
Identification and Characterization of Aptamers Targeting Ovarian Cancer Biomarker Human Epididymis Protein 4 for the Application in Urine




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aptamers and Proteins
2.2. Urine Preparation
2.3. HE4 Protein Immobilization
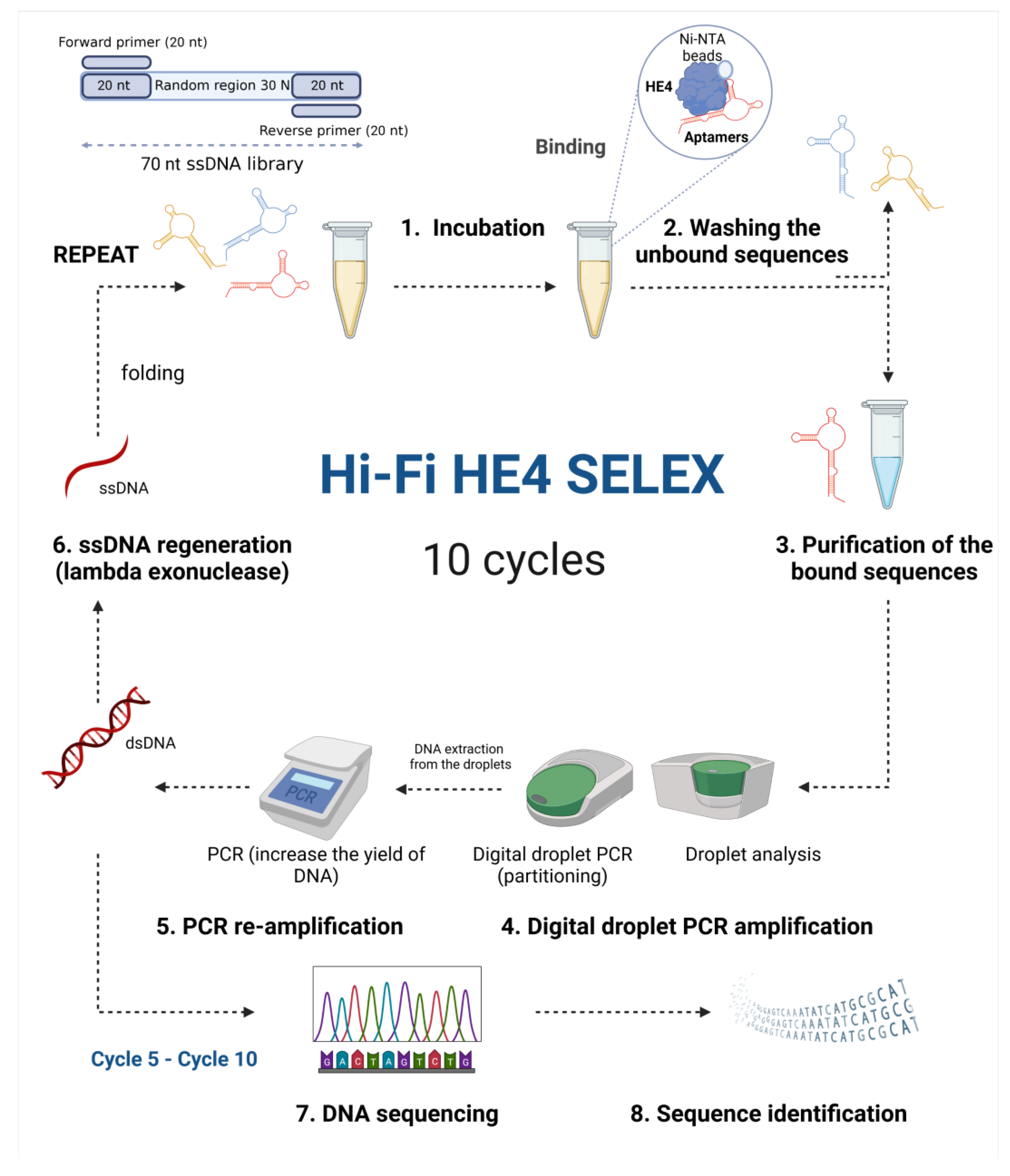
2.4. Digital Droplet PCR-Based High-Fidelity SELEX to Ovarian Cancer Biomarker HE4
2.5. DNA Sequencing and Bioinformatics
2.6. Aptamers-HE4 Protein Binding Analysis in Urine by Thermofluorimetry
3. Results
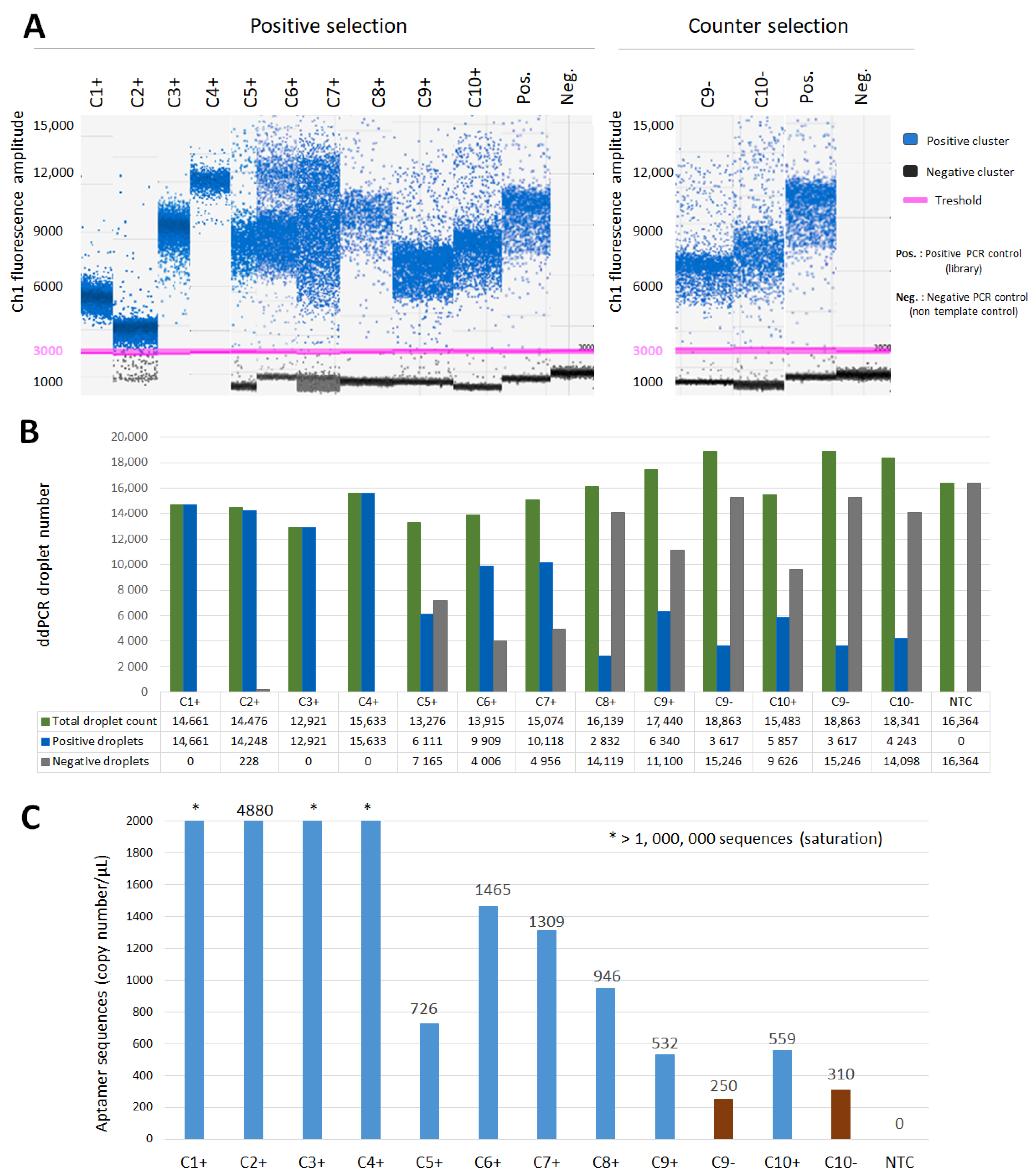
3.1. Selection and ddPCR Amplification of Anti-HE4 Diagnostic Aptamers
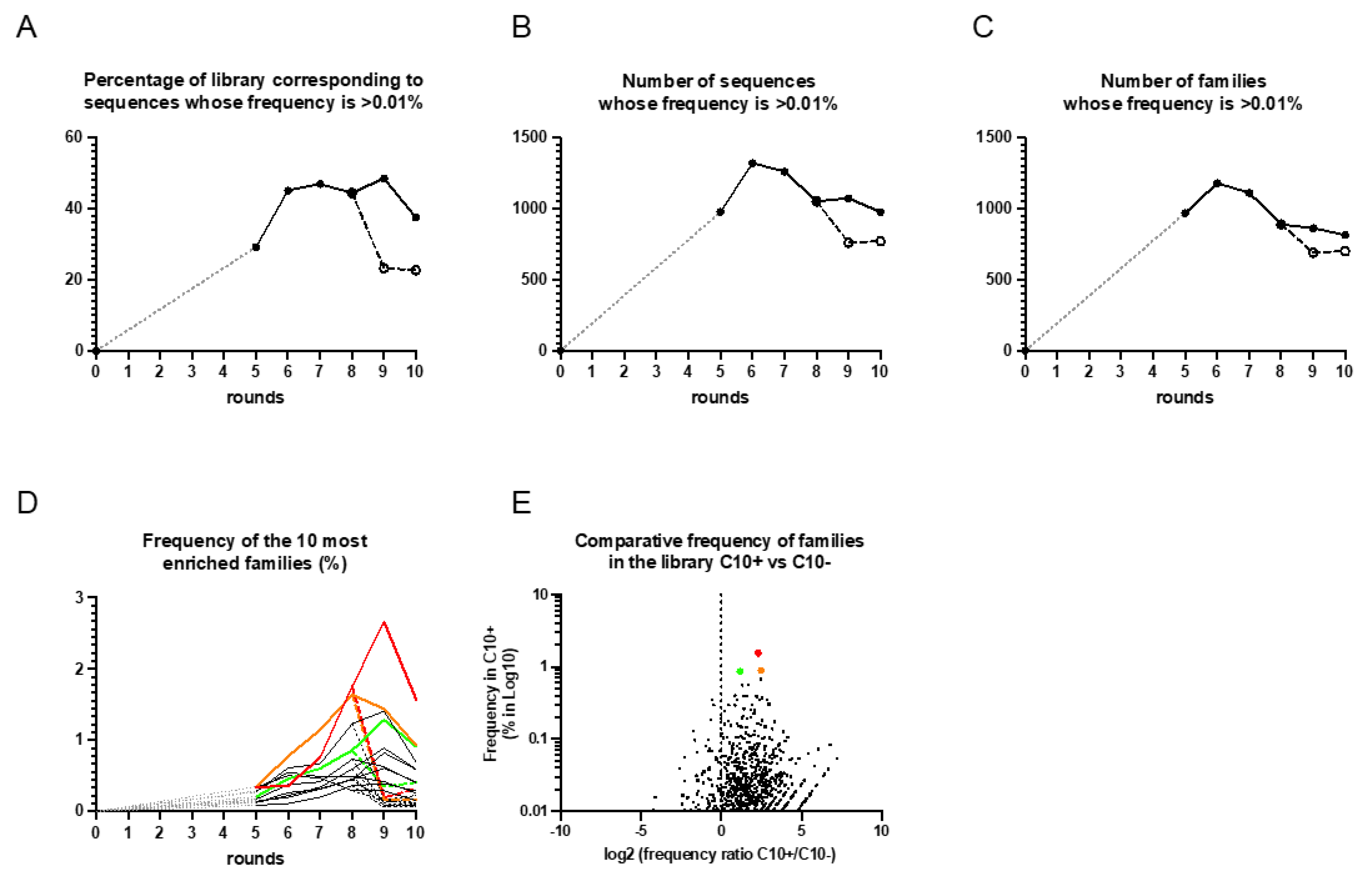
3.2. DNA Sequencing and Identification of the Enriched Aptamers to HE4
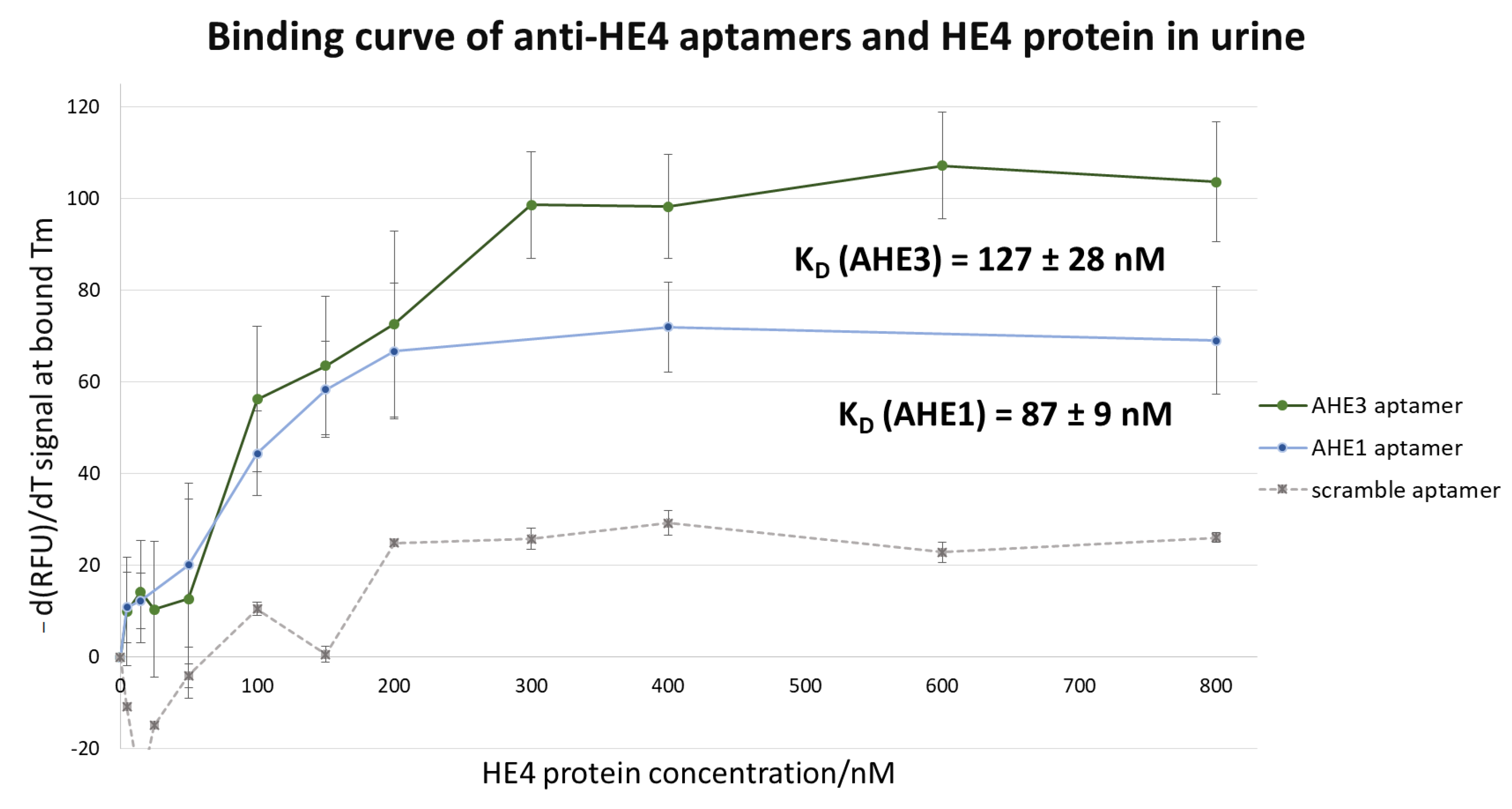
3.3. Thermofluorimetric Analysis of the Aptamers-HE4 Binding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Reid, F.; Bhatla, N.; Oza, A.M.; Blank, S.V.; Cohen, R.; Adams, T.; Benites, A.; Gardiner, D.; Gregory, S.; Suzuki, M.; et al. The World Ovarian Cancer Coalition Every Woman Study: Identifying challenges and opportunities to improve survival and quality of life. Int. J. Gynecol. Cancer 2021, 31, 238–244. [Google Scholar] [CrossRef]
- Berek, J.S.; Kehoe, S.T.; Kumar, L.; Friedlander, M. FIGO CANCER REPORT 2018: Cancer of the Ovary, Fallopian Tube, and Peritoneum. Int. J. Gynecol. Obstet. 2018, 143, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Colombo, N.; Sessa, C.; Bois, A.D.; Ledermann, J.; McCluggage, W.G.; McNeish, I.; Morice, P.; Pignata, S.; Ray-Coquard, I.; Vergote, I.; et al. On behalf of the ESMO–ESGO Ovarian Cancer Consensus Conference Working Group. ESMO–ESGO consensus conference recommendations on ovarian cancer: Pathology and molecular biology, early and advanced stages, borderline tumours and recurrent disease. Int. J. Gynecol. Cancer 2019, 29, 728–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, J.A.; Raja, F.A.; Fotopoulou, C.; Gonzalez-Martin, A.; Colombo, N.; Sessa, C. Newly Diagnosed and Relapsed Epithelial Ovarian Carcinoma: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2013, 24, vi24–vi32. [Google Scholar] [CrossRef]
- Drapkin, R.; Von Horsten, H.H.; Lin, Y.; Mok, S.C.; Crum, C.P.; Welch, W.R.; Hecht, J.L. Human Epididymis Protein 4 (HE4) Is a Secreted Glycoprotein That Is Overexpressed by Serous and Endometrioid Ovarian Carcinomas. Cancer Res. 2005, 65, 2162–2169. [Google Scholar] [CrossRef] [Green Version]
- Heliström, I.; Raycraft, J.; Hayden-Ledbetter, M.; Ledbetter, J.A.; Schummer, M.; McIntosh, M.; Drescher, C.; Urban, N.; Hellström, K.E. The HE4 (WFDC2) Protein Is a Biomarker for Ovarian Carcinoma. Cancer Res. 2003, 63, 3695–3700. [Google Scholar]
- Qu, W.; Gao, Q.; Chen, H.; Tang, Z.; Zhu, X. HE4-Test of Urine and Body Fluids for Diagnosis of Gynecologic Cancer. Expert Rev. Mol. Diagn. 2017, 17, 239–244. [Google Scholar] [CrossRef]
- Scaletta, G.; Plotti, F.; Luvero, D.; Capriglione, S.; Montera, R.; Miranda, A.; Lopez, S.; Terranova, C.; De Cicco Nardone, C.; Angioli, R. The Role of Novel Biomarker HE4 in the Diagnosis, Prognosis and Follow-up of Ovarian Cancer: A Systematic Review. Expert Rev. Anticancer. Ther. 2017, 17, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Bian, L.I.H.; Chang, L.I.L.I.; Gao, X.I.N. Comparison of Serum Human Epididymis Protein 4 and Carbohydrate Antigen 125 as Markers in Ovarian Cancer: A Meta—Analysis. Mol. Clin. Oncol. 2014, 2, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, R.G.; McMeekin, D.S.; Brown, A.K.; DiSilvestro, P.; Miller, M.C.; Allard, W.J.; Gajewski, W.; Kurman, R.; Bast, R.C.; Skates, S.J. A Novel Multiple Marker Bioassay Utilizing HE4 and CA125 for the Prediction of Ovarian Cancer in Patients with a Pelvic Mass. Gynecol. Oncol. 2009, 112, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Gislefoss, R.E.; Langseth, H.; Bolstad, N.; Nustad, K.; Mørkrid, L. HE4 as an Early Detection Biomarker of Epithelial Ovarian Cancer: Investigations in Prediagnostic Specimens from the Janus Serumbank. Int. J. Gynecol. Cancer 2015, 25, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- O’Laughlin, D.J.; Strelow, B.; Fellows, N.; Kelsey, E.; Peters, S.; Stevens, J.; Tweedy, J. Addressing Anxiety and Fear during the Female Pelvic Examination. J. Prim. Care Community Health 2021, 12, 2150132721992195. [Google Scholar] [CrossRef] [PubMed]
- Nash, Z.; Menon, U. Ovarian Cancer Screening: Current Status and Future Directions. Best Pr. Res. Clin. Obstet. Gynaecol. 2020, 65, 32–45. [Google Scholar] [CrossRef]
- Grayson, K.; Gregory, E.; Khan, G.; Guinn, B.-A. Urine Biomarkers for the Early Detection of Ovarian Cancer—Are We There Yet? Biomark. Cancer 2019, 11, 1179299X19830977. [Google Scholar] [CrossRef]
- Harpole, M.; Davis, J.; Espina, V. Current State of the Art for Enhancing Urine Biomarker Discovery. Expert Rev. Proteom. 2016, 13, 609–626. [Google Scholar] [CrossRef] [Green Version]
- Jing, J.; Gao, Y. Urine Biomarkers in the Early Stages of Diseases: Current Status and Perspective. Discov. Med. 2018, 25, 57–65. [Google Scholar] [PubMed]
- Hellstrom, I.; Heagerty, P.J.; Swisher, E.M.; Liu, P.; Jaffar, J.; Agnew, K.; Hellström, K.E. Detection of the HE4 Protein in Urine as a Biomarker for Ovarian Neoplasms. Cancer Lett. 2010, 296, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Q.; Luo, G.; Yi, T.; Wang, Q.; Wang, D.; Zhang, G.; Jiang, X.; Guo, X. Diagnostic Value of Urinary-to-Serum Human Epididymis Protein 4 Ratio in Ovarian Cancer. Biomed. Rep. 2017, 7, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.B.; Yip, Y.Y.; Swisher, E.M.; Agnew, K.; Hellstrom, K.E.; Hellstrom, I. Detection of the HE4 Protein in Urine as a Biomarker for Ovarian Neoplasms: Clinical Correlates. Gynecol. Oncol. 2015, 137, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Macuks, R.; Baidekalna, I.; Donina, S. Urinary Concentrations of Human Epidydimis Secretory Protein 4 (HE4) in the Diagnosis of Ovarian Cancer: A Case-Control Study. Asian Pac. J. Cancer Prev. 2012, 13, 4695–4698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, M.; Deng, J.; Cheng, X.; Yan, Z.; Li, Q. Diagnostic Accuracy of Urine HE4 in Patients with Ovarian Cancer: A Meta-Analysis. Oncotarget 2017, 8, 9660–9671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciancio, D.R.; Vargas, M.R.; Thiel, W.H.; Bruno, M.A.; Giangrande, P.H.; Mestre, M.B. Aptamers as Diagnostic Tools in Cancer. Pharmaceuticals 2018, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Chen, J.; Wu, M.; Zhao, J.X. Aptamers: Active Targeting Ligands for Cancer Diagnosis and Therapy. Theranostics 2015, 5, 322–344. [Google Scholar] [CrossRef]
- Komarova, N.; Kuznetsov, A. Inside the Black Box: What Makes Selex Better? Molecules 2019, 24, 3598. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Gao, L.; Wang, J.; Wang, J. Application and Development of Aptamer in Cancer: From Clinical Diagnosis to Cancer Therapy. J. Cancer 2020, 11, 6902–6915. [Google Scholar] [CrossRef]
- Hanžek, A.; Siatka, C.; Duc, A.-C. High-Specificity Nucleic Acid Aptamers for Detection of Ovarian Cancer Protein Biomarkers: Application in Diagnostics. Aptamers 2021, 5, 7–14. [Google Scholar]
- Ang, A.; Ouellet, E.; Cheung, K.C.; Haynes, C. Highly Efficient and Reliable DNA Aptamer Selection Using the Partitioning Capabilities of Ddpcr: The Hi-Fi Selex Method. Methods Mol. Biol. 2018, 1768, 531–554. [Google Scholar] [CrossRef]
- Sarigul, N.; Korkmaz, F.; Kurultak, İ. A New Artificial Urine Protocol to Better Imitate Human Urine. Sci. Rep. 2019, 9, 20159. [Google Scholar] [CrossRef] [Green Version]
- Quang, N.N.; Miodek, A.; Cibiel, A.; Ducongé, F. Selection of Aptamers Against Whole Living Cells: From Cell-SELEX to Identification of Biomarkers. Methods Mol. Biol. 2017, 1575, 253–272. [Google Scholar] [CrossRef]
- Quang, N.N.; Bouvier, C.; Henriques, A.; Lelandais, B.; Ducongé, F. Time-Lapse Imaging of Molecular Evolution by High-Throughput Sequencing. Nucleic Acids Res. 2018, 46, 7480–7494. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar]
- Zuker, M. Mfold Web Server for Nucleic Acid Folding and Hybridization Prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Hu, J.; Kim, J.; Easley, C.J. Quantifying Aptamer-Protein Binding via Thermofluorimetric Analysis. Anal. Methods 2015, 7, 7358–7362. [Google Scholar] [CrossRef] [Green Version]
- Damase, T.R.; Allen, P.B. Idiosyncrasies of Thermofluorimetric Aptamer Binding Assays. Biotechniques 2019, 66, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong-Qian, L.; Radwan, R.; Polkowski, J.; Raju, S.; Falcone, K.; Simamora, R.; Kettly, T.; Moore, R.G.; Barnes, G. Abstract LB-451: Detection of Urinary HE4 as a Biomarker for Epithelial Ovarian Cancer. Cancer Res. 2011, 71, LB-451. [Google Scholar] [CrossRef]
- Pla, L.; Sancenón, F.; Martínez-bisbal, M.C.; Bañuls, C.; Estañ, N.; Botello-marabotto, M.; Aznar, E.; Sáez, G.; Santiago-felipe, S.; Martínez-máñez, R. A New 8-Oxo-7,8-2’deoxyguanosine Nanoporous Anodic Alumina Aptasensor for Colorectal Cancer Diagnosis in Blood and Urine. Nanoscale 2021, 13, 8648–8657. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cycle | DNA Source | DNA (nmol) | Protein (pmol) | Ratio |
|---|---|---|---|---|
| C1+ | library | 1.25 | 200 | 6.25:1 |
| C2+ | C1+ | 0.47 | 200 | 2.35:1 |
| C3+ | C2+ | 0.08 | 100 | 0.80:1 |
| C4+ | C3+ | 0.13 | 100 | 1.30:1 |
| C5+ | C4+ | 0.15 | 100 | 1.50:1 |
| C6+ | C5+ | 0.04 | 200 | 0.20:1 |
| C7+ | C6+ | 0.03 | 200 | 0.15:1 |
| C8+ | C7+ | 0.08 | 200 | 0.40:1 |
| C9+ | C8+ | 0.10 | 200 | 0.50:1 |
| C9− | C8+ | 0.10 | 200 | 0.50:1 |
| C10+ | C9+ | 0.10 | 100 | 1.00:1 |
| C10− | C9− | 0.10 | 100 | 1.00:1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanžek, A.; Ducongé, F.; Siatka, C.; Duc, A.-C.E. Identification and Characterization of Aptamers Targeting Ovarian Cancer Biomarker Human Epididymis Protein 4 for the Application in Urine. Cancers 2023, 15, 452. https://doi.org/10.3390/cancers15020452
Hanžek A, Ducongé F, Siatka C, Duc A-CE. Identification and Characterization of Aptamers Targeting Ovarian Cancer Biomarker Human Epididymis Protein 4 for the Application in Urine. Cancers. 2023; 15(2):452. https://doi.org/10.3390/cancers15020452
Chicago/Turabian StyleHanžek, Antonija, Frédéric Ducongé, Christian Siatka, and Anne-Cécile E. Duc. 2023. "Identification and Characterization of Aptamers Targeting Ovarian Cancer Biomarker Human Epididymis Protein 4 for the Application in Urine" Cancers 15, no. 2: 452. https://doi.org/10.3390/cancers15020452