
Therapeutic Target Identification and Inhibitor Screening against Riboflavin Synthase of Colorectal Cancer Associated Fusobacterium nucleatum
 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methodology
2.1. Data Retrieval
2.2. Pan-Genome and Core Genome Analysis
2.3. Potential Drug Targets Identification
2.4. Structural Retrieval and Virtual Screening
2.5. ADMET Profiling of Shortlisted Drug Candidates
2.6. Dynamic Simulation Analysis
3. Results
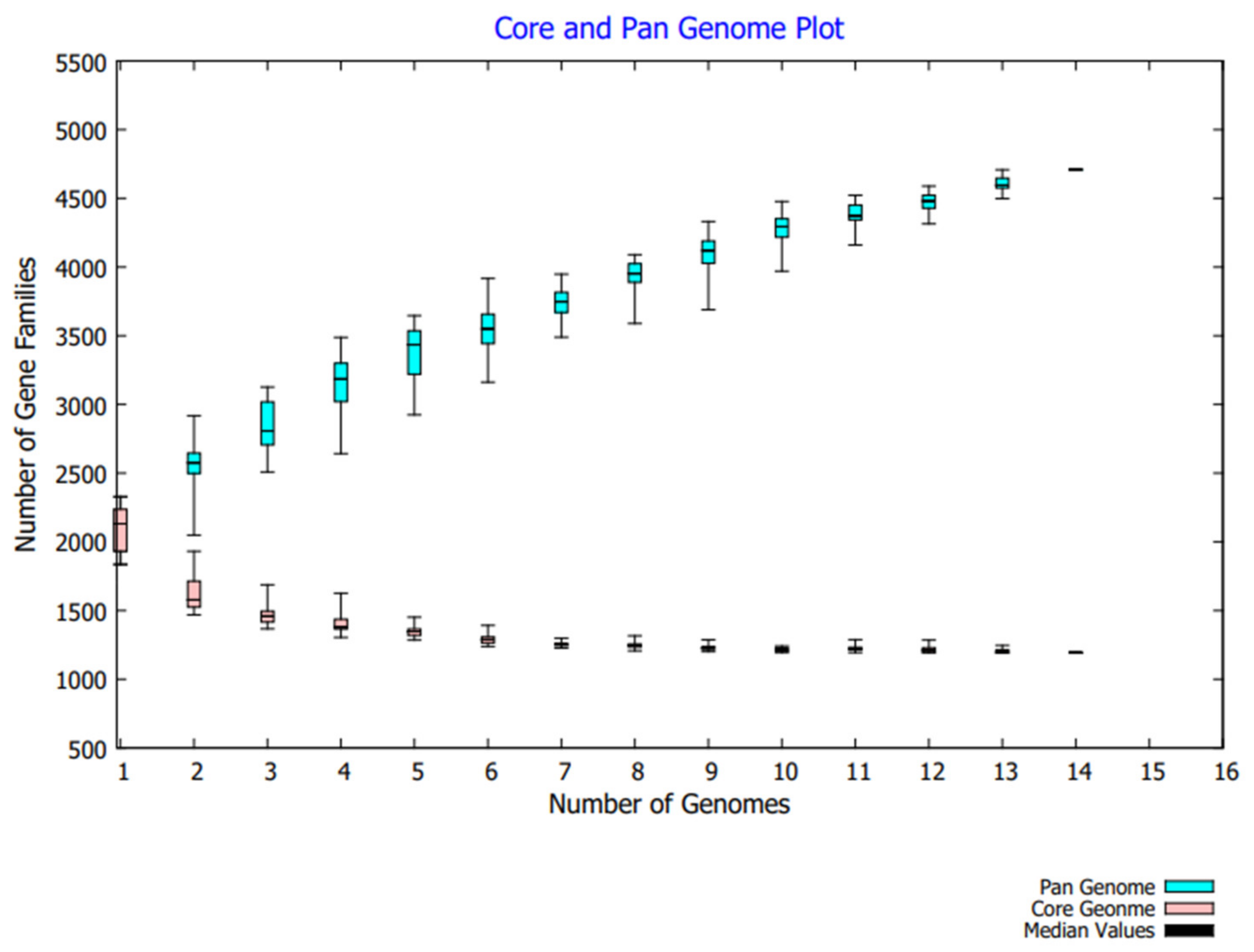
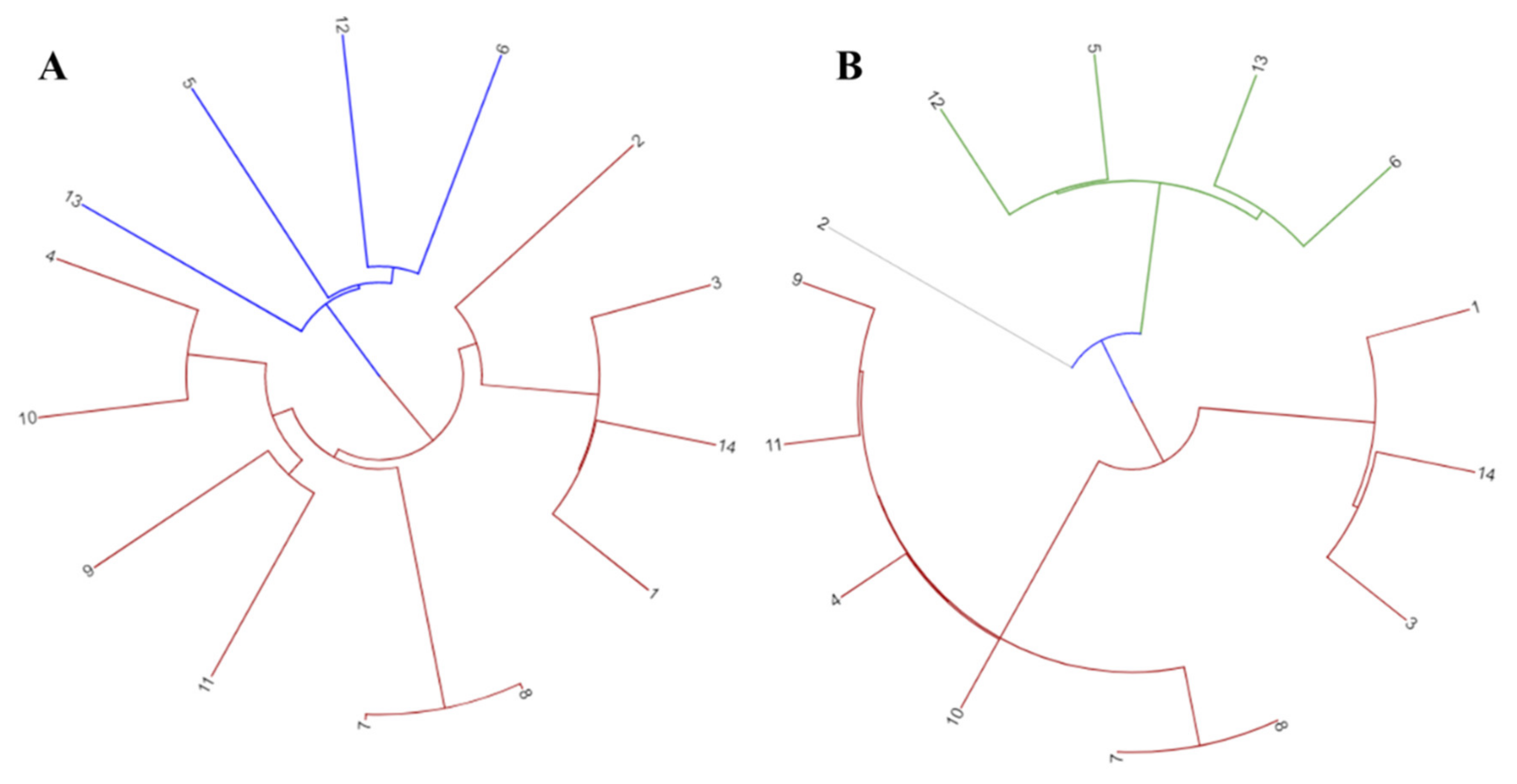
3.1. Pan-Genome Analysis
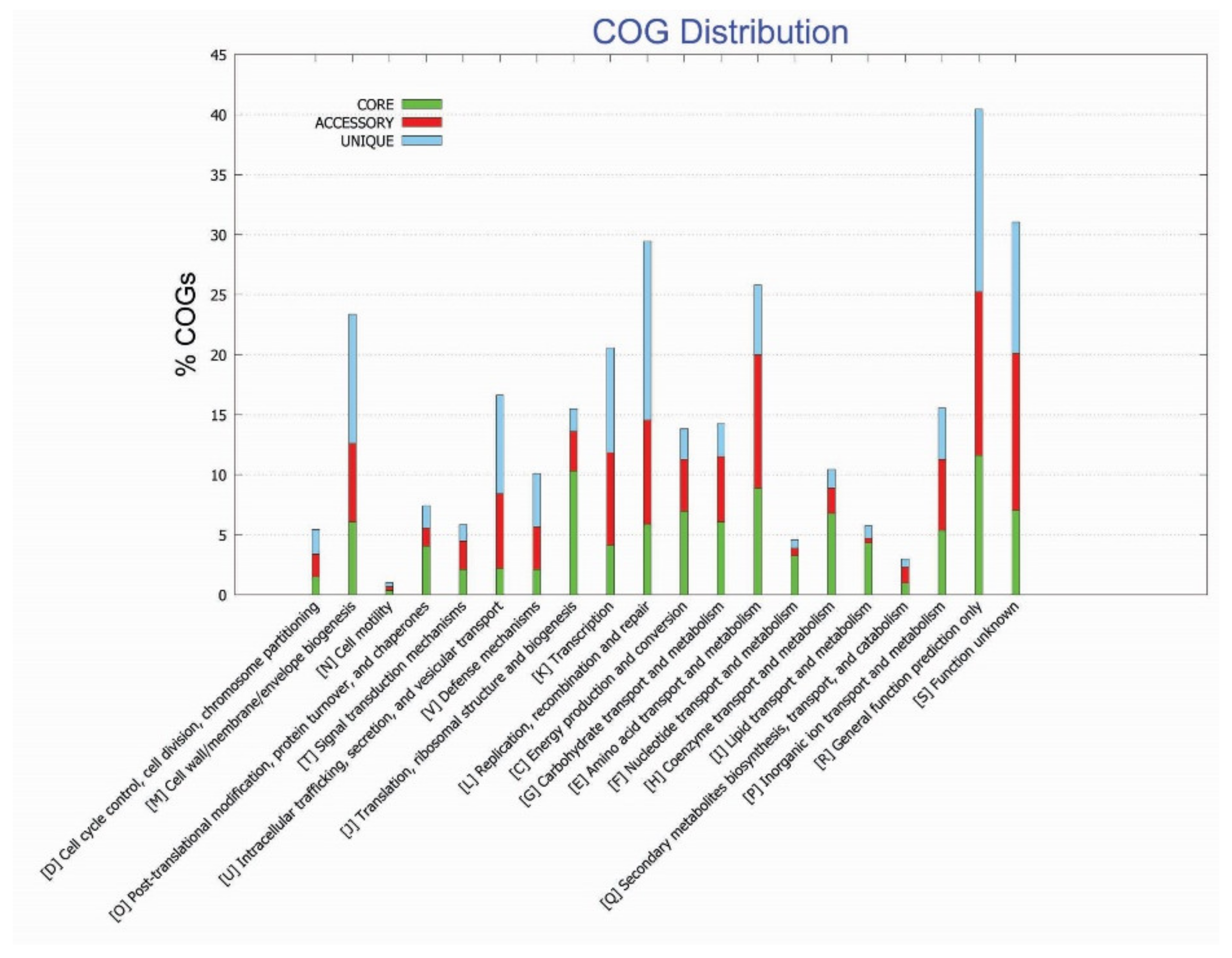
3.2. Functional Annotation Studies
3.3. Potential Drug Target Identification
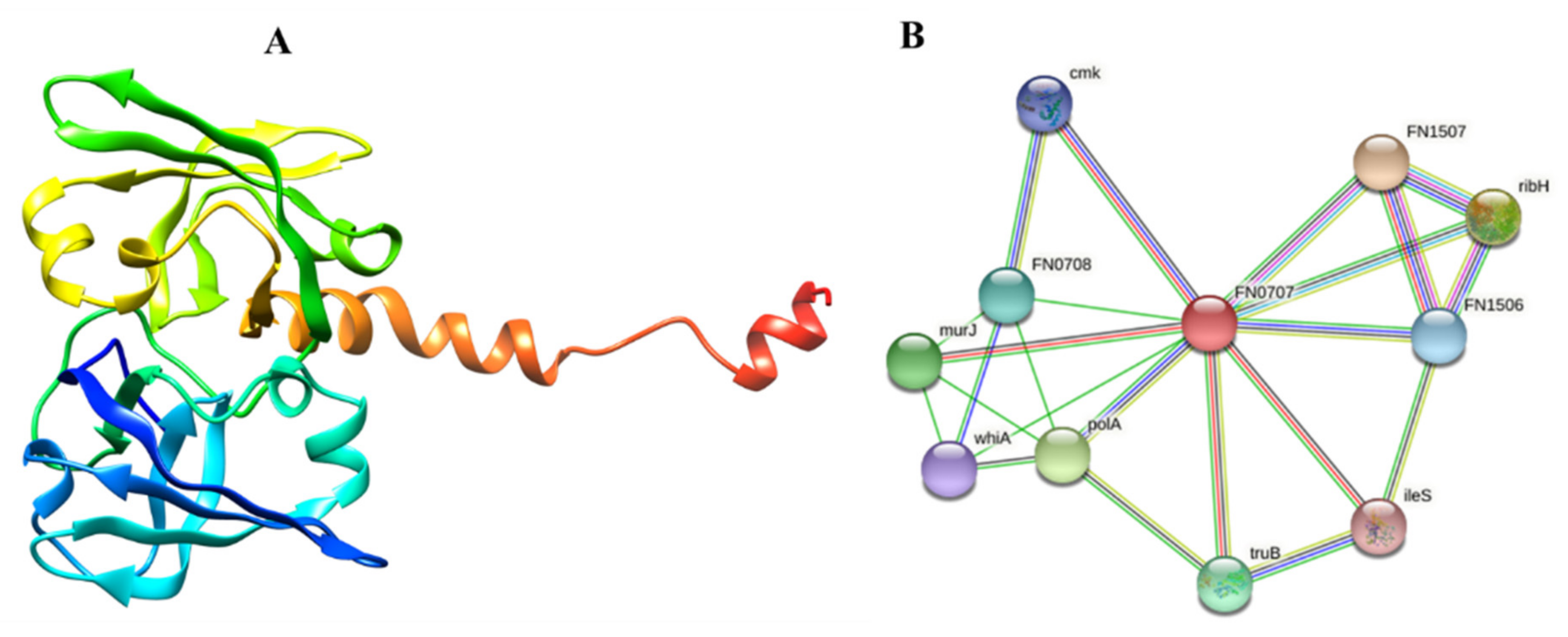
3.4. Significant and Novel Drug Target Prediction
3.5. Virtual Screening Studies
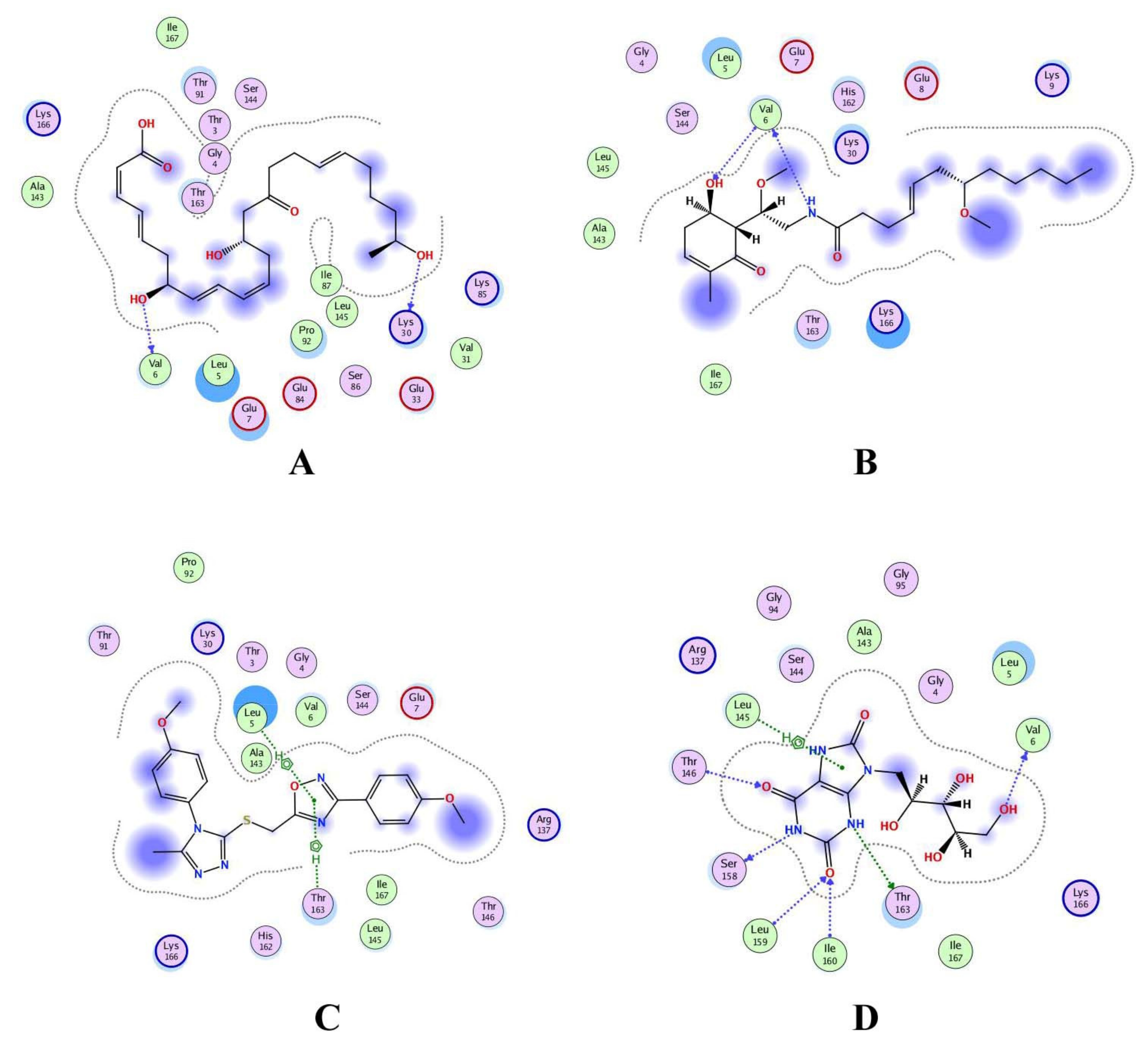
3.6. Interaction Analysis of Shortlisted Compounds
3.7. ADMET Profiling of Shortlisted Drug Candidates
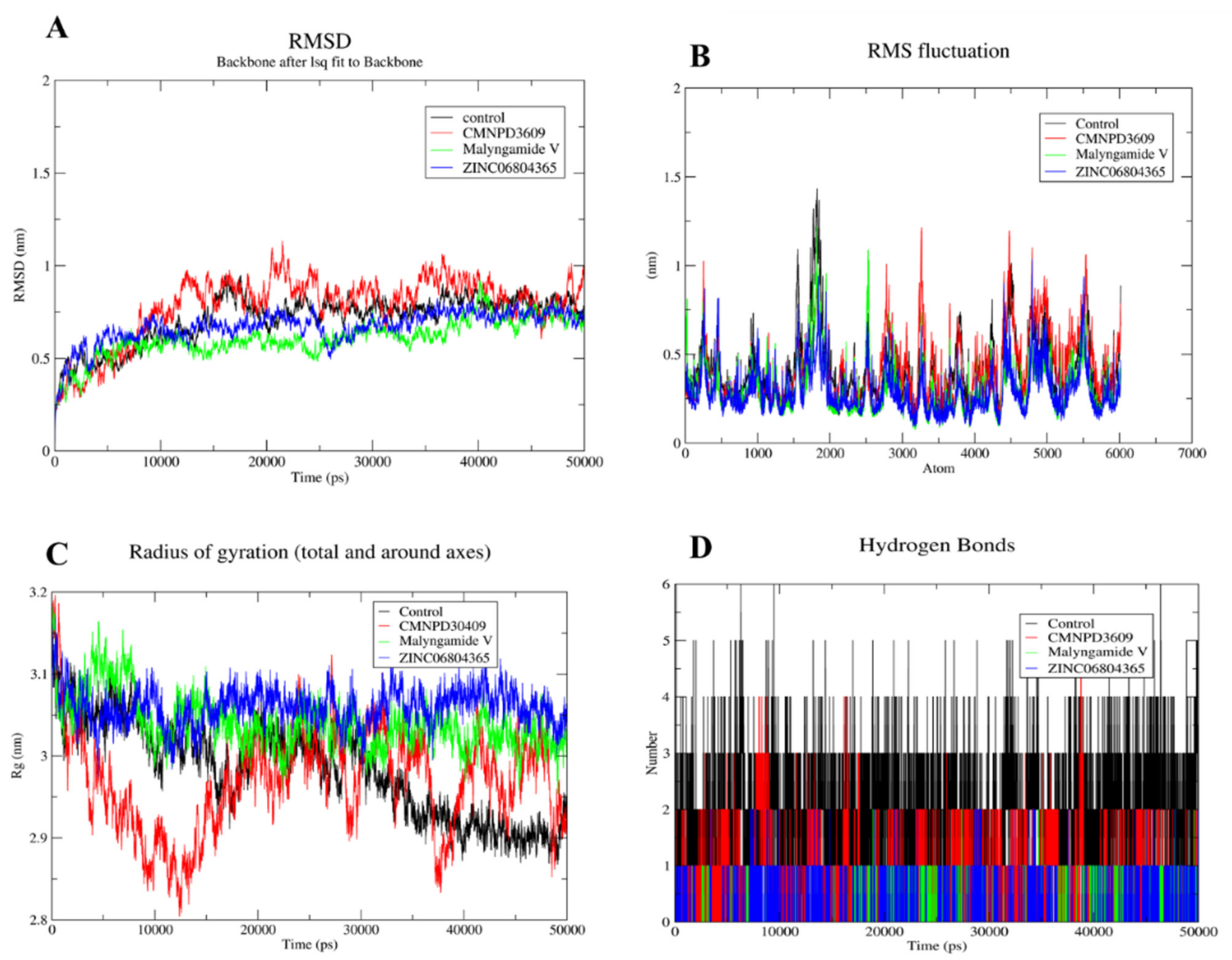
3.8. MD Simulation of Protein-Ligand Complex
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Abed, J.; Emgård, J.E.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A. Fap2 mediates Fusobacterium nucleatum colorectal adenocarcinoma enrichment by binding to tumor-expressed Gal-GalNAc. Cell Host Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellerba, F.; Serrano, D.; Harriet, J.; Pozzi, C.; Segata, N.; NabiNejad, A.; Piperni, E.; Gnagnarella, P.; Macis, D.; Aristarco, V.; et al. Colorectal cancer, Vitamin D and microbiota: A double-blind Phase II randomized trial (ColoViD) in colorectal cancer patients. Neoplasia 2022, 34, 100842. [Google Scholar] [CrossRef] [PubMed]
- Udayasuryan, B.; Ahmad, R.N.; Nguyen, T.T.; Umaña, A.; Monét Roberts, L.; Sobol, P.; Jones, S.D.; Munson, J.M.; Slade, D.J.; Verbridge, S.S. Fusobacterium nucleatum induces proliferation and migration in pancreatic cancer cells through host autocrine and paracrine signaling. Sci. Signal. 2022, 15, eabn4948. [Google Scholar] [CrossRef] [PubMed]
- Vogelstein, B.; Kinzler, K.W. The multistep nature of cancer. Trends Genet. 1993, 9, 138–141. [Google Scholar] [CrossRef]
- Abreu, M.T.; Peek, R.M., Jr. Gastrointestinal malignancy and the microbiome. Gastroenterology 2014, 146, 1534–1546.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zundler, S.; Mardin, C.; Bertz, S.; Vitali, F.; Strauß, R.; Fürst, J.; Neurath, M.F.; Strobel, D. Rectal Cancer Presenting with Absceding Infection Due to Fusobacterium nucleatum. Pathogens 2022, 11, 1113. [Google Scholar] [CrossRef]
- Ahn, H.; Min, K.; Lee, E.; Kim, H.; Kim, S.; Kim, Y.; Kim, G.; Cho, B.; Jeong, C.; Kim, Y.; et al. Whole-Transcriptome Sequencing Reveals Characteristics of Cancer Microbiome in Korean Patients with GI Tract Cancer: Fusobacterium nucleatum as a Therapeutic Target. Microorganisms 2022, 10, 1896. [Google Scholar] [CrossRef]
- Huh, J.W.; Kim, M.J.; Kim, J.; Lee, H.G.; Ryoo, S.B.; Ku, J.L.; Jeong, S.Y.; Park, K.J.; Kim, D.; Kim, J.F.; et al. Enterotypical Prevotella and three novel bacterial biomarkers in preoperative stool predict the clinical outcome of colorectal cancer. Microbiome 2022, 10, 203. [Google Scholar] [CrossRef]
- Keku, T.O.; McCoy, A.N.; Azcarate-Peril, A.M. Fusobacterium spp. and colorectal cancer: Cause or consequence? Trends Microbiol. 2013, 21, 506–508. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, C.; Ibrahim, Y.; Isaacson, B.; Yamin, R.; Abed, J.; Gamliel, M.; Enk, J.; Bar-On, Y.; Stanietsky-Kaynan, N.; Coppenhagen-Glazer, S. Binding of the Fap2 protein of Fusobacterium nucleatum to human inhibitory receptor TIGIT protects tumors from immune cell attack. Immunity 2015, 42, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M. Fusobacterium nucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ma, Q.; Guo, Y.; You, F. The Role of Fusobacterium nucleatum in Colorectal Cancer Cell Proliferation and Migration. Cancers 2022, 14, 5350. [Google Scholar] [CrossRef]
- Lo, C.-H.; Wu, D.-C.; Jao, S.-W.; Wu, C.-C.; Lin, C.-Y.; Chuang, C.-H.; Lin, Y.-B.; Chen, C.-H.; Chen, Y.-T.; Chen, J.-H.; et al. Enrichment of Prevotella intermedia in human colorectal cancer and its additive effects with Fusobacterium nucleatum on the malignant transformation of colorectal adenomas. J. Biomed. Sci. 2022, 29, 88. [Google Scholar] [CrossRef]
- Parhi, L.; Alon-Maimon, T.; Sol, A.; Nejman, D.; Shhadeh, A.; Fainsod-Levi, T.; Yajuk, O.; Isaacson, B.; Abed, J.; Maalouf, N. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Baik, J.E.; Li, L.; Shah, A.; Freedberg, D.E.; Jin, Z.; Wang, T.C.; Han, Y.W. Circulating IgA antibodies against Fusobacterium nucleatum amyloid adhesin FadA are a potential biomarker for colorectal neoplasia. Cancer Res. Commun. 2022, 2, 1497–1503. [Google Scholar] [CrossRef]
- Cavallucci, V.; Palucci, I.; Fidaleo, M.; Mercuri, A.; Masi, L.; Emoli, V.; Bianchetti, G.; Fiori, M.E.; Bachrach, G.; Scaldaferri, F.; et al. Proinflammatory and Cancer-Promoting Pathobiont Fusobacterium nucleatum Directly Targets Colorectal Cancer Stem Cells. Biomolecules 2022, 12, 1256. [Google Scholar] [CrossRef]
- Koveitypour, Z.; Panahi, F.; Vakilian, M.; Peymani, M.; Seyed Forootan, F.; Nasr Esfahani, M.H.; Ghaedi, K. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Slattery, M.L.; Lundgreen, A.; Wolff, R.K. MAP kinase genes and colon and rectal cancer. Carcinogenesis 2012, 33, 2398–2408. [Google Scholar] [CrossRef]
- Gao, Y.; Zou, T.; Xu, P.; Wang, Y.; Jiang, Y.; Chen, Y.X.; Chen, H.; Hong, J.; Fang, J.Y. Fusobacterium nucleatum stimulates cell proliferation and promotes PD-L1 expression via IFIT1-related signal in colorectal cancer. Neoplasia 2022, 35, 100850. [Google Scholar] [CrossRef] [PubMed]
- LaCourse, K.D.; Zepeda-Rivera, M.; Kempchinsky, A.G.; Baryiames, A.; Minot, S.S.; Johnston, C.D.; Bullman, S. The cancer chemotherapeutic 5-fluorouracil is a potent Fusobacterium nucleatum inhibitor and its activity is modified by intratumoral microbiota. Cell Rep. 2022, 41, 111625. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, P.; Zhao, Y.; Zhao, J.; Wu, X.; Zhang, R.; Cha, R.; Yao, Q.; Gao, Y. Nitroreductase-instructed supramolecular assemblies for microbiome regulation to enhance colorectal cancer treatments. Sci. Adv. 2022, 8, eadd2789. [Google Scholar] [CrossRef] [PubMed]
- Almatroudi, A. The incidence rate of colorectal cancer in Saudi Arabia: An observational descriptive epidemiological analysis. Int. J. Gen. Med. 2020, 13, 977. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.N.H.; Thu, T.N.H.; Nguyen, P.H.; Vo, C.N.; Doan, K.V.; Nguyen Ngoc Minh, C.; Nguyen, N.T.; Ta, V.N.D.; Vu, K.A.; Hua, T.D.; et al. Tumour microbiomes and Fusobacterium genomics in Vietnamese colorectal cancer patients. NPJ Biofilms Microbiomes 2022, 8, 87. [Google Scholar] [CrossRef]
- Tondi, D. Novel Targets and Mechanisms in Antimicrobial Drug Discovery. Antibiotics 2021, 10, 141. [Google Scholar] [CrossRef]
- Costa, D.; Iraola, G. Pathogenomics of emerging Campylobacter species. Clin. Microbiol. Rev. 2019, 32, e00072-18. [Google Scholar] [CrossRef]
- Basharat, Z.; Khan, K.; Jalal, K.; Ahmad, D.; Hayat, A.; Alotaibi, G.; Al Mouslem, A.; Alkhayl, F.F.A.; Almatroudi, A. An in silico hierarchal approach for drug candidate mining and validation of natural product inhibitors against pyrimidine biosynthesis enzyme in the antibiotic-resistant Shigella flexneri. Infect. Genet. Evol. 2022, 98, 105233. [Google Scholar] [CrossRef]
- Vasaikar, S.; Huang, C.; Wang, X.; Petyuk, V.A.; Savage, S.R.; Wen, B.; Dou, Y.; Zhang, Y.; Shi, Z.; Arshad, O.A. Proteogenomic analysis of human colon cancer reveals new therapeutic opportunities. Cell 2019, 177, 1035–1049.e19. [Google Scholar] [CrossRef] [Green Version]
- Caputo, A.; Fournier, P.-E.; Raoult, D. Genome and pan-genome analysis to classify emerging bacteria. Biol. Direct 2019, 14, 5. [Google Scholar] [CrossRef]
- Gordon, S.P.; Contreras-Moreira, B.; Levy, J.J.; Djamei, A.; Czedik-Eysenberg, A.; Tartaglio, V.S.; Session, A.; Martin, J.; Cartwright, A.; Katz, A. Gradual polyploid genome evolution revealed by pan-genomic analysis of Brachypodium hybridum and its diploid progenitors. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Allemailem, K.S. A Comprehensive Computer Aided Vaccine Design Approach to Propose a Multi-Epitopes Subunit Vaccine against Genus Klebsiella Using Pan-Genomics, Reverse Vaccinology, and Biophysical Techniques. Vaccines 2021, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Basharat, Z.; Jahanzaib, M.; Rahman, N. Therapeutic target identification via differential genome analysis of antibiotic resistant Shigella sonnei and inhibitor evaluation against a selected drug target. Infect. Genet. Evol. 2021, 94, 105004. [Google Scholar] [CrossRef] [PubMed]
- Basharat, Z.; Akhtar, U.; Khan, K.; Alotaibi, G.; Jalal, K.; Abbas, M.N.; Hayat, A.; Ahmad, D.; Hassan, S.S. Differential analysis of Orientia tsutsugamushi genomes for therapeutic target identification and possible intervention through natural product inhibitor screening. Comput. Biol. Med. 2022, 141, 105165. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Lin, Y.; Liu, T.; Lai, F.-L.; Zhang, C.-T.; Gao, F.; Zhang, R. DEG 15, an update of the Database of Essential Genes that includes built-in analysis tools. Nucleic Acids Res. 2021, 49, D677–D686. [Google Scholar] [CrossRef]
- Liu, S.; Wang, S.-X.; Liu, W.; Wang, C.; Zhang, F.-Z.; Ye, Y.-N.; Wu, C.-S.; Zheng, W.-X.; Rao, N.; Guo, F.-B. CEG 2.0: An updated database of clusters of essential genes including eukaryotic organisms. Database 2020, 2020, baaa112. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef]
- Queen, J.; Domingue, J.C.; White, J.R.; Stevens, C.; Udayasuryan, B.; Nguyen, T.T.D.; Wu, S.; Ding, H.; Fan, H.; McMann, M.; et al. Comparative Analysis of Colon Cancer-Derived Fusobacterium nucleatum Subspecies: Inflammation and Colon Tumorigenesis in Murine Models. mBio 2022, 13, e0299121. [Google Scholar] [CrossRef]
- Bi, D.; Zhu, Y.; Gao, Y.; Li, H.; Zhu, X.; Wei, R.; Xie, R.; Cai, C.; Wei, Q.; Qin, H. Profiling Fusobacterium infection at high taxonomic resolution reveals lineage-specific correlations in colorectal cancer. Nat. Commun. 2022, 13, 3336. [Google Scholar] [CrossRef]
- Bi, D.; Zhu, Y.; Gao, Y.; Li, H.; Zhu, X.; Wei, R.; Xie, R.; Wei, Q.; Qin, H. A newly developed PCR-based method revealed distinct Fusobacterium nucleatum subspecies infection patterns in colorectal cancer. Microb. Biotechnol. 2021, 14, 2176–2186. [Google Scholar] [CrossRef]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef] [Green Version]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basharat, Z.; Jahanzaib, M.; Yasmin, A.; Khan, I.A. Pan-genomics, drug candidate mining and ADMET profiling of natural product inhibitors screened against Yersinia pseudotuberculosis. Genomics 2021, 113, 238–244. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Vera Alvarez, R.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic Acids Res. 2021, 49, D274–D281. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef] [PubMed]
- Sarangi, A.N.; Lohani, M.; Aggarwal, R. Proteome mining for drug target identification in Listeria monocytogenes strain EGD-e and structure-based virtual screening of a candidate drug target penicillin binding protein 4. J. Microbiol. Methods 2015, 111, 9–18. [Google Scholar] [CrossRef]
- Chen, L.; Xiong, Z.; Sun, L.; Yang, J.; Jin, Q. VFDB 2012 update: Toward the genetic diversity and molecular evolution of bacterial virulence factors. Nucleic Acids Res. 2012, 40, D641–D645. [Google Scholar] [CrossRef]
- Kiefer, F.; Arnold, K.; Künzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Pires, D.E.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef] [PubMed]
- Pronk, S.; Páll, S.; Schulz, R.; Larsson, P.; Bjelkmar, P.; Apostolov, R.; Shirts, M.R.; Smith, J.C.; Kasson, P.M.; Van Der Spoel, D. GROMACS 4.5: A high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics 2013, 29, 845–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, K.; Khan, K.; Haleem, D.J.; Uddin, R. In silico study to identify new monoamine oxidase type a (MAO-A) selective inhibitors from natural source by virtual screening and molecular dynamics simulation. J. Mol. Struct. 2022, 1254, 132244. [Google Scholar] [CrossRef]
- Luo, J.; Wu, J. A new algorithm for essential proteins identification based on the integration of protein complex co-expression information and edge clustering coefficient. Int. J. Data Min. Bioinform. 2015, 12, 257–274. [Google Scholar] [CrossRef]
- Shanmugham, B.; Pan, A. Identification and characterization of potential therapeutic candidates in emerging human pathogen Mycobacterium abscessus: A novel hierarchical in silico approach. PLoS ONE 2013, 8, e59126. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Jalal, K.; Khan, A.; Al-Harrasi, A.; Uddin, R. Comparative Metabolic Pathways Analysis and Subtractive Genomics Profiling to Prioritize Potential Drug Targets Against Streptococcus pneumoniae. Front. Microbiol. 2021, 12, 796363. [Google Scholar] [CrossRef]
- Tutino, V.; Defrancesco, M.L.; Tolomeo, M.; De Nunzio, V.; Lorusso, D.; Paleni, D.; Caruso, M.G.; Notarnicola, M.; Barile, M. The expression of riboflavin transporters in human colorectal cancer. Anticancer Res. 2018, 38, 2659–2667. [Google Scholar]
- Mohd Yusof, H. Characterisation of Metabolic Changes in Colorectal Cancer Cells of Different Stages; Universiti Teknologi MARA: Shah Alam, Malaysia, 2020. [Google Scholar]
- Lu, Y.-T.; Gunathilake, M.; Lee, J.; Choi, I.J.; Kim, Y.-I.; Kim, J. Riboflavin intake, MTRR genetic polymorphism (rs1532268) and gastric cancer risk in a Korean population: A case–control study. Br. J. Nutr. 2022, 127, 1026–1033. [Google Scholar] [CrossRef]
- Serer, M.I.; Carrica, M.d.C.; Trappe, J.; López Romero, S.; Bonomi, H.R.; Klinke, S.; Cerutti, M.L.; Goldbaum, F.A. A high-throughput screening for inhibitors of riboflavin synthase identifies novel antimicrobial compounds to treat brucellosis. FEBS J. 2019, 286, 2522–2535. [Google Scholar] [CrossRef]
- Dietl, A.-M.; Meir, Z.; Shadkchan, Y.; Osherov, N.; Haas, H. Riboflavin and pantothenic acid biosynthesis are crucial for iron homeostasis and virulence in the pathogenic mold Aspergillus fumigatus. Virulence 2018, 9, 1036–1049. [Google Scholar] [CrossRef] [Green Version]
- Braun, P.; Gingras, A.C. History of protein–protein interactions: From egg-white to complex networks. Proteomics 2012, 12, 1478–1498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A. Protein Interaction Networks: Computational Analysis; Cambridge University Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Shahinozzaman, M.; Taira, N.; Ishii, T.; Halim, M.A.; Hossain, M.A.; Tawata, S. Anti-inflammatory, anti-diabetic, and anti-Alzheimer’s effects of prenylated flavonoids from Okinawa propolis: An investigation by experimental and computational studies. Molecules 2018, 23, 2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, S.Y.; Wu, V.W. A review on the special radiotherapy techniques of colorectal cancer. Front. Oncol. 2019, 9, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.-H.; Li, B.-B.; Wang, B.; Zhao, J.; Zhang, X.-Y.; Li, T.-T.; Li, W.-B.; Tang, D.; Qiu, M.-J.; Wang, X.-C. The role of Fusobacterium nucleatum in colorectal cancer: From carcinogenesis to clinical management. Chronic Dis. Transl. Med. 2019, 5, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Fukugaiti, M.H.; Ignacio, A.; Fernandes, M.R.; Ribeiro Júnior, U.; Nakano, V.; Avila-Campos, M.J. High occurrence of Fusobacterium nucleatum and Clostridium difficile in the intestinal microbiota of colorectal carcinoma patients. Braz. J. Microbiol. 2015, 46, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell 2017, 170, 548–563.e16. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: Implications for the microbial “pan-genome”. Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [Green Version]
- Jalal, K.; Khan, K.; Hayat, A.; Ahmad, D.; Alotaibi, G.; Uddin, R.; Mashraqi, M.M.; Alzamami, A.; Aurongzeb, M.; Basharat, Z. Mining therapeutic targets from the antibiotic-resistant Campylobacter coli and virtual screening of natural product inhibitors against its riboflavin synthase. Mol Divers 2022. [Google Scholar] [CrossRef]
- Long, Q.; Ji, L.; Wang, H.; Xie, J. Riboflavin biosynthetic and regulatory factors as potential novel anti-infective drug targets. Chem. Biol. Drug Des. 2010, 75, 339–347. [Google Scholar] [CrossRef]
- Aris, S.N.A.M.; Rahman, R.N.Z.R.A.; Ali, M.S.; Jonet, M.A.; Motomura, T.; Noor, N.D.M.; Shariff, F.M.; Hsu, K.; Leow, T.C. Unraveling the crystal structure of Leptospira kmetyi riboflavin synthase and computational analyses for potential development of new antibacterials. J. Mol. Struct. 2022, 165, 133420. [Google Scholar] [CrossRef]
- Lien, V.T.; Kristiansen, M.K.; Pettersen, S.; Haugen, M.H.; Olberg, D.E.; Waaler, J.; Klaveness, J. Towards dual inhibitors of the MET kinase and WNT signaling pathway; design, synthesis and biological evaluation. RSC Adv. 2019, 9, 37092–37100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, L.; Zhao, Z. Malyngamide F Possesses Anti-Inflammatory and Antinociceptive Activity in Rat Models of Inflammation. Pain Res. Manag. 2021, 2021, 4919391. [Google Scholar] [CrossRef]
- Cheng, Z.; Lou, L.; Liu, D.; Li, X.; Proksch, P.; Yin, S.; Lin, W. Versiquinazolines A–K, Fumiquinazoline-Type Alkaloids from the Gorgonian-Derived Fungus Aspergillus versicolor LZD-14-1. J. Nat. Prod. 2016, 79, 2941–2952. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No. | Name | Biosample Accession | Genome Size (Mbp) | Isolation Source | Coding DNA Sequences | Reference |
|---|---|---|---|---|---|---|
| 1. | F. nucleatum Fn146CP | SAMN20819806 | 2.082 | Tissue | 1949 | [38] |
| 2. | F. nucleatum Fn10-CTX3 | SAMN20819805 | 2.101 | Tissue | 2026 | [38] |
| 3. | F. nucleatum Fn3760T | SAMN20819808 | 2.299 | Tissue | 2234 | [38] |
| 4. | F. nucleatum Fn173CP | SAMN20819807 | 2.121 | Tissue | 2041 | [38] |
| 5. | F. nucleatum FnS043-1 | SAMN20819807 | 2.288 | Tissue | 2245 | [38] |
| 6. | F. nucleatum subsp. animalis strain THCT5A4 | SAMN20819807 | 2.491 | Gut | 2360 | [39] |
| 7. | F. nucleatum subsp. polymorphum strain THCT15E1 | SAMN18042967 | 2.526 | Gut | 2405 | [39] |
| 8. | F. nucleatum subsp. animalis strain THCT7A2 | SAMN18042965 | 2.515 | Gut | 2339 | [39,40] |
| 9. | F. nucleatum subsp. polymorphum strain THCT7E2 | SAMN18042966 | 2.547 | Gut | 2420 | [39] |
| 10. | F. nucleatum subsp. vincentii strain THCT14A3 | SAMN18042968 | 2.053 | Gut | 1903 | [39] |
| 11. | F. nucleatum subsp. animalis strain THCT6B3 | SAMN18042964 | 2.269 | Gut | 2116 | [39] |
| 12. | F. nucleatum subsp. animalis strain P2_CP | SAMN07448031 | 2.351 | Colorectal primary tumor | 2346 | [41] |
| 13. | F. nucleatum subsp. animalis strain P2_LM | SAMN07448032 | 2.346 | Liver metastasis | 2353 | - |
| 14. | F. nucleatum CC53 | SAMN02469329 | 2.070 | Colon adenocarcinoma | 1879 | [42] |
| S. No. | Ligand | Receptor | Interaction | Distance | E (kcal/mol) | S Value (kcal/mol) |
|---|---|---|---|---|---|---|
| 1 | Ribityl control | −6.40 | ||||
| N 2 | O SER 158 | H-donor | 2.87 | −5.9 | ||
| N 4 | OG1 THR 163 | H-donor | 3.06 | −2.0 | ||
| O 21 | O VAL 6 | H-donor | 2.91 | −1.4 | ||
| O 9 | CA LEU 159 | H-acceptor | 3.43 | −0.5 | ||
| O 9 | N ILE 160 | H-acceptor | 3.22 | −2.8 | ||
| O 10 | N THR 146 | H-acceptor | 2.95 | −2.5 | ||
| 5-ring | CA LEU 145 | pi-H | 4.49 | −1.0 | ||
| 2 | CMNPD3609 | −7.63 | ||||
| O 27 | O LYS 30 | H-donor | 3.18 | −0.8 | ||
| O 56 | O VAL 6 | H-donor | 2.94 | −1.8 | ||
| 3 | Malyngamide V | −7.03 | ||||
| N 13 | O VAL 6 | H-donor | 3.08 | −2.8 | ||
| O 7 | N VAL 6 | H-acceptor | 3.07 | −1.2 | ||
| 4 | ZINC06804365 | −7.01 | ||||
| 5-ring | CA LEU 5 | pi-H | 4.44 | −1.6 | ||
| 5-ring | CG2 THR 163 | pi-H | 4.28 | −0.5 | ||
| Compounds ID | CMNPD3609 | Malyngamide-V | ZINC6804365 |
| Compound Name | Isomacrolactic Acid | Malyngamide-V | 34M (PDB ID) |
| Structure |  |  |  |
| GI absorption | 100 | 92.819 | 98.36 |
| Caco2 | 1.317 | 1.076 | 1.242 |
| Water solubility | −0.102 | −4.386 | −3.651 |
| Skin permeability | −2.282 | −2.827 | −2.735 |
| BBB permeant | No | ||
| Lipinski | Yes | ||
| Binding Affinity (kcal/mol) | −7.63 | −7.03 | −5.9 |
| Radar |  |  |  |
| Ames | Yes | No | No |
| Max. tolerated dose (human) | 0.959 | 0.21 | 0.805 |
| Hepatotoxicity | No | No | Yes |
| Skin sensation | No | No | No |
| T.Pyriformis toxicity | −0.964 | 0.575 | 0.286 |
| Minnow toxicity | 4.806 | 0.929 | −2.353 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alturki, N.A.; Mashraqi, M.M.; Jalal, K.; Khan, K.; Basharat, Z.; Alzamami, A. Therapeutic Target Identification and Inhibitor Screening against Riboflavin Synthase of Colorectal Cancer Associated Fusobacterium nucleatum. Cancers 2022, 14, 6260. https://doi.org/10.3390/cancers14246260
Alturki NA, Mashraqi MM, Jalal K, Khan K, Basharat Z, Alzamami A. Therapeutic Target Identification and Inhibitor Screening against Riboflavin Synthase of Colorectal Cancer Associated Fusobacterium nucleatum. Cancers. 2022; 14(24):6260. https://doi.org/10.3390/cancers14246260
Chicago/Turabian StyleAlturki, Norah A., Mutaib M. Mashraqi, Khurshid Jalal, Kanwal Khan, Zarrin Basharat, and Ahmad Alzamami. 2022. "Therapeutic Target Identification and Inhibitor Screening against Riboflavin Synthase of Colorectal Cancer Associated Fusobacterium nucleatum" Cancers 14, no. 24: 6260. https://doi.org/10.3390/cancers14246260