Effective Glycosylation of Cucurbitacin Mediated by UDP-Glycosyltransferase UGT74AC1 and Molecular Dynamics Exploration of Its Substrate Binding Conformations

Abstract
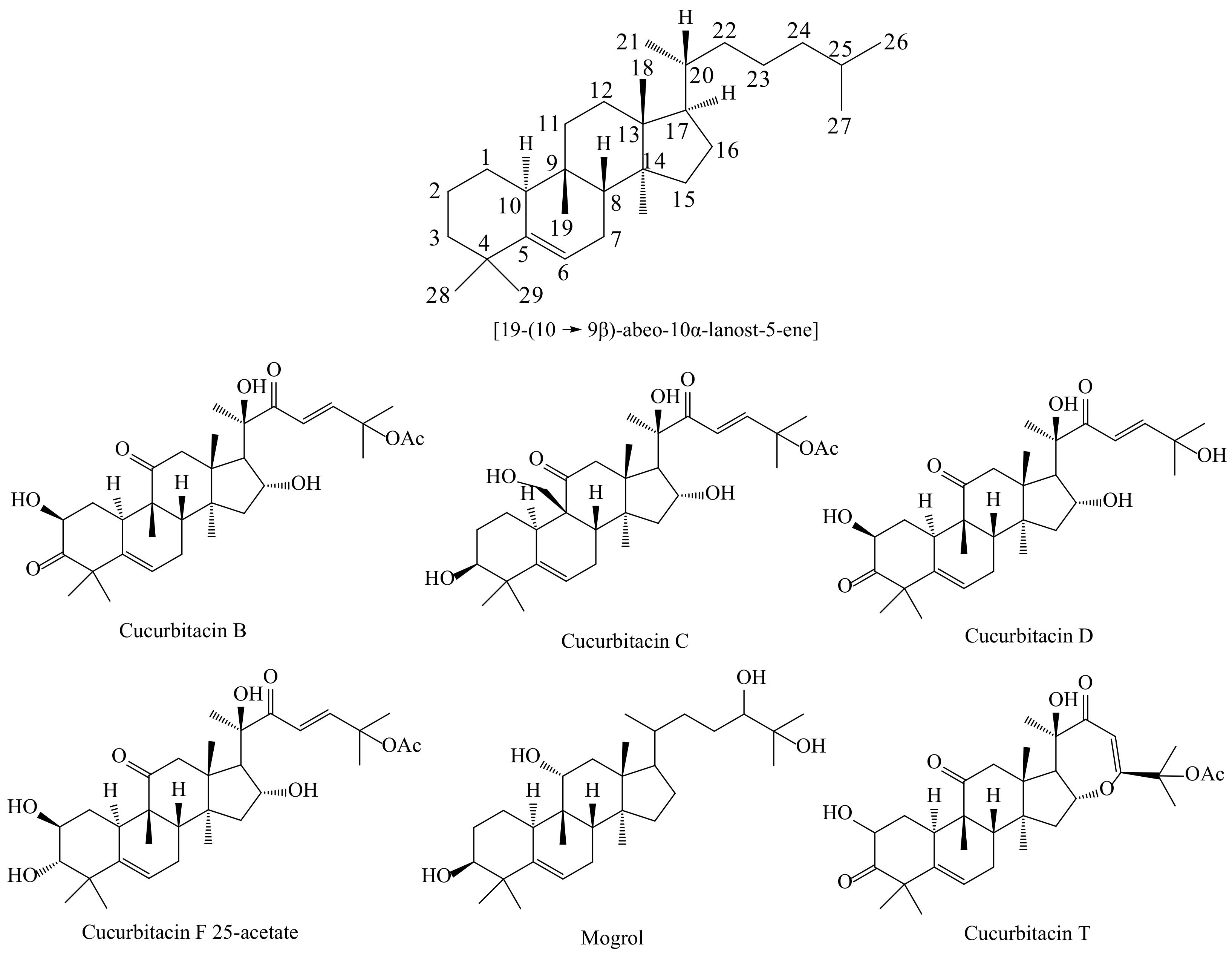
:1. Introduction
2. Results and Discussion
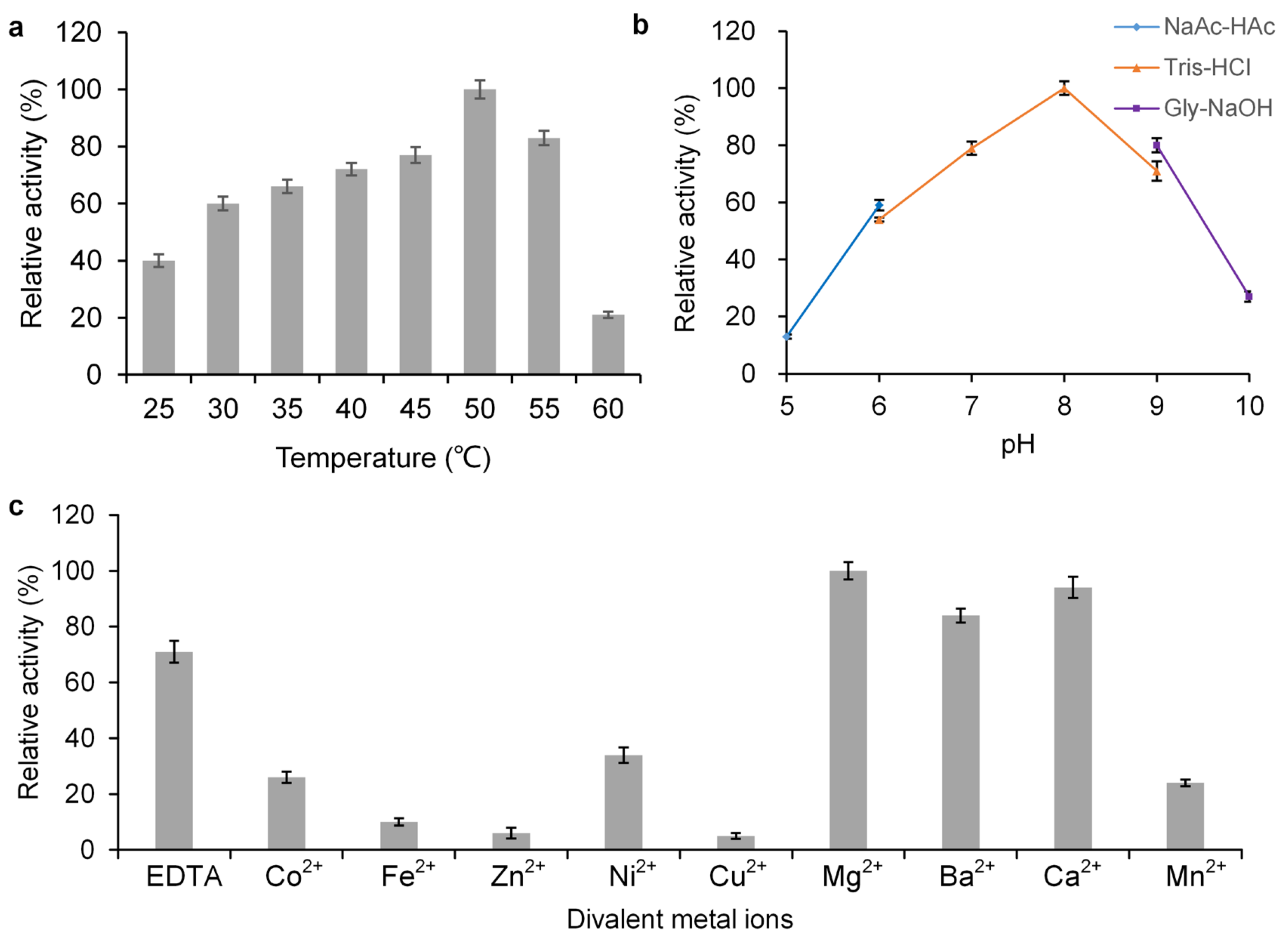
2.1. Characterization of GT-SM Subsection
2.2. Kinetic Analysis of GT-SM toward Cucurbitacin F 25-Acetate
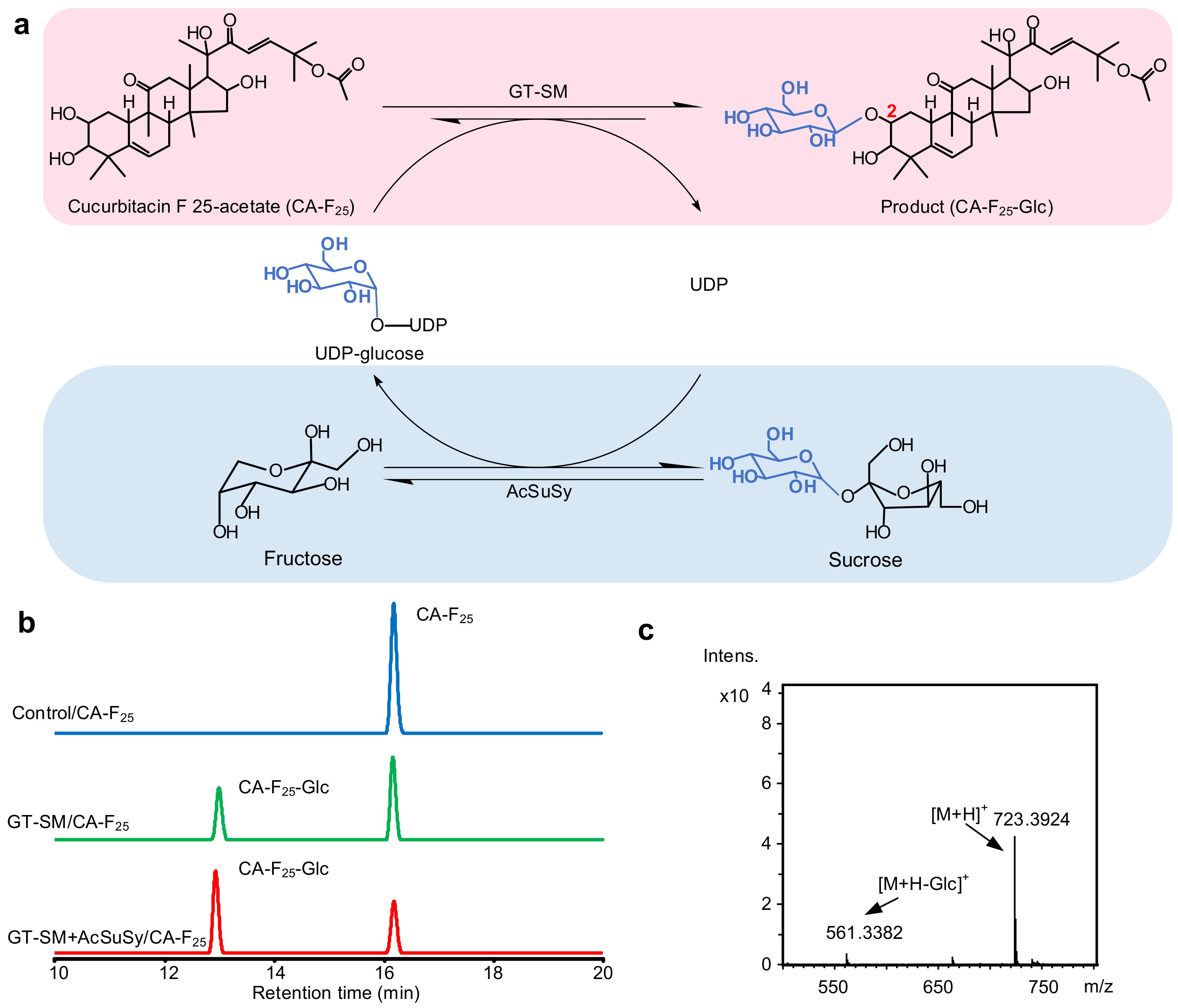
2.3. Glycosylation of Cucurbitacin F 25-Acetate by GT-SM
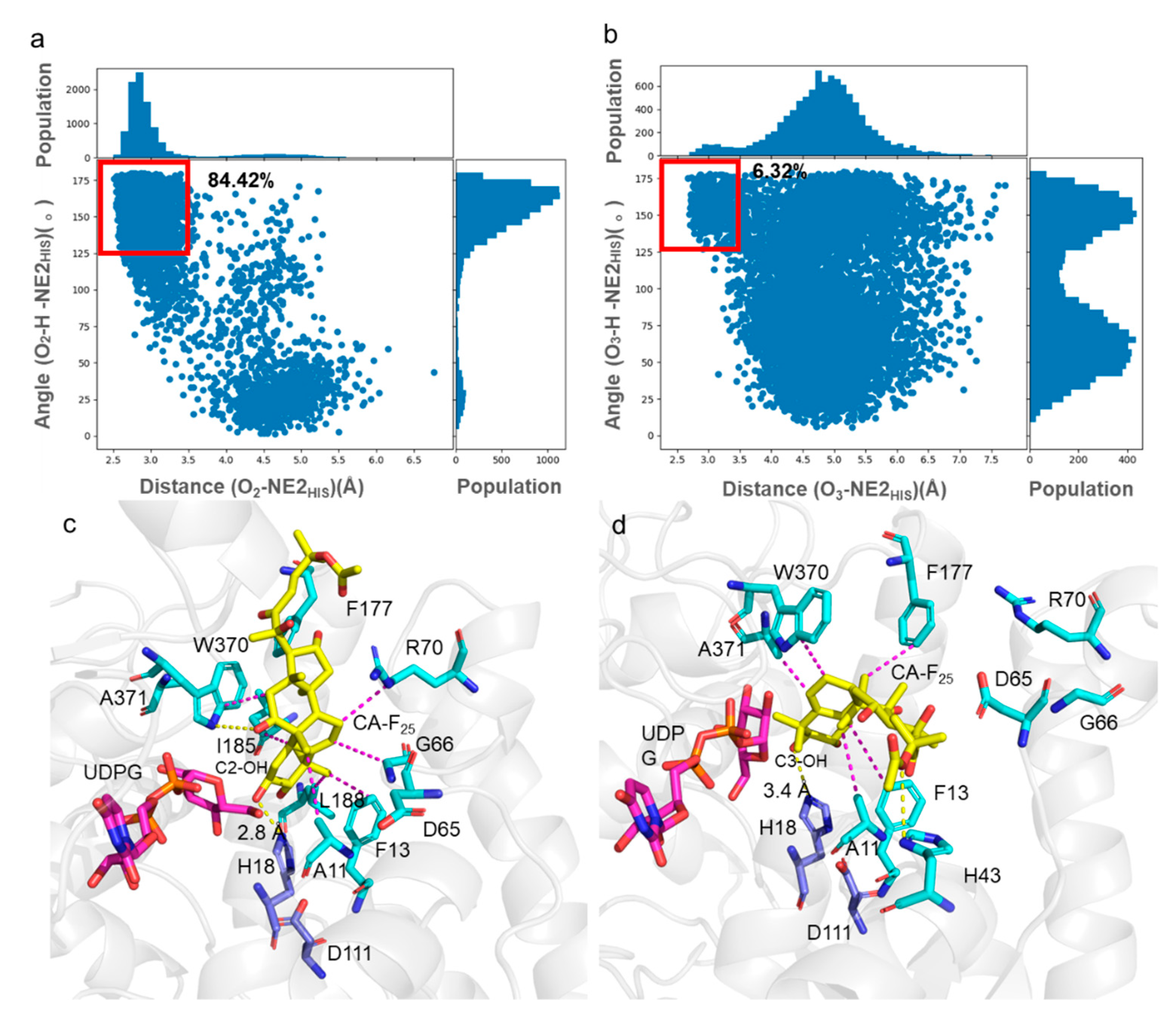
2.4. Molecular Basis for Recognition of GT-SM towards Cucurbitacin F 25-Acetate
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Expression and Purification of Glycosyltransferases
3.3. Activity Assay of Glycosyltransferase
3.4. Optimization of Glycosyltransferase Activity
3.5. Kinetic Analysis of Glycosyltransferase towards Cucurbitacin F 25-acetate
3.6. Molecular Dynamics Simulations
3.7. Glycosyltransferase/Sucrose Synthase Coupled Reaction
3.8. Purification and Structural Analysis of Glycoside
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Garg, S.; Kaul, S.C.; Wadhwa, R. Cucurbitacin B and cancer intervention: Chemistry, biology and mechanisms (Review). Int. J. Oncol. 2017, 52, 19–37. [Google Scholar] [CrossRef]
- Konopa, J.; Matuszkiewicz, A.; Hrabowska, M.; Onoszka, K. Cucurbitacines, cytotoxic and antitumor substances from Bryonia alba L. Part II: Biological studies. Arzneim. Forsch. 1974, 24, 1741. [Google Scholar] [CrossRef]
- Escandell, J.M.; Kaler, P.; Recio, M.C.; Sasazuki, T.; Shirasawa, S.; Augenlicht, L.; Ríos, J.-L.; Klampfer, L. Activated kRas protects colon cancer cells from cucurbitacin-induced apoptosis: The role of p53 and p21. Biochem. Pharmacol. 2008, 76, 198–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.; Chiu, M.H.; Nie, R.L.; Cordell, G.A.; Qiu, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. [Google Scholar] [CrossRef]
- Itkin, M.; Davidovich-Rikanati, R.; Cohen, S.; Portnoy, V.; Doron-Faigenboim, A.; Oren, E.; Freilich, S.; Tzuri, G.; Baranes, N.; Shen, S.; et al. The biosynthetic pathway of the nonsugar, high-intensity sweetener mogroside V from Siraitia grosvenorii. Proc. Natl. Acad. Sci. USA 2016, 113, E7619–E7628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Ma, Y.; Zhou, Y.; Zhang, H.; Duan, L.; Chen, H.; Zeng, J.; Zhou, Q.; Wang, S.; Gu, W.; et al. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science 2014, 346, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Xue, X.; Liu, Z.; Ma, Y.; Zeng, K.; Han, L.; Qi, J.; Ro, D.; Bak, S.; Huang, S.; et al. Developmentally regulated glucosylation of bitter triterpenoid in cucumber by the UDP-glucosyltransferase UGT73AM3. Mol. Plant 2017, 10, 1000–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.H.; Li, J.; Yang, J.G.; Men, Y.; Zeng, Y.; Cai, Y.; Sun, Y.X. Enzymatic synthesis of novel glycyrrhizic acid glucosides using a promiscuous bacillus glycosyltransferase. Catalysts 2018, 8, 615. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Jang, J.; Woo, M.; Ahn, J.S.; Kim, J.S.; Hong, Y. Enzymatic glycosylation of nonbenzoquinone geldanamycin analogs via Bacillus UDP-glycosyltransferase. Appl. Environ. Microbiol. 2012, 78, 7680–7686. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Sun, P.; Zhou, F.; Li, Y.; Chen, K.; Jia, H.; Yan, M.; Gong, D.; Ouyang, P. Synthesis of rebaudioside D, using glycosyltransferase UGTSL2 and in situ UDP-glucose regeneration. Food Chem. 2018, 259, 286–291. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, Q.; Feng, K.; Yu, X.; Xiong, A. DcMYB113, a root-specific R2R3-MYB, conditions anthocyanin biosynthesis and modification in carrot. Plant Biotechnol. J. 2020, 18, 1585–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.H.; Liu, C.; Zhu, Y.M.; Zhang, J.G.; Men, Y.; Zeng, Y.; Sun, Y.X. Functional characterization of cucurbitadienol synthase and triterpene glycosyltransferase involved in biosynthesis of mogrosides from Siraitia grosvenorii. Plant Cell Physiol. 2015, 56, 1172–1182. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Huang, S. Multi-omics data-driven investigations of metabolic diversity of plant triterpenoids. Plant. J. 2019, 97, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Liu, J.; Wu, H.; Wang, B.; Zhu, H.; Qiao, J. Glycosyltransferases: Mechanisms and Applications in Natural Product Development. Chem. Soc. Rev. 2015, 44, 8350–8374. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.G.; Mu, S.C.; Shang, N.; Liu, C.; Zhu, Y.M.; Cai, Y.; Liu, P.; Lin, J.; Liu, W.; et al. Efficient O-glycosylation of triterpenes enabled by protein engineering of plant glycosyltransferase UGT74AC1. ACS Catal. 2020, 10, 3629–3639. [Google Scholar] [CrossRef]
- Yang, T.; Zhang, J.; Ke, D.; Yang, W.; Tang, M.; Jiang, J.; Cheng, G.; Li, J.; Cheng, W.; Wei, Y.; et al. Hydrophobic recognition allows the glycosyltransferase UGT76G1 to catalyze its substrate in two orientations. Nat. Commun. 2019, 10, 3214. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; He, X.; Achnine, L.; Blount, J.W.; Dixon, R.A.; Wang, X. Crystal structures of a multifunctional triterpene/flavonoid glycosyltransferase from Medicago truncatula. Plant Cell 2005, 17, 3141–3154. [Google Scholar] [CrossRef] [Green Version]
- Brazier-Hicks, M.; Offen, W.A.; Gershater, M.C.; Revett, T.J.; Lim, E.-K.; Bowles, D.J.; Davies, G.J.; Edwards, R. Characterization and Engineering of the Bifunctional N- and O-Glucosyltransferase Involved in Xenobiotic Metabolism in Plants. Proc. Natl. Acad. Sci. USA 2007, 104, 20238–20243. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gao, Y.; Liu, X.; Guo, F.; Ma, C.; Liang, J.; Feng, X.; Li, C. Mining of sucrose synthases from Glycyrrhiza uralensis and their application in the construction of an efficient UDP-recycling system. J. Agric. Food Chem. 2019, 67, 11694–11702. [Google Scholar] [CrossRef]
- Bungaruang, L.; Gutmann, A.; Nidetzky, B. Leloir Glycosyltransferases and Natural Product Glycosylation: Biocatalytic Synthesis of the C-Glucoside Nothofagin, a Major Antioxidant of Redbush Herbal Tea. Adv. Synth. Catal. 2013, 355, 14–15. [Google Scholar] [CrossRef] [Green Version]
- Gutmann, A.; Bungaruang, L.; Weber, H.; Leypold, M.; Breinbauer, R.; Nidetzky, B. Towards the synthesis of glycosylated dihydrochalcone natural products using glycosyltransferase-catalysed cascade reactions. Green Chem. 2014, 16, 4417–4425. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.H.; Liu, C.; Li, J.; Dong, C.; Yang, J.G.; Dai, Z.; Zhang, X.; Sun, Y.X. One-pot synthesis of ginsenoside rh2 and bioactive unnatural ginsenoside by coupling promiscuous glycosyltransferase from Bacillus subtilis 168 to sucrose synthase. J. Agric. Food Chem. 2018, 66, 2830–2837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, S.; Jin, Y.; Dai, Y.; Chen, Y.; Wu, X. Efficient biocatalytic preparation of rebaudioside KA: Highly selective glycosylation coupled with UDPG regeneration. Sci. Rep. 2020, 10, 6230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuppner, H.; Wagner, H. New cucurbitacin glycosides from Picrorhiza kurrooa. Planta Med. 1989, 55, 559–563. [Google Scholar] [CrossRef]
- Pei, J.; Chen, A.; Sun, Q.; Zhao, L.; Cao, F.; Tang, F. Construction of a novel UDP-rhamnose regeneration system by a two-enzyme reaction system and application in glycosylation of flavonoid. Biochem. Eng. J. 2018, 2018, 33–42. [Google Scholar] [CrossRef]
- Dai, L.H.; Li, J.; Yao, P.; Zhu, Y.M.; Men, Y.; Zeng, Y.; Yang, J.G.; Sun, Y.X. Exploiting the aglycon promiscuity of glycosyltransferase Bs-YjiC from Bacillus subtilis and its application in synthesis of glycosides. J. Biotechnol. 2017, 2017, 69–76. [Google Scholar] [CrossRef]
- Xie, K.; Chen, R.; Li, J.; Wang, R.; Chen, D.; Dou, X.; Dai, J. Exploring the catalytic promiscuity of a new glycosyltransferase from Carthamus tinctorius. Org. Lett. 2014, 16, 4874–4877. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K. Glide: A New Approach for Rapid, Accurate Docking and Scoring. 1. Method and Assessment of Docking Accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- DeLano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Substrate | Km (µM) | kcat (s−1) | kcat/Km (s−1 µM−1) | References |
|---|---|---|---|---|---|
| GT-SM | CA-F25 | 39.34 ± 4.33 | 4.66 ± 0.54 | 120 ± 9.23 | This study |
| GT-SM | Mogrol | 9.59 ± 0.5 | 0.11 ± 0.03 | 11.47 ± 0.5 | [15] |
| UGT74AC1-M7 | Mogrol | 10.10 ± 0.3 | 0.86 ± 0.4 | 85.00 ± 6.6 | [15] |
| UGT73AM3 | Cucurbitacin C | 9.32 ± 1.84 | 5.33 × 10−5 | 5.72 × 10−6 | [7] |
| C | δ13C (ppm) | δ1H (ppm) |
|---|---|---|
| 1 | 32.1 | CH2, 0.90 (m), 1.77 (m) |
| 2 | 81.9 | CH, 3.50 (br,s) |
| 3 | 79.1 | CH, 2.87 (d, J = 9.0Hz) |
| 4 | 41.4 | - |
| 5 | 140.7 | - |
| 6 | 117.8 | CH, 5.64 (d, J = 5.4Hz) |
| 7 | 23.3 | CH2, 1.84 (m), 2.27 (m) |
| 8 | 42.2 | CH, 1.78 (m) |
| 9 | 47.6 | - |
| 10 | 32.5 | CH, 2.45 (d, J = 12.8Hz) |
| 11 | 212.8 | - |
| 12 | 48.6 | CH2, 2.34 (m), 3.27 (m) |
| 13 | 49.8 | - |
| 14 | 46.9 | - |
| 15 | 45.4 | CH2, 1.23 (m), 1.64 (m) |
| 16 | 69.2 | CH, 4.43 (m) |
| 17 | 59.0 | CH, 2.34 (m) |
| 18 | 19.8 | CH3, 0.72 (s) |
| 19 | 19.7 | CH3, 0.92 (s) |
| 20 | 78.5 | - |
| 21 | 25.4 | CH3, 1.26 (s) |
| 22 | 203.7 | - |
| 23 | 121.7 | CH, 6.80 (s) |
| 24 | 148.4 | CH, 6.81 (s) |
| 25 | 79.3 | - |
| 26 | 26.1 | CH3, 1.49 (s) |
| 27 | 26.1 | CH3, 1.50 (s) |
| 28 | 21.5 | CH3, 0.90 (s) |
| 29 | 24.6 | CH3, 1.11 (s) |
| 30 | 18.2 | CH3, 1.18 (s) |
| 31 | 169.3 | - |
| 32 | 21.7 | CH3, 1.97 (s) |
| 2-O-Glc | - | |
| C-1 | 104.6 | CH, 4.34 (d, J = 7.80Hz) |
| C-2 | 74.1 | CH, 2.97 (m) |
| C-3 | 76.2 | CH, 3.15 (m) |
| C-4 | 69.6 | CH, 3.08 (m) |
| C-5 | 76.6 | CH, 3.08 (m) |
| C-6 | 60.8 | CH2, 3.49 (dd, J1 = 5.2Hz, J2 = 11.4Hz), 3.63 (d, J = 11.2Hz) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, S.; Li, J.; Liu, C.; Zeng, Y.; Men, Y.; Cai, Y.; Chen, N.; Ma, H.; Sun, Y. Effective Glycosylation of Cucurbitacin Mediated by UDP-Glycosyltransferase UGT74AC1 and Molecular Dynamics Exploration of Its Substrate Binding Conformations. Catalysts 2020, 10, 1466. https://doi.org/10.3390/catal10121466
Mu S, Li J, Liu C, Zeng Y, Men Y, Cai Y, Chen N, Ma H, Sun Y. Effective Glycosylation of Cucurbitacin Mediated by UDP-Glycosyltransferase UGT74AC1 and Molecular Dynamics Exploration of Its Substrate Binding Conformations. Catalysts. 2020; 10(12):1466. https://doi.org/10.3390/catal10121466
Chicago/Turabian StyleMu, Shicheng, Jiao Li, Cui Liu, Yan Zeng, Yan Men, Yi Cai, Ning Chen, Hongwu Ma, and Yuanxia Sun. 2020. "Effective Glycosylation of Cucurbitacin Mediated by UDP-Glycosyltransferase UGT74AC1 and Molecular Dynamics Exploration of Its Substrate Binding Conformations" Catalysts 10, no. 12: 1466. https://doi.org/10.3390/catal10121466