5-Aminolevulinic Acid (5-ALA)-Induced Protoporphyrin IX Fluorescence by Glioma Cells—A Fluorescence Microscopy Clinical Study

 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drug Preparation
2.2. Clinical Material
2.3. Microscopic Detection of 5-ALA
2.4. Lectin Staining and Immunofluorescence
2.5. Antigen Retrieval and Auto-Fluorescence Removal
3. Results
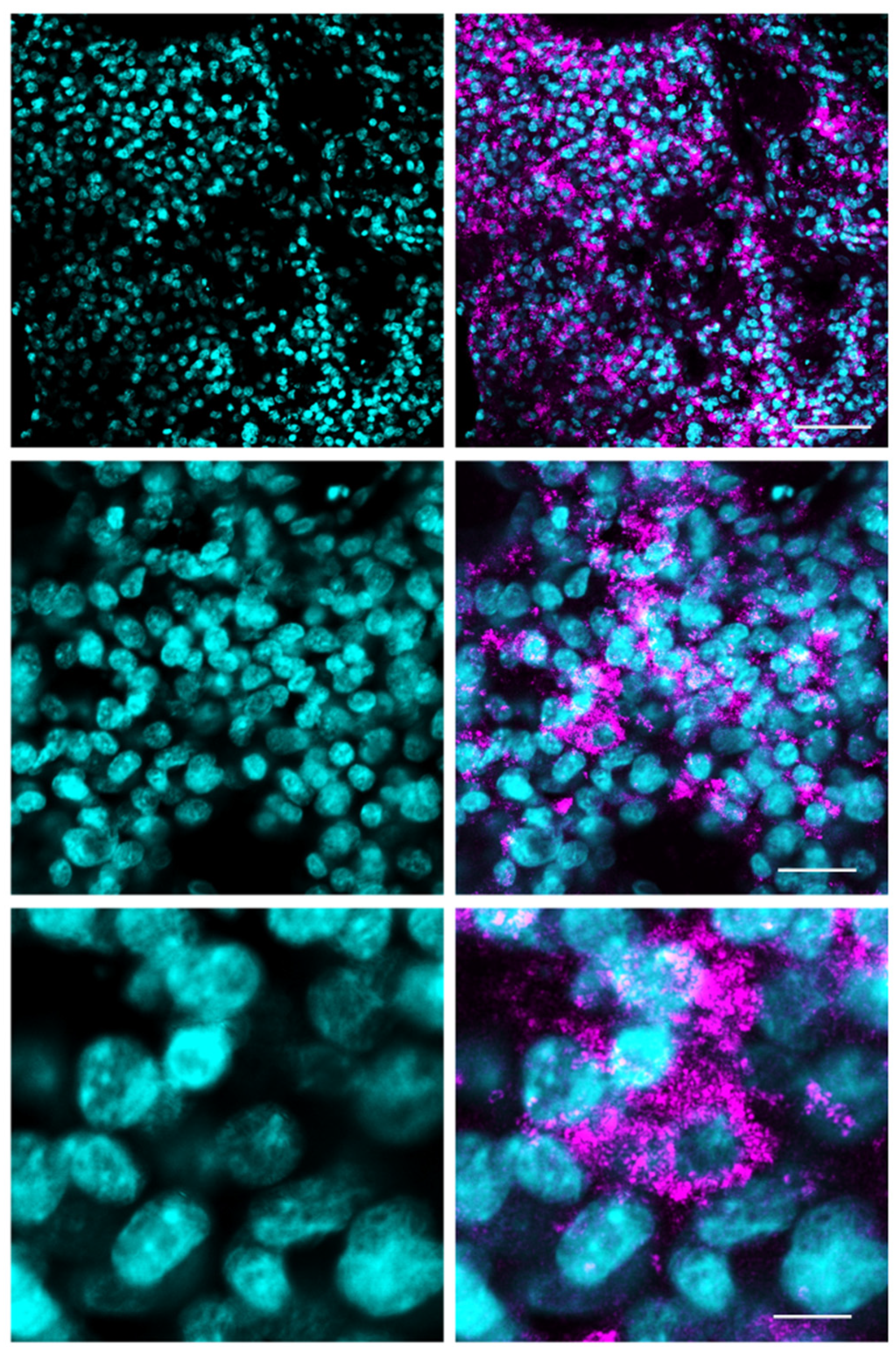
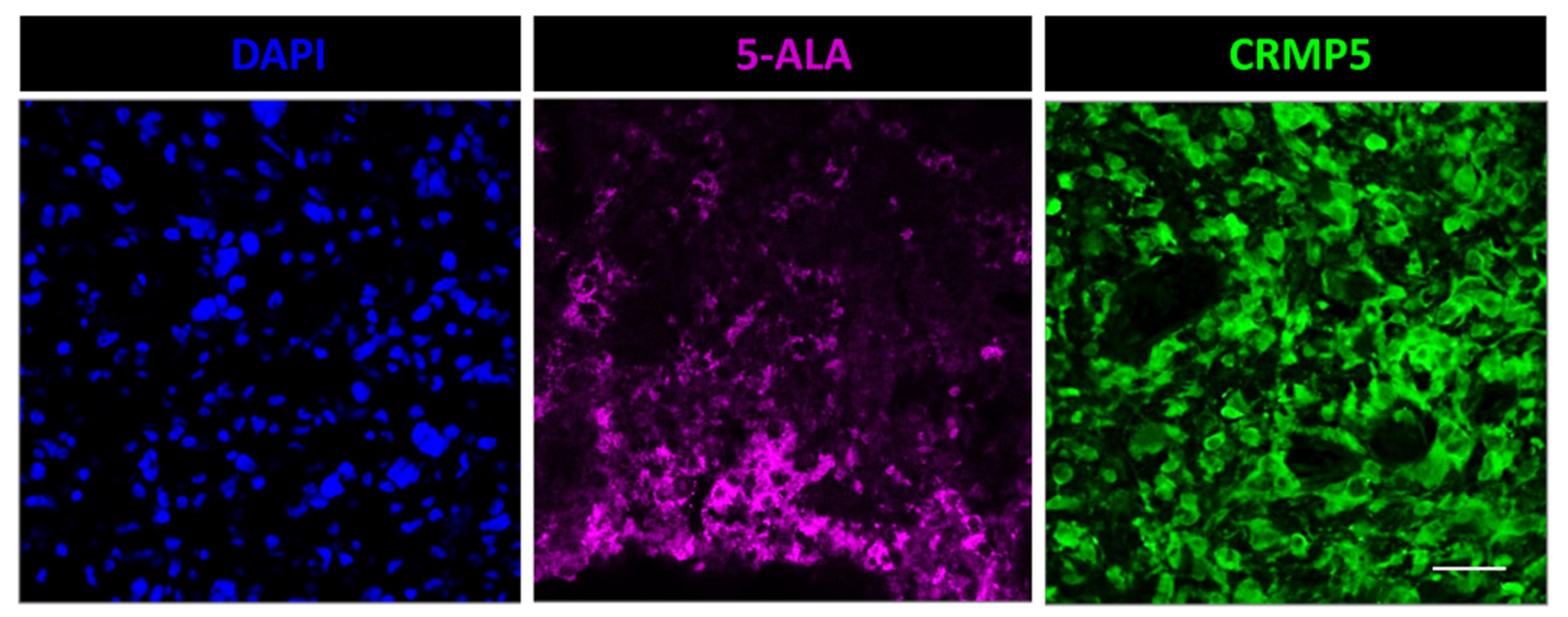
3.1. Fluorescence Microscopy for 5-ALA and Lectin Co-Staining
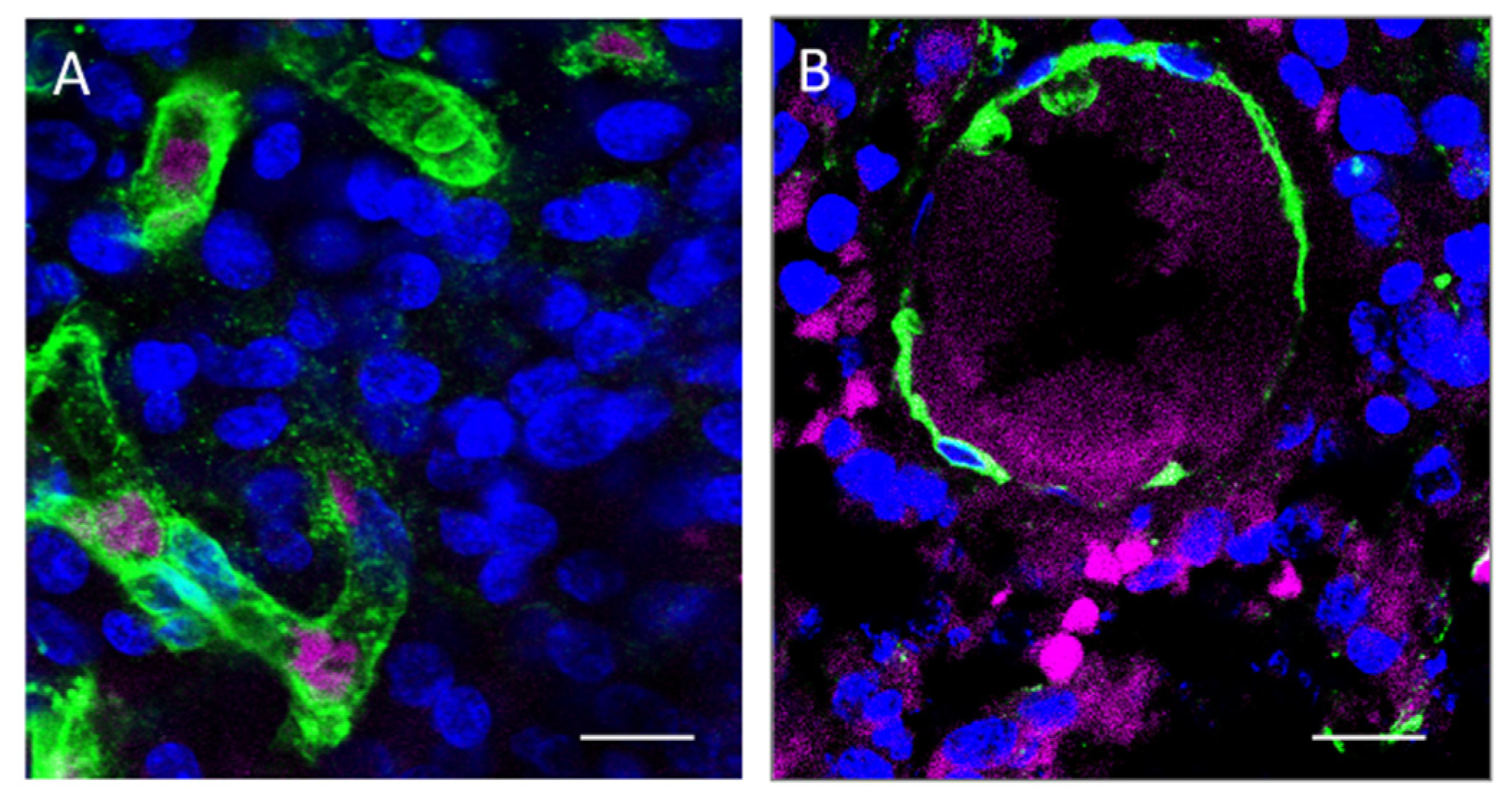
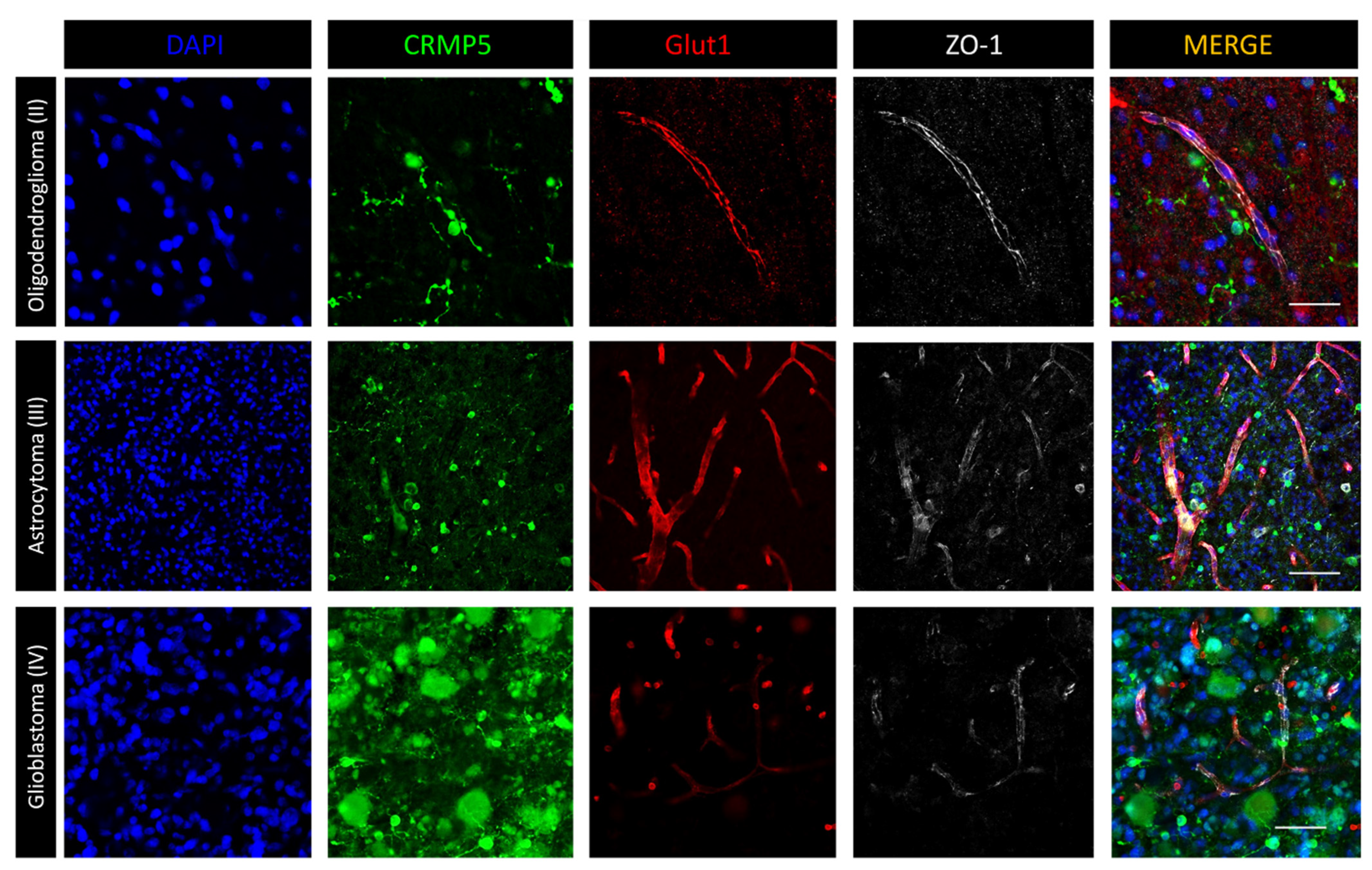
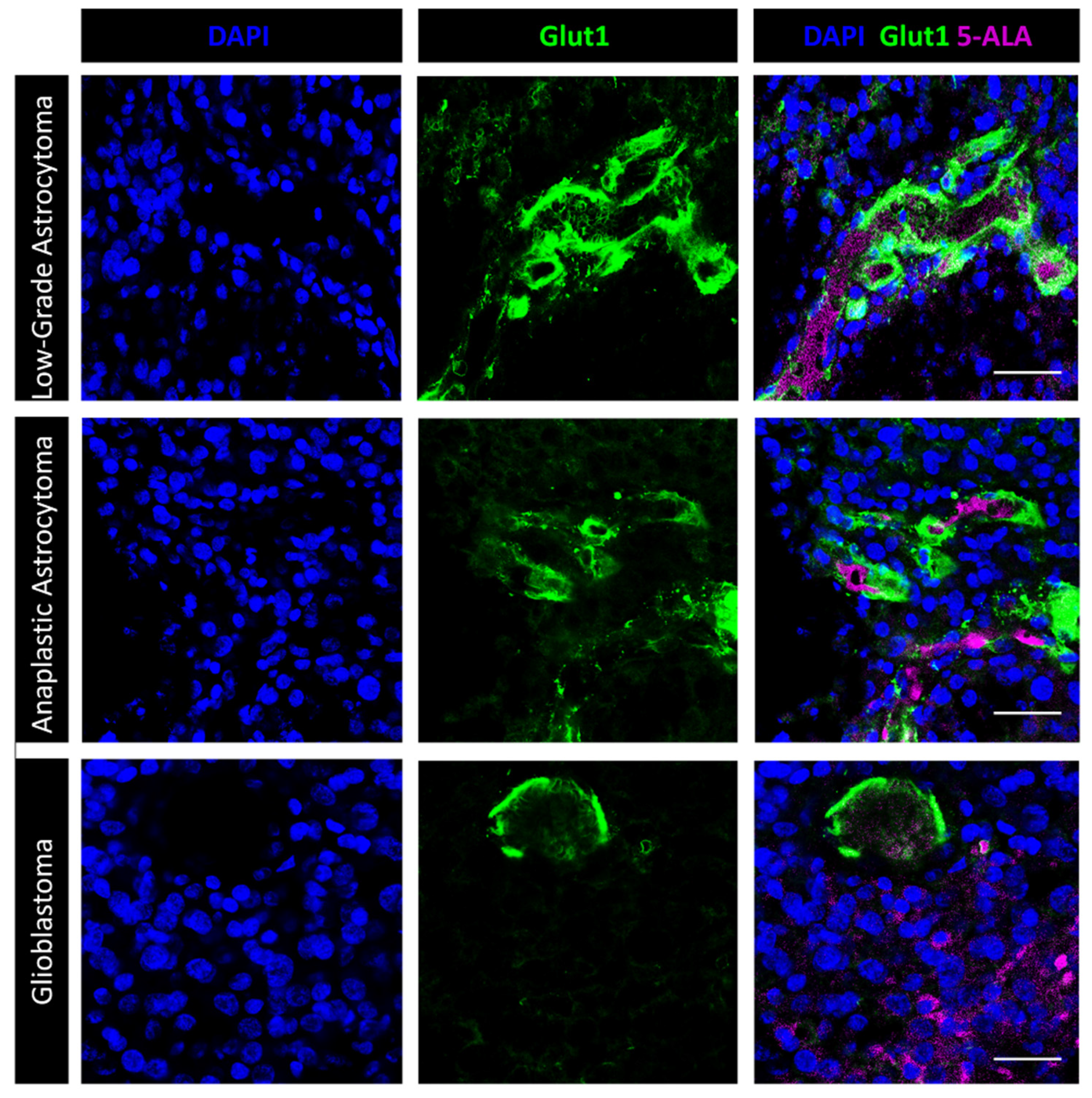
3.2. Immunofluorescence Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Teixidor, P.; Arráez, M.Á.; Villalba, G.; Garcia, R.; Tardáguila, M.; González, J.J.; Rimbau, J.; Vidal, X.; Montané, E. Safety and Efficacy of 5-Aminolevulinic Acid for High Grade Glioma in Usual Clinical Practice: A Prospective Cohort Study. PLoS ONE 2016, 11, e0149244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjipanayis, C.G.; Stummer, W.; Sheehan, J.P. 5-ALA fluorescence-guided surgery of CNS tumors. J. Neurooncol. 2019, 141, 477–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazurek, M.; Kulesza, B.; Stoma, F.; Osuchowski, J.; Mańdziuk, S.; Rola, R. Characteristics of Fluorescent Intraoperative Dyes Helpful in Gross Total Resection of High-Grade Gliomas-A Systematic Review. Diagnostics 2020, 10, 1100. [Google Scholar] [CrossRef]
- Belykh, E.; Shaffer, K.V.; Lin, C.; Byvaltsev, V.A.; Preul, M.C.; Chen, L. Blood-Brain Barrier, Blood-Brain Tumor Barrier, and Fluorescence-Guided Neurosurgical Oncology: Delivering Optical Labels to Brain Tumors. Front. Oncol. 2020, 10, 739. [Google Scholar] [CrossRef] [PubMed]
- Chang, E.; Patel, C.B.; Pohling, C.; Young, C.; Song, J.; Flores, T.A.; Zeng, Y.; Joubert, L.M.; Arami, H.; Natarajan, A.; et al. Tumor treating fields increases membrane permeability in glioblastoma cells. Cell Death Discov. 2018, 4, 113. [Google Scholar] [CrossRef]
- McNicholas, K.; MacGregor, M.N.; Gleadle, J.M. In order for the light to shine so brightly, the darkness must be present-why do cancers fluoresce with 5-aminolaevulinic acid? Br. J. Cancer 2019, 121, 631–639. [Google Scholar] [CrossRef]
- Mĺkvy, P.; Messmann, H.; Regula, J.; Conio, M.; Pauer, M.; Millson, C.E.; MacRobert, A.J.; Bown, S.G. Sensitization and photodynamic therapy (PDT) of gastrointestinal tumors with 5-aminolaevulinic acid (ALA) induced protoporphyrin IX (PPIX). A pilot study. Neoplasma 1995, 42, 109–113. [Google Scholar]
- Stummer, W.; Pichlmeier, U.; Meinel, T.; Wiestler, O.D.; Zanella, F.; Reulen, H.J.; ALA-Glioma Study Group. Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: A randomised controlled multicentre phase III trial. Lancet Oncol. 2006, 7, 392–401. [Google Scholar] [CrossRef]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K. (Eds.) WHO Classification of Tumours of the Central Nervous System, 4th ed.; IARC: Lyon, France, 2016. [Google Scholar]
- D’Alessandris, Q.G.; Biffoni, M.; Martini, M.; Runci, D.; Buccarelli, M.; Cenci, T.; Signore, M.; Stancato, L.; Olivi, A.; De Maria, R.; et al. The clinical value of patient-derived glioblastoma tumorspheres in predicting treatment response. Neuro-Oncology 2017, 19, 1097–1108. [Google Scholar] [CrossRef] [Green Version]
- Grande, S.; Palma, A.; Ricci-Vitiani, L.; Luciani, A.M.; Buccarelli, M.; Biffoni, M.; Molinari, A.; Calcabrini, A.; D’Amore, E.; Guidoni, L.; et al. Metabolic Heterogeneity Evidenced by MRS among Patient-Derived Glioblastoma Multiforme Stem-Like Cells Accounts for Cell Clustering and Different Responses to Drugs. Stem Cells Int. 2018, 2018, 3292704. [Google Scholar] [CrossRef] [Green Version]
- Hagiya, Y.; Endo, Y.; Yonemura, Y.; Takahashi, K.; Ishizuka, M.; Abe, F.; Tanaka, T.; Okura, I.; Nakajima, M.; Ishikawa, T.; et al. Pivotal roles of peptide transporter PEPT1 and ATP-binding cassette (ABC) transporter ABCG2 in 5-aminolevulinic acid (ALA)-based photocytotoxicity of gastric cancer cells in vitro. Photodiagn. Photodyn. Ther. 2012, 9, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, Y.; Ishii, T.; Kohda, T.; Ishizuka, M.; Yamazaki, K.; Nishimura, Y.; Tanaka, T.; Dan, S.; Nakajima, M. Mechanistic study of PpIX accumulation using the JFCR39 cell panel revealed a role for dynamin 2-mediated exocytosis. Sci. Rep. 2019, 9, 8666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampazzo, E.; Della Puppa, A.; Frasson, C.; Battilana, G.; Bianco, S.; Scienza, R.; Basso, G.; Persano, L. Phenotypic and functional characterization of Glioblastoma cancer stem cells identified trough 5-aminolevulinic acid-assisted surgery. J. Neurooncol. 2014, 116, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.B.; Kockro, R.A.; Actor, B.; Sarnthein, J.; Ludwig Bernays, R. Combining 5-ALA Fluorescence and Intraoperative MRI in Glioblastoma Surgery: A Histology-Based Evaluation. Neurosurgery 2016, 78, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.W.; Valdes, P.A.; Harris, B.T.; Fontaine, K.M.; Hartov, A.; Fan, X.; Ji, S.; Lollis, S.S.; Pogue, B.W.; Leblond, F.; et al. Coregistered fluorescence-enhanced tumor resection of malignant glioma: Relationships between delta-aminolevulinic acid-induced protoporphyrin IX fluorescence, magnetic resonance imagingenhancement, and neuropathological parameters. J. Neurosurg. 2011, 114, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.W.; Olson, J.D.; Evans, L.T.; Kolste, K.K.; Kanick, S.C.; Fan, X.; Bravo, J.J.; Wilson, B.C.; Leblond, F.; Marois, M.; et al. Red-light excitation of protoporphyrin IX fluorescence for subsurface tumor detection. J. Neurosurg. 2018, 128, 1690–1697. [Google Scholar] [CrossRef]
- Haas, T.L.; Sciuto, M.R.; Brunetto, L.; Valvo, C.; Signore, M.; Fiori, M.E.; di Martino, S.; Giannetti, S.; Morgante, L.; Boe, A.; et al. Integrin α7 Is a Functional Marker and Potential Therapeutic Target in Glioblastoma. Cell Stem Cell 2017, 21, 35–50.e9. [Google Scholar] [CrossRef] [Green Version]
- Brot, S.; Malleval, C.; Benetollo, C.; Auger, C.; Meyronet, D.; Rogemond, V.; Honnorat, J.; Moradi-Améli, M. Identification of a new CRMP5 isoform present in the nucleus of cancer cells and enhancing their proliferation. Exp. Cell Res. 2013, 319, 588–599. [Google Scholar] [CrossRef]
- Moutal, A.; Honnorat, J.; Massoma, P.; Désormeaux, P.; Bertrand, C.; Malleval, C.; Watrin, C.; Chounlamountri, N.; Mayeur, M.E.; Besançon, R.; et al. CRMP5 controls glioblastoma cell proliferation and survival through Notch-dependent signaling. Cancer Res. 2015, 75, 3519–3528. [Google Scholar] [CrossRef] [Green Version]
- Falchetti, M.L.; D’Alessandris, Q.G.; Pacioni, S.; Buccarelli, M.; Morgante, L.; Giannetti, S.; Lulli, V.; Martini, M.; Larocca, L.M.; Vakana, E.; et al. Glioblastoma endothelium drives bevacizumab-induced infiltrative growth via modulation of PLXDC1. Int. J. Cancer 2019, 144, 1331–1344. [Google Scholar] [CrossRef] [Green Version]
- Pacioni, S.; D’Alessandris, Q.G.; Buccarelli, M.; Boe, A.; Martini, M.; Larocca, L.M.; Bolasco, G.; Ricci-Vitiani, L.; Falchetti, M.L.; Pallini, R. Brain Invasion along Perivascular Spaces by Glioma Cells: Relationship with Blood-Brain Barrier. Cancers 2019, 12, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Palasuberniam, P.; Myers, K.A.; Wang, C.; Chen, B. Her2 oncogene transformation enhances 5-aminolevulinic acid-mediated protoporphyrin IX productioand photodynamic therapy response. Oncotarget 2016, 7, 57798–57810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, D.; Hervey-Jumper, S.L.; Chang, S.; Molinaro, A.M.; McDermott, M.W.; Phillips, J.J.; Berger, M.S. A prospective Phase II clinical trial of 5-aminolevulinic acid to assess the correlation of intraoperative fluorescence intensity and degree of histologic cellularity during resection of high-grade gliomas. J. Neurosurg. 2016, 124, 1300–1309. [Google Scholar] [CrossRef] [Green Version]
- Ennis, S.R.; Novotny, A.; Xiang, J.; Shakui, P.; Masada, T.; Stummer, W.; Smith, D.E.; Keep, R.F. Transport of 5-aminolevulinic acid between blood and brain. Brain Res. 2003, 959, 226–234. [Google Scholar] [CrossRef]
- Kiesel, B.; Mischkulnig, M.; Woehrer, A.; Martinez-Moreno, M.; Millesi, M.; Mallouhi, A.; Czech, T.; Preusser, M.; Hainfellner, J.A.; Wolfsberger, S.; et al. Systematic histopathological analysis of different 5-aminolevulinic acidinduced fluorescence levels in newly diagnosed glioblastomas. J. Neurosurg. 2018, 129, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Stummer, W.; Stocker, S.; Wagner, S.; Stepp, H.; Fritsch, C.; Goetz, C.; Goetz, A.E.; Kiefmann, R.; Reulen, H.J. Intraoperative detection of malignant gliomas by 5-aminolevulinic acid-induced porphyrin fluorescence. Neurosurgery 1998, 42, 518–525. [Google Scholar] [CrossRef]
- Belykh, E.; Miller, E.J.; Patel, A.A.; Bozkurt, B.; Yağmurlu, K.; Robinson, T.R.; Nakaji, P.; Spetzler, R.F.; Lawton, M.T.; Nelson, L.Y.; et al. Optical characterization of neurosurgical operating microscopes: Quantitative fluorescence and assessment of PpIX photobleaching. Sci. Rep. 2018, 8, 12543. [Google Scholar] [CrossRef]
- Wei, L.; Chen, Y.; Yin, C.; Borwege, S.; Sanai, N.; Liu, J.T.C. Optical-sectioning microscopy of protoporphyrin IX fluorescence in human gliomas: Standardization and quantitative comparison with histology. J. Biomed. Opt. 2017, 22, 46005. [Google Scholar] [CrossRef]
- Jaber, M.; Ewelt, C.; Wölfer, J.; Brokinkel, B.; Thomas, C.; Hasselblatt, M.; Grauer, O.; Stummer, W. Is visible aminolevulinic acid-induced fluorescence an independent biomarker for prognosis in histologically confirmed (World Health Organization 2016) low grade gliomas? Neurosurgery 2019, 84, 1214–1224. [Google Scholar] [CrossRef]
- Olivo, M.; Wilson, B.C. Mapping ALA-induced PPIX fluorescence in normal brain and brain tumour using confocal fluorescence microscopy. Int. J. Oncol. 2004, 25, 37–45. [Google Scholar] [CrossRef]
- Hebeda, K.M.; Saarnak, A.E.; Olivo, M.; Sterenborg, H.J.; Wolbers, J.G. 5-Aminolevulinic acid induced endogenous porphyrin fluorescence in 9L and C6 brain tumours and in the normal rat brain. Acta Neurochir. 1998, 140, 503–512. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Age/Sex | Tumor Location | No. Samples | Histology (WHO Grade) | Molecular Profile | Comment |
|---|---|---|---|---|---|---|
| 1 | 40/M | R frontal | 3 | Astrocytoma (II) | IDH mutant | |
| 2 | 34/M | R frontal | 4 | Oligodendroglioma (II) | IDH mutant | |
| 3 | 59/M | R parietal | 4 | Oligodendroglioma (II) | IDH mutant | Recurrent |
| 4 | 49/M | L frontal | 2 | Astrocytoma (III) | IDH mutant | |
| 5 | 40/F | R frontal | 3 | Astrocytoma (III) | IDH wild type | Recurrent |
| 6 | 44/F | L frontal | 4 | Oligodendroglioma (III) | IDH mutant | |
| 7 | 29/M | R frontal | 2 | Oligodendroglioma (III) | IDH mutant | |
| 8 | 42/M | L temporal | 3 | Oligodendroglioma (III) | IDH mutant | |
| 9 | 48/M | R occipital | 2 | Glioblastoma (IV) | IDH1 wild type, EGFRvIII + | Recurrent |
| 10 | 43/M | R temporal | 1 | Glioblastoma (IV) | IDH wild type | |
| 11 | 65/M | L frontal | 1 | Glioblastoma (IV) | IDH mutant | |
| 12 | 72/F | R temporal | 2 | Glioblastoma (IV) | IDH1 wild type, EGFRvIII + | |
| 13 | 54/M | R parietal | 1 | Glioblastoma (IV) | IDH1 wild type, EGFRvIII + | |
| 14 | 68/F | R temporal | 3 | Glioblastoma (IV) | IDH1 wild type, EGFRvIII + | |
| 15 | 76/F | R frontal | 1 | Glioblastoma (IV) | IDH wild type | |
| 16 | 85/F | L temporal | 2 | Glioblastoma (IV) | IDH wild type | |
| 17 | 50/M | L temporal | 3 | Glioblastoma (IV) | IDH wild type | Recurrent |
| 18 | 75/M | R parietal | 1 | Glioblastoma (IV) | IDH wild type | |
| 19 | 48/M | L frontal | 3 | Glioblastoma (IV) | IDH mutant | Secondary |
| 20 | 58/F | L frontal | 2 | Glioblastoma (IV) | IDH wild type | Recurrent |
| 21 | 43/F | R frontal | 1 | Glioblastoma (IV) | IDH wild type | |
| 22 | 34/F | R frontal | 3 | Glioblastoma (IV) | IDH mutant | Secondary |
| 23 | 78/M | L temporal | 2 | Glioblastoma (IV) | IDH wild type | |
| 24 | 54/M | R temporal | 1 | Glioblastoma (IV) | IDH wild type | |
| 25 | 55/M | R frontal | 1 | Glioblastoma (IV) | IDH wild type | |
| 26 | 70/M | R temporal | 3 | Glioblastoma (IV) | IDH wild type | |
| 27 | 68/M | R frontal | 1 | Glioblastoma (IV) | IDH wild type |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacioni, S.; D’Alessandris, Q.G.; Giannetti, S.; Della Pepa, G.M.; Offi, M.; Giordano, M.; Caccavella, V.M.; Falchetti, M.L.; Lauretti, L.; Pallini, R. 5-Aminolevulinic Acid (5-ALA)-Induced Protoporphyrin IX Fluorescence by Glioma Cells—A Fluorescence Microscopy Clinical Study. Cancers 2022, 14, 2844. https://doi.org/10.3390/cancers14122844
Pacioni S, D’Alessandris QG, Giannetti S, Della Pepa GM, Offi M, Giordano M, Caccavella VM, Falchetti ML, Lauretti L, Pallini R. 5-Aminolevulinic Acid (5-ALA)-Induced Protoporphyrin IX Fluorescence by Glioma Cells—A Fluorescence Microscopy Clinical Study. Cancers. 2022; 14(12):2844. https://doi.org/10.3390/cancers14122844
Chicago/Turabian StylePacioni, Simone, Quintino Giorgio D’Alessandris, Stefano Giannetti, Giuseppe Maria Della Pepa, Martina Offi, Martina Giordano, Valerio Maria Caccavella, Maria Laura Falchetti, Liverana Lauretti, and Roberto Pallini. 2022. "5-Aminolevulinic Acid (5-ALA)-Induced Protoporphyrin IX Fluorescence by Glioma Cells—A Fluorescence Microscopy Clinical Study" Cancers 14, no. 12: 2844. https://doi.org/10.3390/cancers14122844