. A simple strategy for extracellular production of CotA laccase in Escherichia coli and decolorization of simulated textile effluent by recombinant laccase. Appl. Microbiol. Biotechnol. 2017, 101, 685–696. [Google Scholar] [CrossRef]
- Chen, Y.C. Beware of docking! Trends Pharmacol. Sci. 2015, 36, 78–95. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Relative Energy (kcal/mol) |
|---|---|
| DHP1 | 0.0 a |
| DHP1-R | 750.8 |
| DHP2 | 0.0 b |
| DHP2-R | 763.2 |
| DHP3 | 0.0 c |
| DHP3-R | 784.4 |
| DHP4 | 0.0 d |
| DHP4-R | 763.6 |
| DHP5 | 0.0 e |
| DHP5-R | 765.8 |
| DHP6 | 0.0 f |
| DHP6-R | 763.3 |
| Substrate * Enzyme | DHP1 | DHP2 | DHP3 | DHP5 |
|---|---|---|---|---|
| TvLacc | 61.8 | 45.9 | 100 | 52.8 |
| BacillusLacc | 54.7 | 63.8 | 94.1 | 43.6 |
| Novozym 51003 | 67.2 | 79.4 | 94.1 | 37.7 |
| TtLMCO1 | 79.4 | 76.6 | 100 | 21.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milovanovic, J.; Gündüz, M.G.; Zerva, A.; Petkovic, M.; Beskoski, V.; Thomaidis, N.S.; Topakas, E.; Nikodinovic-Runic, J. Synthesis and Laccase-Mediated Oxidation of New Condensed 1,4-Dihydropyridine Derivatives. Catalysts 2021, 11, 727. https://doi.org/10.3390/catal11060727
Milovanovic J, Gündüz MG, Zerva A, Petkovic M, Beskoski V, Thomaidis NS, Topakas E, Nikodinovic-Runic J. Synthesis and Laccase-Mediated Oxidation of New Condensed 1,4-Dihydropyridine Derivatives. Catalysts. 2021; 11(6):727. https://doi.org/10.3390/catal11060727
Chicago/Turabian StyleMilovanovic, Jelena, Miyase Gözde Gündüz, Anastasia Zerva, Milos Petkovic, Vladimir Beskoski, Nikolaos S. Thomaidis, Evangelos Topakas, and Jasmina Nikodinovic-Runic. 2021. "Synthesis and Laccase-Mediated Oxidation of New Condensed 1,4-Dihydropyridine Derivatives" Catalysts 11, no. 6: 727. https://doi.org/10.3390/catal11060727