
Matrix Metalloproteinase-1 (MMP1) Upregulation through Promoter Hypomethylation Enhances Tamoxifen Resistance in Breast Cancer

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Lentiviral Infection
2.2. Study Subjects
2.3. Quantitative Real-Time RT-PCR (qRT-PCR) and Methylation-Specific PCR (MSP)
2.4. Proliferation and Tamoxifen Sensitivity Assays
2.5. Western Blot Analysis
2.6. Apoptosis Assay
2.7. Xenograft Mouse Model
2.8. Immunohistochemistry (IHC) Staining
2.9. Data Mining and Statistical Analysis
3. Results
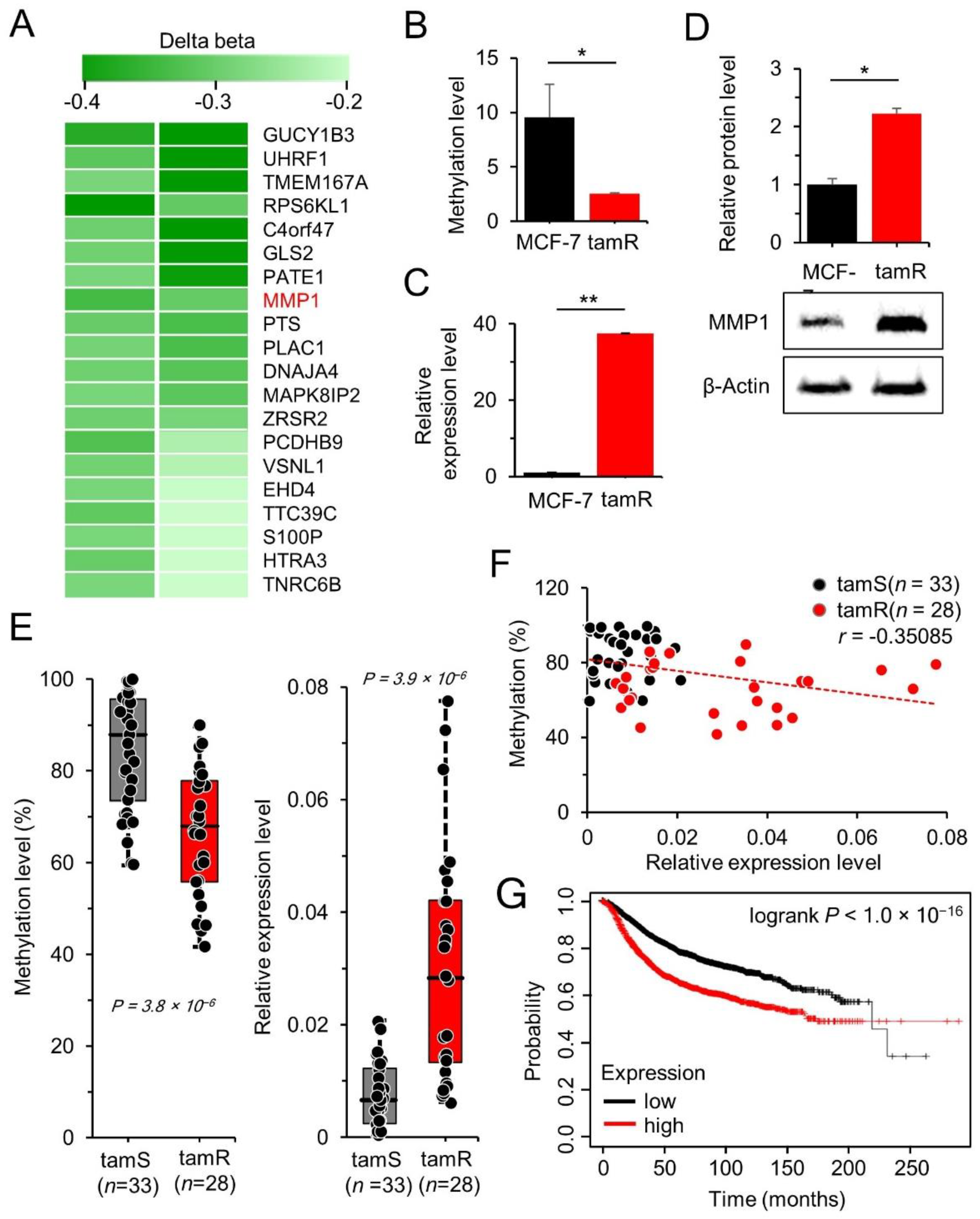
3.1. MMP1 Is Upregulated by DNA Hypomethylation in tamR Breast Cancer
3.2. MMP1 Stimulates MCF-7/tamR Cell Proliferation and Enhances Tam Resistance
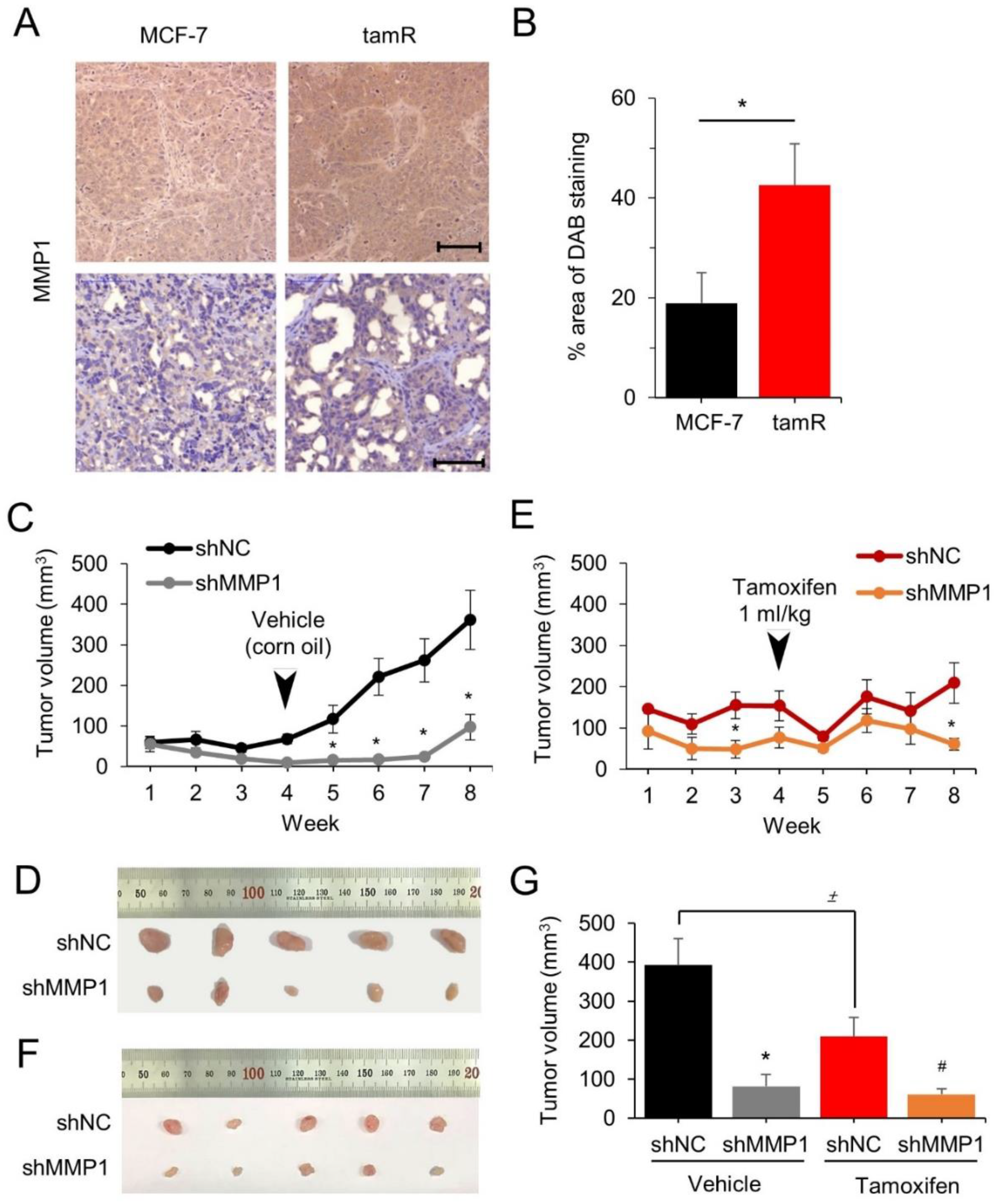
3.3. MMP1 Increases Tumor Growth of the MCF-7/tamR Cells in Xenografted Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, C.; Roldan, M.; Ruedas-Rama, M.J.; Orte, A.; Martin, M. Breast Cancer Cell Subtypes Display Different Metabolic Phenotypes That Correlate with Their Clinical Classification. Biology 2021, 10, 1267. [Google Scholar] [CrossRef] [PubMed]
- Crump, L.S.; Wyatt, G.L.; Rutherford, T.R.; Richer, J.K.; Porter, W.W.; Lyons, T.R. Hormonal Regulation of Semaphorin 7a in ER+ Breast Cancer Drives Therapeutic Resistance. Cancer Res. 2021, 81, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Radmacher, M.D.; Simon, R. Estimation of tamoxifen’s efficacy for preventing the formation and growth of breast tumors. J. Natl. Cancer Inst. 2000, 92, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.Y.; Kim, Y.C.; Oh, J.H.; Kim, M.H. HOXA5 confers tamoxifen resistance via the PI3K/AKT signaling pathway in ER-positive breast cancer. J. Cancer 2021, 12, 4626–4637. [Google Scholar] [CrossRef] [PubMed]
- Kastrati, I.; Joosten, S.E.P.; Semina, S.E.; Alejo, L.H.; Brovkovych, S.D.; Stender, J.D.; Horlings, H.M.; Kok, M.; Alarid, E.T.; Greene, G.L.; et al. The NF-κB Pathway Promotes Tamoxifen Tolerance and Disease Recurrence in Estrogen Receptor-Positive Breast Cancers. Mol. Cancer Res. 2020, 18, 1018–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieuwerts, A.M.; Inda, M.A.; Smid, M.; van Ooijen, H.; van de Stolpe, A.; Martens, J.W.M.; Verhaegh, W.F.J. ER and PI3K Pathway Activity in Primary ER Positive Breast Cancer Is Associated with Progression-Free Survival of Metastatic Patients under First-Line Tamoxifen. Cancers 2020, 12, 802. [Google Scholar] [CrossRef] [Green Version]
- Belachew, E.B.; Sewasew, D.T. Molecular Mechanisms of Endocrine Resistance in Estrogen-Positive Breast Cancer. Front. Endocrinol. 2021, 12, 599586. [Google Scholar] [CrossRef]
- Lee, E.-S.; Han, W.; Kim, M.K.; Kim, J.; Yoo, T.-k.; Lee, M.H.; Lee, K.H.; Kim, T.Y.; Moon, H.-G.; Im, S.-A.; et al. Factors associated with late recurrence after completion of 5-year adjuvant tamoxifen in estrogen receptor positive breast cancer. BMC Cancer 2016, 16, 430. [Google Scholar] [CrossRef] [Green Version]
- Viedma-Rodríguez, R.; Baiza-Gutman, L.; Salamanca-Gómez, F.; Diaz-Zaragoza, M.; Martínez-Hernández, G.; Ruiz Esparza-Garrido, R.; Velázquez-Flores, M.A.; Arenas-Aranda, D. Mechanisms associated with resistance to tamoxifen in estrogen receptor-positive breast cancer (review). Oncol. Rep. 2014, 32, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Machuca, T.N.; Hsin, M.K.; Ott, H.C.; Chen, M.; Hwang, D.M.; Cypel, M.; Waddell, T.K.; Keshavjee, S. Injury-specific ex vivo treatment of the donor lung: Pulmonary thrombolysis followed by successful lung transplantation. Am. J. Respir. Crit. Care Med. 2013, 188, 878–880. [Google Scholar] [CrossRef] [PubMed]
- Goetz, M.P.; Rae, J.M.; Suman, V.J.; Safgren, S.L.; Ames, M.M.; Visscher, D.W.; Reynolds, C.; Couch, F.J.; Lingle, W.L.; Flockhart, D.A.; et al. Pharmacogenetics of tamoxifen biotransformation is associated with clinical outcomes of efficacy and hot flashes. J. Clin. Oncol. 2005, 23, 9312–9318. [Google Scholar] [CrossRef] [PubMed]
- Cossetti, R.J.; Tyldesley, S.K.; Speers, C.H.; Zheng, Y.; Gelmon, K.A. Comparison of breast cancer recurrence and outcome patterns between patients treated from 1986 to 1992 and from 2004 to 2008. J. Clin. Oncol. 2015, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Lønning, P.E. The potency and clinical efficacy of aromatase inhibitors across the breast cancer continuum. Ann. Oncol. 2011, 22, 503–514. [Google Scholar] [CrossRef]
- Bayliss, J.; Hilger, A.; Vishnu, P.; Diehl, K.; El-Ashry, D. Reversal of the estrogen receptor negative phenotype in breast cancer and restoration of antiestrogen response. Clin. Cancer Res. 2007, 13, 7029–7036. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Zhang, P.; Deng, W.; Oesterreich, S.; Lu, Y.; Mills, G.B.; Lee, A.V. Insulin-like growth factor-I inhibits progesterone receptor expression in breast cancer cells via the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin pathway: Progesterone receptor as a potential indicator of growth factor activity in breast cancer. Mol. Endocrinol. 2003, 17, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, J.A.; El-Ashry, D. ER re-expression and re-sensitization to endocrine therapies in ER-negative breast cancers. J. Mammary Gland Biol. Neoplasia 2009, 14, 67–78. [Google Scholar] [CrossRef]
- Hamadneh, L.; Al-Lakkis, L.; Alhusban, A.A.; Tarawneh, S.; Abu-Irmaileh, B.; Albustanji, S.; Al-Bawab, A.Q. Changes in Lactate Production, Lactate Dehydrogenase Genes Expression and DNA Methylation in Response to Tamoxifen Resistance Development in MCF-7 Cell Line. Genes 2021, 12, 777. [Google Scholar] [CrossRef]
- Muluhngwi, P.; Klinge, C.M. Identification and Roles of miR-29b-1-3p and miR29a-3p-Regulated and Non-Regulated lncRNAs in Endocrine-Sensitive and Resistant Breast Cancer Cells. Cancers 2021, 13, 3530. [Google Scholar] [CrossRef]
- Yang, X.; Phillips, D.L.; Ferguson, A.T.; Nelson, W.G.; Herman, J.G.; Davidson, N.E. Synergistic activation of functional estrogen receptor (ER)-alpha by DNA methyltransferase and histone deacetylase inhibition in human ER-alpha-negative breast cancer cells. Cancer Res. 2001, 61, 7025–7029. [Google Scholar]
- Williams, K.E.; Anderton, D.L.; Lee, M.P.; Pentecost, B.T.; Arcaro, K.F. High-density array analysis of DNA methylation in Tamoxifen-resistant breast cancer cell lines. Epigenetics 2014, 9, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, A.; Zotenko, E.; Locke, W.J.; Korbie, D.; Millar, E.K.; Pidsley, R.; Stirzaker, C.; Graham, P.; Trau, M.; Musgrove, E.A.; et al. DNA methylation of oestrogen-regulated enhancers defines endocrine sensitivity in breast cancer. Nat. Commun. 2015, 6, 7758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachdeva, M.; Wu, H.; Ru, P.; Hwang, L.; Trieu, V.; Mo, Y.Y. MicroRNA-101-mediated Akt activation and estrogen-independent growth. Oncogene 2011, 30, 822–831. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Toyama, T.; Sugiura, H.; Fujii, Y.; Yamashita, H. miR-206 Expression is down-regulated in estrogen receptor alpha-positive human breast cancer. Cancer Res. 2008, 68, 5004–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Li, Q.; Liu, C.; Wang, C.; Li, Y. Overexpression miR-24-3p repressed Bim expression to confer tamoxifen resistance in breast cancer. J. Cell. Biochem. 2019, 120, 12966–12976. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jin, L.J.; Zhang, X.Y. Exosomal miRNA-205 promotes breast cancer chemoresistance and tumorigenesis through E2F1. Aging 2021, 13, 18498–18514. [Google Scholar] [CrossRef]
- Kim, S.S.; Lee, M.H.; Lee, M.O. Histone methyltransferases regulate the transcriptional expression of ERα and the proliferation of tamoxifen-resistant breast cancer cells. Breast Cancer Res. Treat. 2020, 180, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zheng, J.; Yu, J.; Wu, Y.; Guo, J.; Xu, Z.; Sun, X. Knockdown of MMP-1 inhibits the progression of colorectal cancer by suppressing the PI3K/Akt/c-myc signaling pathway and EMT. Oncol. Rep. 2020, 43, 1103–1112. [Google Scholar] [CrossRef]
- Jeong, D.; Ham, J.; Kim, H.W.; Kim, H.; Ji, H.W.; Yun, S.H.; Park, J.E.; Lee, K.S.; Jo, H.; Han, J.H.; et al. ELOVL2: A novel tumor suppressor attenuating tamoxifen resistance in breast cancer. Am. J. Cancer Res. 2021, 11, 2568–2589. [Google Scholar] [CrossRef]
- Lee, S.; Park, S.; Lee, H.; Jeong, D.; Ham, J.; Choi, E.H.; Kim, S.J. ChIP-seq analysis reveals alteration of H3K4 trimethylation occupancy in cancer-related genes by cold atmospheric plasma. Free Radic. Biol. Med. 2018, 126, 133–141. [Google Scholar] [CrossRef]
- Kim, S.J.; Kelly, W.K.; Fu, A.; Haines, K.; Hoffman, A.; Zheng, T.; Zhu, Y. Genome-wide methylation analysis identifies involvement of TNF-α mediated cancer pathways in prostate cancer. Cancer Lett. 2011, 302, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, H.; Jeong, D.; Ham, J.; Park, S.; Choi, E.H.; Kim, S.J. Cold atmospheric plasma restores tamoxifen sensitivity in resistant MCF-7 breast cancer cell. Free Radic. Biol. Med. 2017, 110, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.; Barraclough, R.; Vasieva, O. Gene Expression Meta-Analysis of Potential Metastatic Breast Cancer Markers. Curr. Mol. Med. 2017, 17, 200–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.M.; Lv, L.; Tang, Y.; Zhang, L.; Wang, L.F. MMP-1 is overexpressed in triple-negative breast cancer tissues and the knockdown of MMP-1 expression inhibits tumor cell malignant behaviors in vitro. Oncol. Lett. 2019, 17, 1732–1740. [Google Scholar] [CrossRef] [Green Version]
- Coezy, E.; Borgna, J.L.; Rochefort, H. Tamoxifen and metabolites in MCF7 cells: Correlation between binding to estrogen receptor and inhibition of cell growth. Cancer Res. 1982, 42, 317–323. [Google Scholar]
- Pardo, A.; Selman, M. MMP-1: The elder of the family. Int. J. Biochem. Cell Biol. 2005, 37, 283–288. [Google Scholar] [CrossRef]
- Brinckerhoff, C.E.; Rutter, J.L.; Benbow, U. Interstitial collagenases as markers of tumor progression. Clin. Cancer Res. 2000, 6, 4823–4830. [Google Scholar]
- Poplineau, M.; Dufer, J.; Antonicelli, F.; Trussardi-Regnier, A. Epigenetic regulation of proMMP-1 expression in the HT1080 human fibrosarcoma cell line. Int. J. Oncol. 2011, 38, 1713–1718. [Google Scholar] [CrossRef]
- Thaler, J.D.; Achari, Y.; Lu, T.; Shrive, N.G.; Hart, D.A. Estrogen receptor beta and truncated variants enhance the expression of transfected MMP-1 promoter constructs in response to specific mechanical loading. Biol. Sex Differ. 2014, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.H.; Park, Y.H.; Jun, H.J.; Kong, J.; Kim, J.H.; Kim, J.A.; Yun, J.; Sun, J.M.; Won, Y.W.; Lee, S.; et al. Matrix Metalloproteinase-1 Expression Can Be Upregulated through Mitogen-Activated Protein Kinase Pathway under the Influence of Human Epidermal Growth Factor Receptor 2 Synergized with Estrogen Receptor. Mol. Cancer Res. 2010, 8, 1037–1047. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Sowers, J.Y.; Houston, K.D. IGFBP-1 Expression Promotes Tamoxifen Resistance in Breast Cancer Cells via Erk Pathway Activation. Front. Endocrinol. 2020, 11, 233. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Mukasa, A.; Inda, M.M.; Zhang, J.; Chin, L.; Cavenee, W.; Furnari, F. Guanylate binding protein 1 is a novel effector of EGFR-driven invasion in glioblastoma. J. Exp. Med. 2011, 208, 2657–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqui, M.; Bohrer, L.R.; Brady, N.J.; Chuntova, P.; Kemp, S.E.; Wardwell, C.T.; Nelson, A.C.; Schwertfeger, K.L. Epiregulin contributes to breast tumorigenesis through regulating matrix metalloproteinase 1 and promoting cell survival. Mol. Cancer 2015, 14, 138. [Google Scholar] [CrossRef] [Green Version]
- Hanzawa, M.; Shindoh, M.; Higashino, F.; Yasuda, M.; Inoue, N.; Hida, K.; Ono, M.; Kohgo, T.; Nakamura, M.; Notani, K.; et al. Hepatocyte growth factor upregulates E1AF that induces oral squamous cell carcinoma cell invasion by activating matrix metalloproteinase genes. Carcinogenesis 2000, 21, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Hamadneh, L.; Bahader, M.; Abuarqoub, R.; AlWahsh, M.; Alhusban, A.; Hikmat, S. PI3K/AKT and MAPK1 molecular changes preceding matrix metallopeptidases overexpression during tamoxifen-resistance development are correlated to poor prognosis in breast cancer patients. Breast Cancer 2021, 28, 1358–1366. [Google Scholar] [CrossRef]
- Yang, P.; Li, J.; Peng, C.; Tan, Y.; Chen, R.; Peng, W.; Gu, Q.; Zhou, J.; Wang, L.; Tang, J.; et al. TCONS_00012883 promotes proliferation and metastasis via DDX3/YY1/MMP1/PI3K-AKT axis in colorectal cancer. Clin. Transl. Med. 2020, 10, e211. [Google Scholar] [CrossRef]
- Gao, Z.; Wu, J.; Wu, X.; Zheng, J.; Ou, Y. SRPX2 boosts pancreatic cancer chemoresistance by activating PI3K/AKT axis. Open Med. 2020, 15, 1072–1082. [Google Scholar] [CrossRef]
- Jang, M.; Park, R.; Park, Y.-I.; Park, Y.; Lee, J.I.; Namkoong, S.; Lee, E.-J.; Park, J. LNX1 Contributes to Cell Cycle Progression and Cisplatin Resistance. Cancers 2021, 13, 4066. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Accession No. | Description | Δ β-Value a | Fold Change b |
|---|---|---|---|---|
| GUCY1B3 | NM_000857 | guanylate cyclase 1 soluble subunit beta 1 | −0.47 | −2.24 |
| UHRF1 | NM_001048201 | ubiquitin-like with PHD and ring finger domains 1 | −0.42 | −2.37 |
| TMEM167A | NM_174909 | transmembrane protein 167A | −0.37 | −2.32 |
| RPS6KL1 | NM_031464 | ribosomal protein S6 kinase-like 1 | −0.36 | −2.64 |
| C4orf47 | NM_001114357 | chromosome 4 open reading frame 47 | −0.35 | −1.87 |
| GLS2 | NM_013267 | glutaminase 2 | −0.34 | −2.15 |
| PATE1 | NM_138294 | prostate and testis expressed 1 | −0.33 | −1.60 |
| MMP1 | NM_002421 | matrix metallopeptidase 1 | −0.32 | −1.96 |
| PTS | NM_000317 | 6-pyruvoyltetrahydropterin synthase | −0.31 | −3.70 |
| PLAC1 | NM_021796 | placenta enriched 1 | −0.30 | −2.10 |
| DNAJA4 | NM_018602 | DnaJ heat shock protein family (Hsp40) member A4 | −0.30 | −1.62 |
| MAPK8IP2 | NM_012324 | mitogen-activated protein kinase 8 interacting protein 2 | −0.29 | −1.87 |
| ZRSR2 | NM_005089 | zinc finger CCCH-type, RNA binding motif and serine/arginine rich 2 | −0.29 | −1.54 |
| PCDHB9 | NM_019119 | protocadherin beta 9 | −0.27 | −3.16 |
| VSNL1 | NM_003385 | visinin-like 1 | −0.25 | −1.66 |
| EHD4 | NM_139265 | EH domain containing 4 | −0.24 | −1.88 |
| TTC39C | NM_153211 | tetratricopeptide repeat domain 39C | −0.24 | −1.94 |
| S100P | NM_005980 | S100 calcium binding protein P | −0.23 | −1.54 |
| HTRA3 | NM_053044 | HtrA serine peptidase 3 | −0.22 | −1.61 |
| TNRC6B | NM_001024843 | trinucleotide repeat containing adaptor 6B | −0.22 | −2.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.W.; Park, J.E.; Baek, M.; Kim, H.; Ji, H.W.; Yun, S.H.; Jeong, D.; Ham, J.; Park, S.; Lu, X.; et al. Matrix Metalloproteinase-1 (MMP1) Upregulation through Promoter Hypomethylation Enhances Tamoxifen Resistance in Breast Cancer. Cancers 2022, 14, 1232. https://doi.org/10.3390/cancers14051232
Kim HW, Park JE, Baek M, Kim H, Ji HW, Yun SH, Jeong D, Ham J, Park S, Lu X, et al. Matrix Metalloproteinase-1 (MMP1) Upregulation through Promoter Hypomethylation Enhances Tamoxifen Resistance in Breast Cancer. Cancers. 2022; 14(5):1232. https://doi.org/10.3390/cancers14051232
Chicago/Turabian StyleKim, Hyeon Woo, Jae Eun Park, Minjae Baek, Heejoo Kim, Hwee Won Ji, Sung Hwan Yun, Dawoon Jeong, Juyeon Ham, Sungbin Park, Xinpei Lu, and et al. 2022. "Matrix Metalloproteinase-1 (MMP1) Upregulation through Promoter Hypomethylation Enhances Tamoxifen Resistance in Breast Cancer" Cancers 14, no. 5: 1232. https://doi.org/10.3390/cancers14051232