
Far-Red Fluorescent Murine Glioma Model for Accurate Assessment of Brain Tumor Progression

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Cell Culture and Transfection
2.2. Mice and Ethics Statement
2.3. Surgical Procedures and Orthotopic Transplantation of Glioma GL261-Kat Cells In Vivo
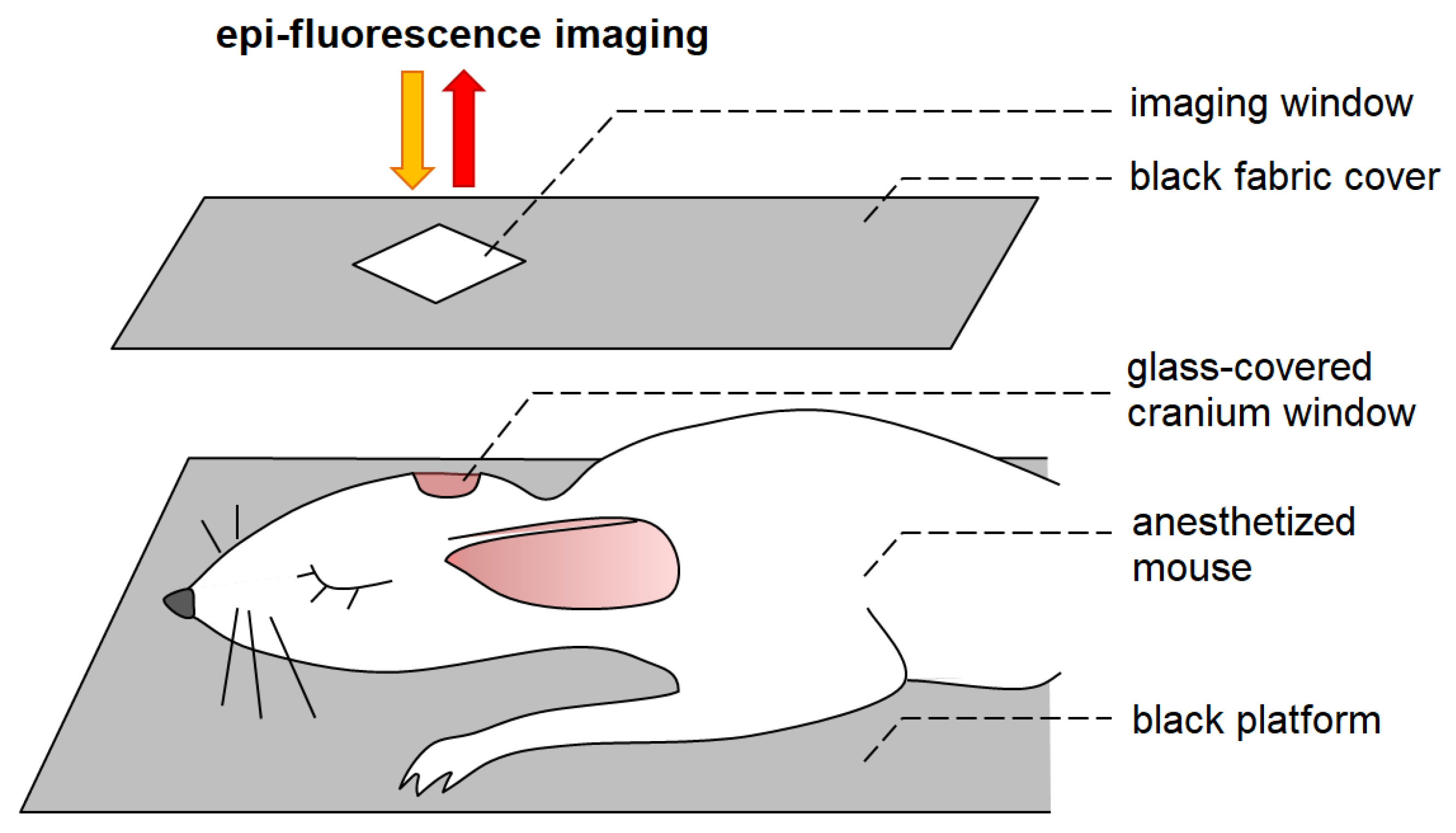
2.4. Epi-Fluorescence Imaging In Vivo
2.5. Magnetic Resonance Imaging
2.6. Neurological Status Determination
2.7. Open Field Test
2.8. Passive Avoidance Test
2.9. Histological Analysis
2.10. Statistical Analysis
3. Results
3.1. Creation of the Stable Fluorescent Murine Glioma GL261-Kat Cell Line
3.2. Morphological Alterations of Brain Tissue in Orthotopic Mouse Glioma Model
3.3. Assessment of Behavioral and Cognitive Reactions of Mice in the Orthotopic Glioma GL261-Kat Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Śledzińska, P.; Bebyn, M.G.; Furtak, J.; Kowalewski, J.; Lewandowska, M.A. Prognostic and Predictive Biomarkers in Gliomas. Int. J. Mol. Sci. 2021, 22, 10373. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.G.; Fine, H.A. Diffuse Glioma Heterogeneity and Its Therapeutic Implications. Cancer Discov. 2021, 11, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Ferris, S.P.; Hofmann, J.W.; Solomon, D.A.; Perry, A. Characterization of gliomas: From morphology to molecules. Virchows Arch. 2017, 471, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2014–2018. Neuro Oncol. 2021, 23, iii1–iii105. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Cote, D.J.; Ascha, M.; Kruchko, C.; Barnholtz-Sloan, J.S. Adult Glioma Incidence and Survival by Race or Ethnicity in the United States From 2000 to 2014. JAMA Oncol. 2018, 4, 1254–1262. [Google Scholar] [CrossRef] [Green Version]
- Tykocki, T.; Eltayeb, M. Ten-year survival in glioblastoma. A systematic review. J. Clin. Neurosci. 2018, 54, 7–13. [Google Scholar] [CrossRef]
- Barnholtz-Sloan, J.S.; Ostrom, Q.T.; Cote, D. Epidemiology of Brain Tumors. Neurol. Clin. 2018, 36, 395–419. [Google Scholar] [CrossRef]
- Hicks, W.H.; Bird, C.E.; Traylor, J.I.; Shi, D.D.; El Ahmadieh, T.Y.; Richardson, T.E.; McBrayer, S.K.; Abdullah, K.G. Contemporary Mouse Models in Glioma Research. Cells 2021, 10, 712. [Google Scholar] [CrossRef]
- Lenting, K.; Verhaak, R.; Ter Laan, M.; Wesseling, P.; Leenders, W. Glioma: Experimental models and reality. Acta Neuropathol. 2017, 133, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Holland, E.C. Glioma models. Biochim. Biophys. Acta 2001, 1551, M19–M27. [Google Scholar] [CrossRef]
- Torres-Garcia, L.; JM, P.D.; Brandi, E.; Haikal, C.; Mudannayake, J.M.; Brás, I.C.; Gerhardt, E.; Li, W.; Svanbergsson, A.; Outeiro, T.F.; et al. Monitoring the interactions between alpha-synuclein and Tau in vitro and in vivo using bimolecular fluorescence complementation. Sci. Rep. 2022, 12, 2987. [Google Scholar] [CrossRef] [PubMed]
- Peskova, N.N.; Brilkina, A.A.; Gorokhova, A.A.; Shilyagina, N.Y.; Kutova, O.M.; Nerush, A.S.; Orlova, A.G.; Klapshina, L.G.; Vodeneev, V.V.; Balalaeva, I.V. The localization of the photosensitizer determines the dynamics of the secondary production of hydrogen peroxide in cell cytoplasm and mitochondria. J. Photochem. Photobiol. B 2021, 219, 6. [Google Scholar] [CrossRef]
- Sokolova, E.; Kutova, O.; Grishina, A.; Pospelov, A.; Guryev, E.; Schulga, A.; Deyev, S.; Balalaeva, I. Penetration Efficiency of Antitumor Agents in Ovarian Cancer Spheroids: The Case of Recombinant Targeted Toxin DARPin-LoPE and the Chemotherapy Drug, Doxorubicin. Pharmaceutics 2019, 11, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilkina, A.A.; Peskova, N.N.; Dudenkova, V.V.; Gorokhova, A.A.; Sokolova, E.A.; Balalaeva, I.V. Monitoring of hydrogen peroxide production under photodynamic treatment using protein sensor HyPer. J. Photochem. Photobiol. B 2018, 178, 296–301. [Google Scholar] [CrossRef]
- Hodgson, L.; Qiu, W.; Dong, C.; Henderson, A.J. Use of green fluorescent protein-conjugated beta-actin as a novel molecular marker for in vitro tumor cell chemotaxis assay. Biotechnol. Prog. 2000, 16, 1106–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, M.A.; Lwin, T.M.; Amirfakhri, S.; Nishino, H.; Hoffman, R.M.; Yazaki, P.J.; Bouvet, M. The Use of Fluorescent Anti-CEA Antibodies to Label, Resect and Treat Cancers: A Review. Biomolecules 2021, 11, 1819. [Google Scholar] [CrossRef]
- Warnders, F.J.; Lub-de Hooge, M.N.; de Vries, E.G.E.; Kosterink, J.G.W. Influence of protein properties and protein modification on biodistribution and tumor uptake of anticancer antibodies, antibody derivatives, and non-Ig scaffolds. Med. Res. Rev. 2018, 38, 1837–1873. [Google Scholar] [CrossRef]
- Hoffman, R.M. Use of fluorescent proteins and color-coded imaging to visualize cancer cells with different genetic properties. Cancer Metastasis Rev. 2016, 35, 5–19. [Google Scholar] [CrossRef]
- Kantelhardt, S.R.; Kalasauskas, D.; König, K.; Kim, E.; Weinigel, M.; Uchugonova, A.; Giese, A. In vivo multiphoton tomography and fluorescence lifetime imaging of human brain tumor tissue. J. Neurooncol. 2016, 127, 473–482. [Google Scholar] [CrossRef]
- Bouvet, M.; Hoffman, R.M. Tumor imaging technologies in mouse models. Methods Mol. Biol. 2015, 1267, 321–348. [Google Scholar] [PubMed]
- Zdobnova, T.; Sokolova, E.; Stremovskiy, O.; Karpenko, D.; Telford, W.; Turchin, I.; Balalaeva, I.; Deyev, S. A novel far-red fluorescent xenograft model of ovarian carcinoma for preclinical evaluation of HER2-targeted immunotoxins. Oncotarget 2015, 6, 30919–30928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, R.M. Fluorescent proteins as visible in vivo sensors. Prog. Mol. Biol. Transl. Sci. 2013, 113, 389–402. [Google Scholar] [PubMed]
- Turchin, I.; Plehanov, V.; Orlova, A.; Kamenskiy, V.; Kleshnin, M.; Shirmanova, M.; Shakhova, N.; Balalaeva, I.; Savitskiy, A. Fluorescence diffuse tomography of small animals with DsRed2 fluorescent protein. Laser Phys. 2006, 16, 741–746. [Google Scholar] [CrossRef]
- Christensen, J.; Vonwil, D.; Shastri, V.P. Non-Invasive In Vivo Imaging and Quantification of Tumor Growth and Metastasis in Rats Using Cells Expressing Far-Red Fluorescence Protein. PLoS ONE 2015, 10, e0132725. [Google Scholar] [CrossRef] [PubMed]
- Fei-Peng, Z.; Guo-Tao, C.; Shou-Ju, W.; Ying, L.; Yu-Xia, T.; Ying, T.; Jian-Dong, W.; Chun-Yan, W.; Xin, W.; Jing, S. Dual-modality imaging probes with high magnetic relaxivity and near-infrared fluorescence based highly aminated mesoporous silica nanoparticles. J. Nanomater. 2016, 2016, 6502127. [Google Scholar] [CrossRef] [Green Version]
- Harrison, V.S.; Carney, C.E.; MacRenaris, K.W.; Waters, E.A.; Meade, T.J. Multimeric Near IR-MR Contrast Agent for Multimodal In Vivo Imaging. J. Am. Chem. Soc. 2015, 137, 9108–9116. [Google Scholar] [CrossRef]
- Winnard, P.T., Jr.; Kluth, J.B.; Raman, V. Noninvasive optical tracking of red fluorescent protein-expressing cancer cells in a model of metastatic breast cancer. Neoplasia 2006, 8, 796–806. [Google Scholar] [CrossRef] [Green Version]
- Biasibetti, E.; Valazza, A.; Capucchio, M.T.; Annovazzi, L.; Battaglia, L.; Chirio, D.; Gallarate, M.; Mellai, M.; Muntoni, E.; Peira, E.; et al. Comparison of Allogeneic and Syngeneic Rat Glioma Models by Using MRI and Histopathologic Evaluation. Comp. Med. 2017, 67, 147–156. [Google Scholar]
- Deshpande, R.; Babu, P. Animal Model for Glioma: A Brief Overview. Arch. Med. Biotechnol. 2018, 4, 2–20. [Google Scholar]
- Novozhilova, M.; Mishchenko, T.; Kondakova, E.; Lavrova, T.; Gavrish, M.; Aferova, S.; Franceschi, C.; Vedunova, M. Features of age-related response to sleep deprivation: In vivo experimental studies. Aging 2021, 13, 19108–19126. [Google Scholar] [CrossRef] [PubMed]
- Prickaerts, J.; Fahrig, T.; Blokland, A. Cognitive performance and biochemical markers in septum, hippocampus and striatum of rats after an i.c.v. injection of streptozotocin: A correlation analysis. Behav. Brain Res. 1999, 102, 73–88. [Google Scholar] [CrossRef]
- Beni-Adani, L.; Gozes, I.; Cohen, Y.; Assaf, Y.; Steingart, R.A.; Brenneman, D.E.; Eizenberg, O.; Trembolver, V.; Shohami, E. A peptide derived from activity-dependent neuroprotective protein (ADNP) ameliorates injury response in closed head injury in mice. J. Pharmacol. Exp. Ther. 2001, 296, 57–63. [Google Scholar]
- Szatmári, T.; Lumniczky, K.; Désaknai, S.; Trajcevski, S.; Hídvégi, E.J.; Hamada, H.; Sáfrány, G. Detailed characterization of the mouse glioma 261 tumor model for experimental glioblastoma therapy. Cancer Sci. 2006, 97, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Shcherbo, D.; Merzlyak, E.M.; Chepurnykh, T.V.; Fradkov, A.F.; Ermakova, G.V.; Solovieva, E.A.; Lukyanov, K.A.; Bogdanova, E.A.; Zaraisky, A.G.; Lukyanov, S.; et al. Bright far-red fluorescent protein for whole-body imaging. Nat. Methods 2007, 4, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Luther, D.C.; Jeon, T.; Goswami, R.; Nagaraj, H.; Kim, D.; Lee, Y.W.; Rotello, V.M. Protein Delivery: If Your GFP (or Other Small Protein) Is in the Cytosol, It Will Also Be in the Nucleus. Bioconjug. Chem. 2021, 32, 891–896. [Google Scholar] [CrossRef]
- Ausman, J.I.; Shapiro, W.R.; Rall, D.P. Studies on the chemotherapy of experimental brain tumors: Development of an experimental model. Cancer Res. 1970, 30, 2394–2400. [Google Scholar]
- Vince, G.H.; Bendszus, M.; Schweitzer, T.; Goldbrunner, R.H.; Hildebrandt, S.; Tilgner, J.; Klein, R.; Solymosi, L.; Christian Tonn, J.; Roosen, K. Spontaneous regression of experimental gliomas--an immunohistochemical and MRI study of the C6 glioma spheroid implantation model. Exp. Neurol. 2004, 190, 478–485. [Google Scholar] [CrossRef]
- Sanchez, V.E.; Lynes, J.P.; Walbridge, S.; Wang, X.; Edwards, N.A.; Nwankwo, A.K.; Sur, H.P.; Dominah, G.A.; Obungu, A.; Adamstein, N.; et al. GL261 luciferase-expressing cells elicit an anti-tumor immune response: An evaluation of murine glioma models. Sci. Rep. 2020, 10, 11003. [Google Scholar] [CrossRef]
- Abdelwahab, M.G.; Sankar, T.; Preul, M.C.; Scheck, A.C. Intracranial implantation with subsequent 3D in vivo bioluminescent imaging of murine gliomas. J. Vis. Exp. 2011, 6, e3403. [Google Scholar] [CrossRef] [Green Version]
- Maes, W.; Van Gool, S.W. Experimental immunotherapy for malignant glioma: Lessons from two decades of research in the GL261 model. Cancer Immunol. Immunother. 2011, 60, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Villamañan, L.; Martínez-Escardó, L.; Arús, C.; Yuste, V.J.; Candiota, A.P. Successful Partnerships: Exploring the Potential of Immunogenic Signals Triggered by TMZ, CX-4945, and Combined Treatment in GL261 Glioblastoma Cells. Int. J. Mol. Sci. 2021, 22, 3453. [Google Scholar] [CrossRef] [PubMed]
- Alzeibak, R.; Mishchenko, T.A.; Shilyagina, N.Y.; Balalaeva, I.V.; Vedunova, M.V.; Krysko, D.V. Targeting immunogenic cancer cell death by photodynamic therapy: Past, present and future. J. Immunother. Cancer 2021, 9, e001926. [Google Scholar] [CrossRef]
- Turubanova, V.D.; Balalaeva, I.V.; Mishchenko, T.A.; Catanzaro, E.; Alzeibak, R.; Peskova, N.N.; Efimova, I.; Bachert, C.; Mitroshina, E.V.; Krysko, O.; et al. Immunogenic cell death induced by a new photodynamic therapy based on photosens and photodithazine. J. Immunother. Cancer 2019, 7, 019–0826. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.D.; Vandenberk, L.; Koks, C.; Verschuere, T.; Boon, L.; Van Gool, S.W.; Agostinis, P. Dendritic cell vaccines based on immunogenic cell death elicit danger signals and T cell-driven rejection of high-grade glioma. Sci. Transl. Med. 2016, 8, 328ra27. [Google Scholar] [CrossRef]
- Vandenberk, L.; Garg, A.D.; Verschuere, T.; Koks, C.; Belmans, J.; Beullens, M.; Agostinis, P.; De Vleeschouwer, S.; Van Gool, S.W. Irradiation of necrotic cancer cells, employed for pulsing dendritic cells (DCs), potentiates DC vaccine-induced antitumor immunity against high-grade glioma. Oncoimmunology 2015, 5, e1083669. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Calero-Pérez, P.; Villamañan, L.; Arias-Ramos, N.; Pumarola, M.; Ortega-Martorell, S.; Julià-Sapé, M.; Arús, C.; Candiota, A.P. Anti-tumour immune response in GL261 glioblastoma generated by Temozolomide Immune-Enhancing Metronomic Schedule monitored with MRSI-based nosological images. NMR Biomed. 2020, 33, 11. [Google Scholar] [CrossRef]
- Zagzag, D.; Miller, D.C.; Chiriboga, L.; Yee, H.; Newcomb, E.W. Green fluorescent protein immunohistochemistry as a novel experimental tool for the detection of glioma cell invasion in vivo. Brain Pathol. 2003, 13, 34–37. [Google Scholar] [CrossRef]
- Zenkoh, J.; Gerelchuluun, A.; Wang, Y.; Miwa, Y.; Ohno, T.; Tsuboi, K. The abscopal effect induced by in situ-irradiated peripheral tumor cells in a murine GL261 brain tumor model. Transl. Cancer Res. 2017, 6, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Winkler, F.; Kienast, Y.; Fuhrmann, M.; Von Baumgarten, L.; Burgold, S.; Mitteregger, G.; Kretzschmar, H.; Herms, J. Imaging glioma cell invasion in vivo reveals mechanisms of dissemination and peritumoral angiogenesis. Glia 2009, 57, 1306–1315. [Google Scholar] [CrossRef]
- Genina, E.A.; Bashkatov, A.N.; Tuchina, D.K.; Dyachenko Timoshina, P.A.; Navolokin, N.; Shirokov, A.; Khorovodov, A.; Terskov, A.; Klimova, M.; Mamedova, A.; et al. Optical properties of brain tissues at the different stages of glioma development in rats: Pilot study. Biomed. Opt. Express 2019, 10, 5182–5197. [Google Scholar] [CrossRef] [PubMed]
- Svaasand, L.O.; Ellingsen, R. Optical properties of human brain. Photochem. Photobiol. 1983, 38, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Resende, F.F.; Bai, X.; Del Bel, E.A.; Kirchhoff, F.; Scheller, A.; Titze-de-Almeida, R. Evaluation of TgH(CX3CR1-EGFP) mice implanted with mCherry-GL261 cells as an in vivo model for morphometrical analysis of glioma-microglia interaction. BMC Cancer 2016, 16, 016–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luker, K.E.; Pata, P.; Shemiakina, I.I.; Pereverzeva, A.; Stacer, A.C.; Shcherbo, D.S.; Pletnev, V.Z.; Skolnaja, M.; Lukyanov, K.A.; Luker, G.D.; et al. Comparative study reveals better far-red fluorescent protein for whole body imaging. Sci. Rep. 2015, 5, 10332. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Kawataki, T.; Endo, K.; Miyazawa, K.; Kinouchi, H.; Saitoh, M. Expression of ZEBs in gliomas is associated with invasive properties and histopathological grade. Oncol. Lett. 2018, 16, 1758–1764. [Google Scholar] [CrossRef] [Green Version]
- Hanihara, M.; Kawataki, T.; Oh-Oka, K.; Mitsuka, K.; Nakao, A.; Kinouchi, H. Synergistic antitumor effect with indoleamine 2,3-dioxygenase inhibition and temozolomide in a murine glioma model. J. Neurosurg. 2016, 124, 1594–1601. [Google Scholar] [CrossRef] [Green Version]
- Hua, L.; Fang, M.; Dong, B.; Guo, S.; Cui, C.; Liu, J.; Yao, Y.; Xiao, Y.; Li, X.; Ren, Y.; et al. Attribution of NKG2DL to the inhibition of early stage allogeneic tumors in mice. Oncotarget 2016, 7, 82369–82383. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||||
| Mouse Group | Number of Squares Passed in the Arena | Time in the Arena Center, s | Number of Upright Postures | Integral Fluorescence Intensity of Glioma GL261-Kat Cells in the Brain Tissue, A.U. (Day 35 after Inoculation) | |
| Periphery | Center | ||||
| Control | 40.2 ± 12.2 | 13.4 ± 4.4 | 100.8 ± 33.7 | 1.2 ± 0.5 | 7480 ± 709 |
| GL25 | 16.75 ± 11.6 | 21 ± 7.8 | 245.5 ± 32.5 * | 0 | 12189 ± 2954 |
| GL50 | 11 ± 6.2 * | 14 ± 6.5 | 233.8 ± 49.4 | 0.2 ± 0.1 | 16127 ± 1770 # |
| (B) | |||||
| Mouse Group | Grooming Time, s | Number of Peeks in Holes | Acts of Urination | Acts of Defecation | |
| Control | 3.4 ± 1.5 | 20.6 ± 5.9 | 0.6 ± 0.3 | 1.8 ± 0.9 | |
| GL25 | 1.3 ± 1.4 | 8.5 ± 2.2 * | 0.3 ± 0.1 | 1.3 ± 0.7 | |
| GL50 | 3.8 ± 1.1 | 2.6 ± 1.6 * | 0.4 ± 0.3 | 3.2 ± 1.4 | |
| Experimental Group | Latent Period of Movement to the Dark Chamber Section, s | Integral Fluorescence Intensity of Glioma GL261-Kat Cells in the Brain Tissue, A.U. (Day 35 after Inoculation) | |
|---|---|---|---|
| Training Session | Retesting | ||
| Control | 27.2 ± 7.5 | 279.8 ± 22.1 | 7480 ± 709 |
| GL25 | 54.0 ± 17.7 | 203.9 ± 67.9 | 12,189 ± 2954 |
| GL50 | 78.5 ± 13.8 * | 220.7 ± 54.4 | 16,127 ± 1770 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishchenko, T.A.; Balalaeva, I.V.; Klimenko, M.O.; Brilkina, A.A.; Peskova, N.N.; Guryev, E.L.; Krysko, D.V.; Vedunova, M.V. Far-Red Fluorescent Murine Glioma Model for Accurate Assessment of Brain Tumor Progression. Cancers 2022, 14, 3822. https://doi.org/10.3390/cancers14153822
Mishchenko TA, Balalaeva IV, Klimenko MO, Brilkina AA, Peskova NN, Guryev EL, Krysko DV, Vedunova MV. Far-Red Fluorescent Murine Glioma Model for Accurate Assessment of Brain Tumor Progression. Cancers. 2022; 14(15):3822. https://doi.org/10.3390/cancers14153822
Chicago/Turabian StyleMishchenko, Tatiana A., Irina V. Balalaeva, Maria O. Klimenko, Anna A. Brilkina, Nina N. Peskova, Evgenii L. Guryev, Dmitri V. Krysko, and Maria V. Vedunova. 2022. "Far-Red Fluorescent Murine Glioma Model for Accurate Assessment of Brain Tumor Progression" Cancers 14, no. 15: 3822. https://doi.org/10.3390/cancers14153822