Biocatalytic Synthesis of a Novel Bioactive Ginsenoside Using UDP-Glycosyltransferase from Bacillus subtilis 168


Abstract
:1. Introduction
2. Results and Discussion
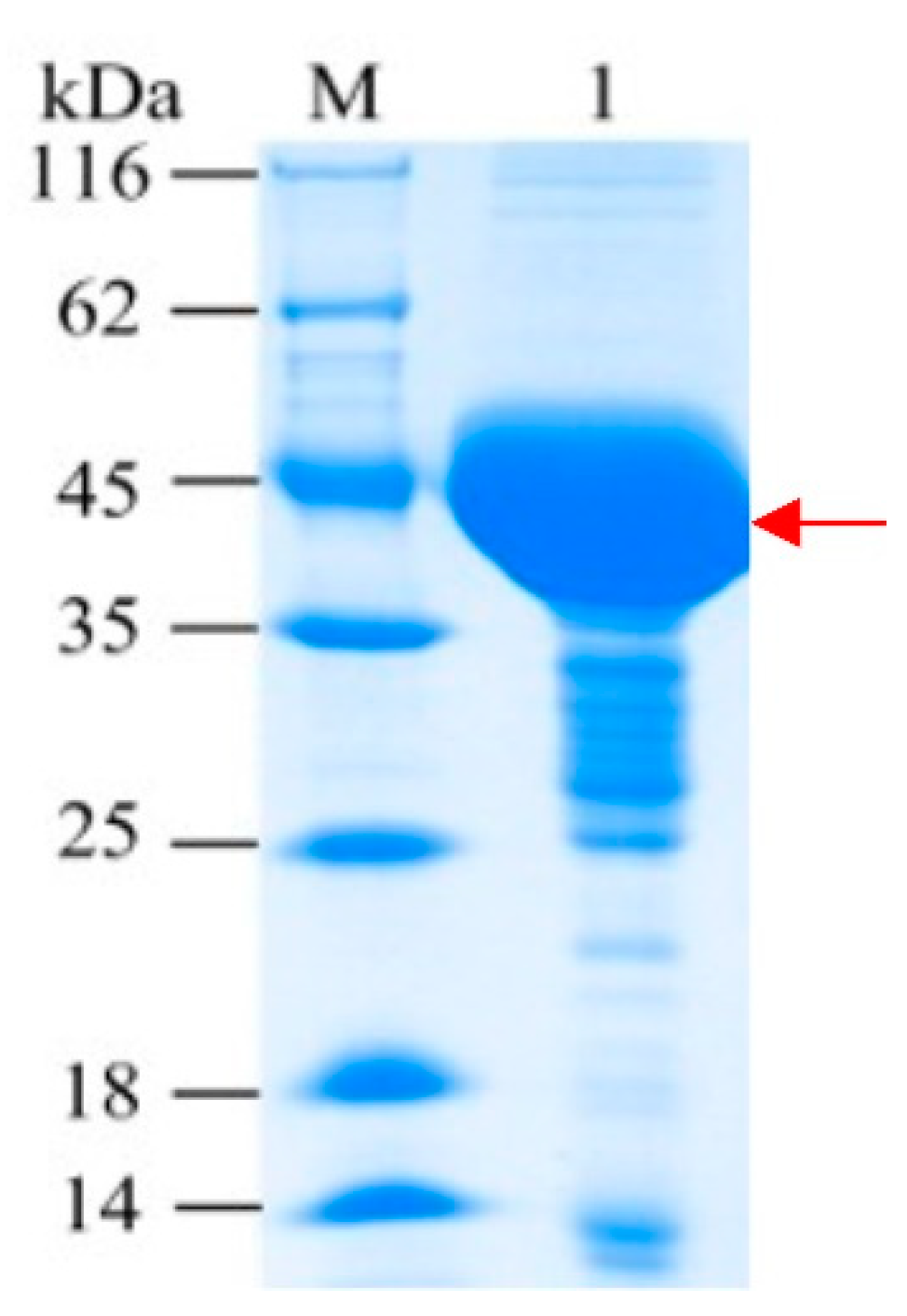
2.1. Expression and Purification of Bs-YjiC
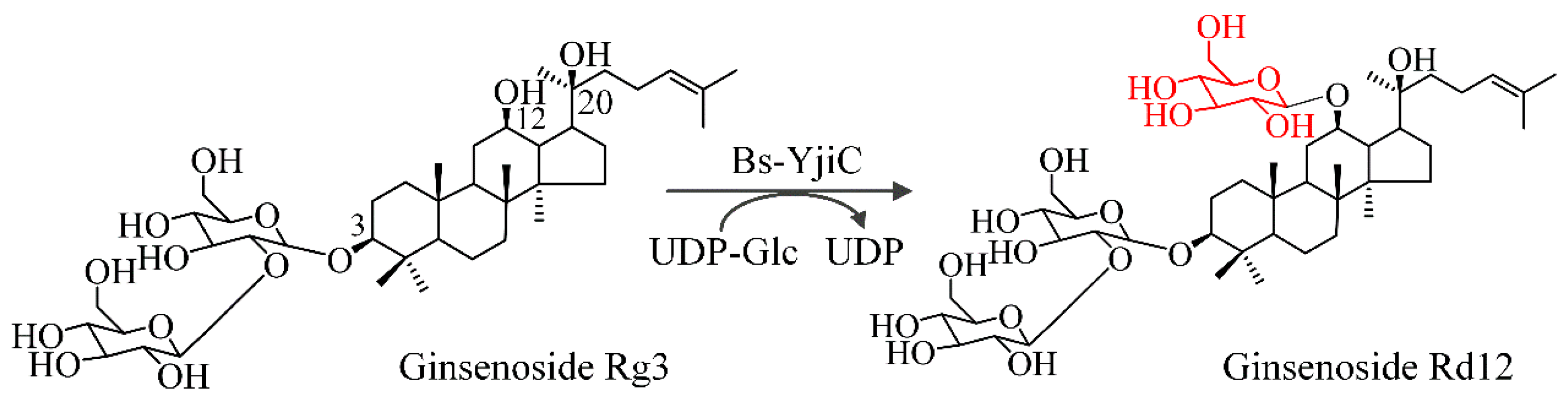
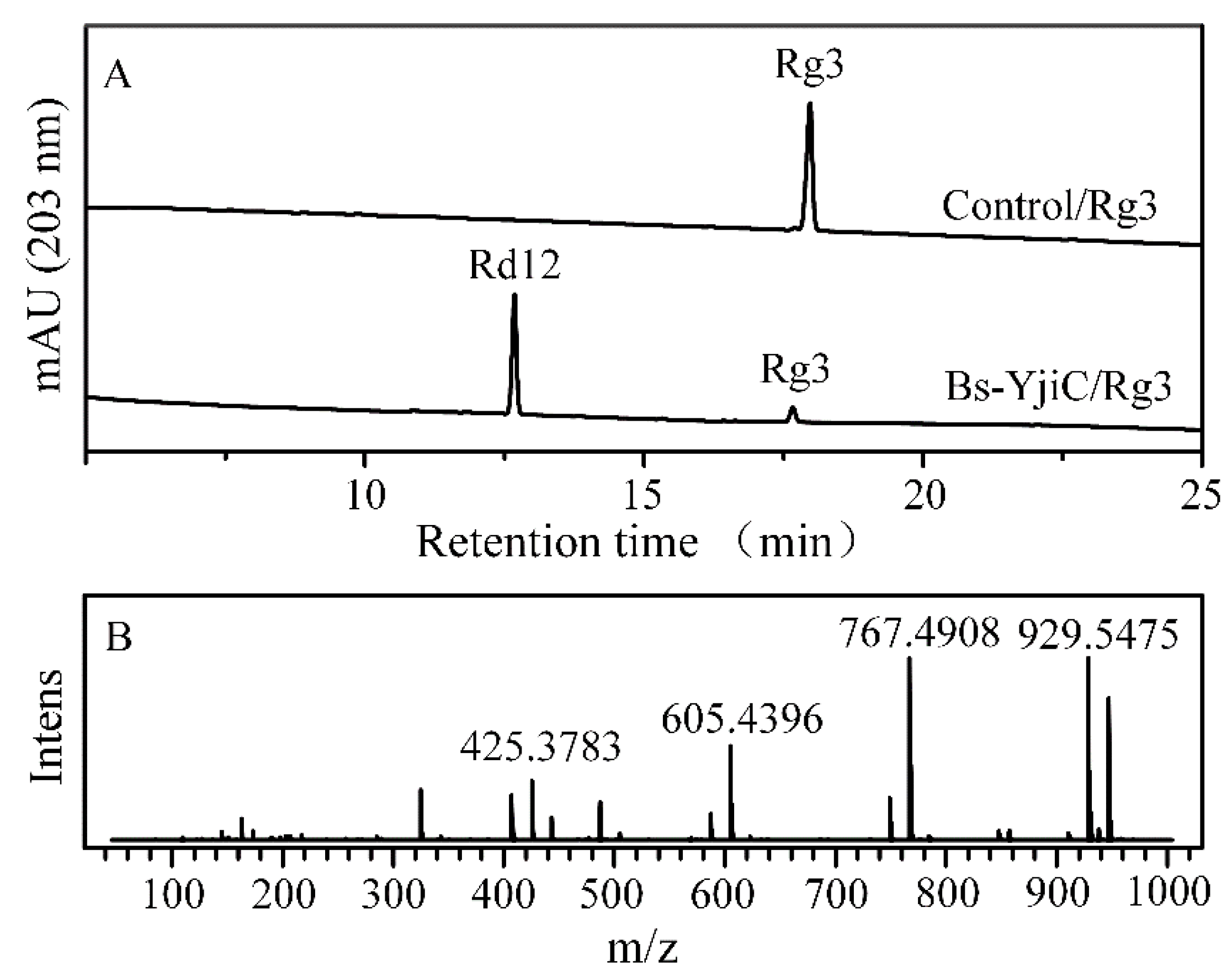
2.2. Glycosylation of Ginsenoside Rg3 with Bs-YjiC
2.3. Aqueous Solubility of Ginsenoside Rd12
2.4. In Vitro Cytotoxicity of Ginsenoside Rd12
3. Materials and Methods
3.1. Chemicals, Reagents, and Cancer Cell Lines
3.2. Heterologous Expression and Enzymatic Activity of Bs-YjiC
3.3. Kinetic Parameters of Bs-YjiC toward Ginsenoside Rg3
3.4. Water Solubility Determination
3.5. Cell Viability Assay
3.6. HPLC and HPLC-ESI-MS Analysis of the Glycosylated Product
3.7. Structural Elucidation of Ginsenoside Rd12 by NMR Spectroscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kim, S.-A.; Shin, K.-C.; Oh, D.-K. Complete biotransformation of protopanaxadiol-type ginsenosides into 20-O-β-glucopyranosyl-20(S)-protopanaxadiol by permeabilized recombinant Escherichia coli cells coexpressing β-glucosidase and chaperone genes. J. Agric. Food Chem. 2019, 67, 8393–8401. [Google Scholar] [CrossRef]
- Wan, X.; Ren, Y.; Sun, X.; Liu, S. Classification and analysis of set prescription preparations containing Panax ginseng in 2015 edition of Chinese Pharmacopoeia (Part I). China Pharm. 2018, 29, 69–73. [Google Scholar]
- Leung, K.W.; Wong, A.S. Ginseng and male reproductive function. Spermatogenesis 2013, 3, e26391. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-Y.; Kim, H.-J.; Kwon, Y.-S.; Choi, Y.-E. The Cyt P450 enzyme CYP716A47 catalyzes the formation of protopanaxadiol from dammarenediol-II during ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol. 2011, 52, 2062–2073. [Google Scholar] [CrossRef]
- Seki, H.; Tamura, K.; Muranaka, T. P450s and UGTs: Key players in the structural diversity of triterpenoid saponins. Plant Cell Physiol. 2015, 56, 1463–1471. [Google Scholar] [CrossRef]
- Yu, T.; Yang, Y.; Kwak, Y.-S.; Song, G.G.; Kim, M.-Y.; Rhee, M.H.; Cho, J.Y. Ginsenoside Rc from Panax ginseng exerts anti-inflammatory activity by targeting TANK-binding kinase 1/interferon regulatory factor-3 and p38/ATF-2. J. Ginseng Res. 2017, 41, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Kim, J.H.; Kim, J.-H.; Yi, Y.-S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.; Chen, W.; Xie, J. Antihyperglycemic effects of ginseng and possible mechanisms. Drugs Future 2008, 33, 507–514. [Google Scholar] [CrossRef]
- Tan, B.K.; Vanitha, J. Immunomodulatory and antimicrobial effects of some traditional Chinese medicinal herbs: A review. Curr. Med. Chem. 2004, 11, 1423–1430. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Li, J.; Yang, J.; Zhu, Y.; Men, Y.; Zeng, Y.; Cai, Y.; Dong, C.; Dai, Z.; Zhang, X. Use of a promiscuous glycosyltransferase from Bacillus subtilis 168 for the enzymatic synthesis of novel protopanaxatriol-type ginsenosides. J. Agric. Food Chem. 2018, 66, 943–949. [Google Scholar] [CrossRef]
- Wei, W.; Wang, P.; Wei, Y.; Liu, Q.; Yang, C.; Zhao, G.; Yue, J.; Yan, X.; Zhou, Z. Characterization of Panax ginseng UDP-glycosyltransferases catalyzing protopanaxatriol and biosyntheses of bioactive ginsenosides F1 and Rh1 in metabolically engineered yeasts. Mol. Plant 2015, 8, 1412–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Fan, Y.; Wei, W.; Wang, P.; Liu, Q.; Wei, Y.; Zhang, L.; Zhao, G.; Yue, J.; Zhou, Z. Production of bioactive ginsenoside compound K in metabolically engineered yeast. Cell Res. 2014, 24, 770–773. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xue, J.; Min, J.; Qin, L.; Zhang, J.; Dai, L. Biocatalytic synthesis of ginsenoside Rh2 using Arabidopsis thaliana glucosyltransferase-catalyzed coupled reactions. J. Biotechnol. 2020, 309, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Gantt, R.W.; Goff, R.D.; Williams, G.J.; Thorson, J.S. Probing the aglycon promiscuity of an engineered glycosyltransferase. Angew. Chem. Int. Ed. 2008, 47, 8889–8892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Zhang, J.; Shao, Y.; Xu, Y.; Ge, H.; Yu, B.; Wang, W. Enzyme-Catalyzed Glycosylation of Curcumin and Its Analogues by Glycosyltransferases from Bacillus subtilis ATCC 6633. Catalysts 2019, 9, 734. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.P.; Bashyal, P.; Parajuli, P.; Yamaguchi, T.; Sohng, J.K. Two trifunctional leloir glycosyltransferases as biocatalysts for natural products glycodiversification. Org. Lett. 2019, 21, 8058–8064. [Google Scholar] [CrossRef]
- Chiang, C.-M.; Wang, T.-Y.; Yang, S.-Y.; Wu, J.-Y.; Chang, T.-S. Production of new isoflavone glucosides from glycosylation of 8-hydroxydaidzein by glycosyltransferase from Bacillus subtilis ATCC 6633. Catalysts 2018, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.M.; Min, J.; Qu, Y.Y.; Zhang, X.; Zhang, J.K.; Yu, X.J.; Dai, L.H. Biocatalytic synthesis of calycosin-7-O-β-D-glucoside with UDP-glucose regeneration system. Catalysts 2020, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-S.; Chiang, C.-M.; Kao, Y.-H.; Wu, J.-Y.; Wu, Y.-W.; Wang, T.-Y. A new triterpenoid glucoside from a novel acidic glycosylation of ganoderic acid a via recombinant glycosyltransferase of Bacillus subtilis. Molecules 2019, 24, 3457. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.P.; Li, T.F.; Kim, E.-H.; Yamaguchi, T.; Park, Y.I.; Kim, J.S.; Sohng, J.K. Enzymatic synthesis of novel phloretin glucosides. Appl. Environ. Microbiol. 2013, 79, 3516–3521. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Li, J.; Yang, J.; Men, Y.; Zeng, Y.; Cai, Y.; Sun, Y. Enzymatic synthesis of novel glycyrrhizic acid glucosides using a promiscuous Bacillus glycosyltransferase. Catalysts 2018, 8, 615. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Li, J.; Yao, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Yang, J.; Sun, Y. Exploiting the aglycon promiscuity of glycosyltransferase Bs-YjiC from Bacillus subtilis and its application in synthesis of glycosides. J. Biotechnol. 2017, 248, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Liu, C.; Li, J.; Dong, C.; Yang, J.; Dai, Z.; Zhang, X.; Sun, Y. One-pot synthesis of ginsenoside Rh2 and bioactive unnatural ginsenoside by coupling promiscuous glycosyltransferase from Bacillus subtilis 168 to sucrose synthase. J. Agric. Food Chem. 2018, 66, 2830–2837. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wei, Y.; Fan, Y.; Liu, Q.; Wei, W.; Yang, C.; Zhang, L.; Zhao, G.; Yue, J.; Yan, X. Production of bioactive ginsenosides Rh2 and Rg3 by metabolically engineered yeasts. Metab. Eng. 2015, 29, 97–105. [Google Scholar] [CrossRef]
- Dai, L.; Liu, C.; Zhu, Y.; Zhang, J.; Men, Y.; Zeng, Y.; Sun, Y. Functional characterization of cucurbitadienol synthase and triterpene glycosyltransferase involved in biosynthesis of mogrosides from Siraitia grosvenorii. Plant Cell Physiol. 2015, 56, 1172–1182. [Google Scholar] [CrossRef]
- Wang, P.; Wei, W.; Ye, W.; Li, X.; Zhao, W.; Yang, C.; Li, C.; Yan, X.; Zhou, Z. Synthesizing ginsenoside Rh2 in Saccharomyces cerevisiae cell factory at high-efficiency. Cell Discov. 2019, 5, 1–14. [Google Scholar] [CrossRef]
- Lu, J.; Yao, L.; Li, J.-X.; Liu, S.-J.; Hu, Y.-Y.; Wang, S.-H.; Liang, W.-X.; Huang, L.-Q.; Dai, Y.-J.; Wang, J. Characterization of UDP-glycosyltransferase involved in biosynthesis of ginsenosides Rg1 and Rb1 and identification of critical conserved amino acid residues for its function. J. Agric. Food Chem. 2018, 66, 9446–9455. [Google Scholar] [CrossRef]
- Jung, S.-C.; Kim, W.; Park, S.C.; Jeong, J.; Park, M.K.; Lim, S.; Lee, Y.; Im, W.-T.; Lee, J.H.; Choi, G. Two ginseng UDP-glycosyltransferases synthesize ginsenoside Rg3 and Rd. Plant Cell Physiol. 2014, 55, 2177–2188. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Dou, X.; Chen, R.; Chen, D.; Fang, C.; Xiao, Z.; Dai, J. Two novel fungal phenolic UDP glycosyltransferases from Absidia coerulea and Rhizopus japonicus. Appl. Environ. Microbiol. 2017, 83, e03103–e03116. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Hou, Y.; Hamza, A.; Zhan, C.-G.; Bugni, T.S.; Thorson, J.S. Probing the regiospecificity of enzyme-catalyzed steroid glycosylation. Org. Lett. 2012, 14, 5424–5427. [Google Scholar] [CrossRef] [Green Version]
- Lepak, A.; Gutmann, A.; Kulmer, S.T.; Nidetzky, B. Creating a water-soluble resveratrol-based antioxidant by site-selective enzymatic glucosylation. ChemBioChem 2015, 16, 1870–1874. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| C | δC (ppm) | δH (ppm) |
|---|---|---|
| 1 | 36.69 | 1.44 (1H, m), 1.59 (1H, m) |
| 2 | 27.61 | 1.37 (1H, m), 1.67 (1H, m) |
| 3 | 91.26 | 3.23 (1H, br.s) |
| 4 | 40.56 | - |
| 5 | 57.48 | 0.79 (1H, m) |
| 6 | 19.19 | 1.41 (1H, m), 1.47 (1H, m) |
| 7 | 36.69 | 1.36 (1H, m), 1.47 (1H, m) |
| 8 | 38.12 | - |
| 9 | 53.15 | 1.69 (1H, overlapped) |
| 10 | 41.94 | - |
| 11 | 31.86 | 1.80 (1H, m), 2.04 (1H, m) |
| 12 | 79.39 | 3.92 (1H, m) |
| 13 | 47.30 | 1.10 (1H, m) |
| 14 | 54.96 | - |
| 15 | 28.65 | 1.26 (1H, m), 1.38 (1H, m) |
| 16 | 28.36 | 1.43 (1H, m), 2.03 (1H, m) |
| 17 | 51.14 | - |
| 18 | 17.44 | 0.98 (3H, s) |
| 19 | 16.74 | 0.96 (3H, s) |
| 20 | 74.72 | - |
| 21 | 26.21 | 1.15 (3H, s) |
| 22 | 35.75 | 1.32 (2H, m) |
| 23 | 23.28 | 1.18 (2H, m), 1.67 (1H, m) |
| 24 | 126.30 | 5.17 (1H, tr) |
| 25 | 131.80 | - |
| 26 | 25.92 | 1.71 (3H, s) |
| 27 | 17.77 | 1.65 (3H, s) |
| 28 | 27.61 | 1.10 (3H, s) |
| 29 | 16.71 | 0.89 (3H, s) |
| 30 | 16.21 | 1.07 (3H, s) |
| C3-O-inner-Glc-1′ | 105.38 | 4.47 (1H, d, J = 8.0 Hz) |
| 2′ | 81.11 | 3.60 (m) |
| 3′ | 77.59 | 3.38 (1H, overlapped) |
| 4′ | 71.55 | 3.13 (1H, overlapped) |
| 5′ | 78.26 | 3.27 (1H, m) |
| 6′ | 62.82 | 3.69 (1H, dd, J = 7.8, 4.2 Hz); 3.86 (1H, overlapped) |
| C3-O-outer-Glc-1″ | 104.50 | 4.70 (1H, d, J = 8.0 Hz) |
| 2″ | 76.27 | 3.14 (m) |
| 3″ | 77.81 | 3.38 (1H, overlapped) |
| 4″ | 70.92 | 3.36 (1H, overlapped) |
| 5″ | 78.33 | 3.24 (1H, m) |
| 6″ | 62.36 | 3.69 (1H, dd, J = 7.8, 4.2 Hz); 3.86 (1H, overlapped) |
| C12-O-inner-Glc-1‴ | 100.53 | 4.53 (1H, d, J = 8.0 Hz) |
| 2‴ | 75.08 | 3.13 (m) |
| 3‴ | 77.96 | 3.14 (1H, overlapped) |
| 4‴ | 71.86 | 3.20 (1H, overlapped) |
| 5‴ | 78.44 | 3.60 (1H, m) |
| 6‴ | 63.08 | 3.67 (1H, dd, J = 8.4, 3.0 Hz); 3.81 (1H, overlapped) |
| Compound | Lovo 1 | DMS53 2 | SNU719 3 |
|---|---|---|---|
| Rg3 | >400 | >400 | 359.4 ± 30.1 |
| Rd12 | 40.7 ± 5.9 | 46.4 ± 4.1 | 112.2 ± 11.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Li, H.; Qu, Y.; Zhang, X.; Zhang, J.; Dai, L. Biocatalytic Synthesis of a Novel Bioactive Ginsenoside Using UDP-Glycosyltransferase from Bacillus subtilis 168. Catalysts 2020, 10, 289. https://doi.org/10.3390/catal10030289
Hu Y, Li H, Qu Y, Zhang X, Zhang J, Dai L. Biocatalytic Synthesis of a Novel Bioactive Ginsenoside Using UDP-Glycosyltransferase from Bacillus subtilis 168. Catalysts. 2020; 10(3):289. https://doi.org/10.3390/catal10030289
Chicago/Turabian StyleHu, Yumei, Hao Li, Yingying Qu, Xiao Zhang, Juankun Zhang, and Longhai Dai. 2020. "Biocatalytic Synthesis of a Novel Bioactive Ginsenoside Using UDP-Glycosyltransferase from Bacillus subtilis 168" Catalysts 10, no. 3: 289. https://doi.org/10.3390/catal10030289