
Effector Mechanisms of CD8+ HLA-DR+ T Cells in Breast Cancer Patients Who Respond to Neoadjuvant Chemotherapy

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Patients and Samples
2.2. Surface Staining and Analysis by Flow Cytometry
2.3. Cell Culture and Intracellular Phenotyping by Flow Cytometry
2.4. Determination of Plasmatic Concentration of IL-12 and IFN-γ
2.5. Statistics
3. Results
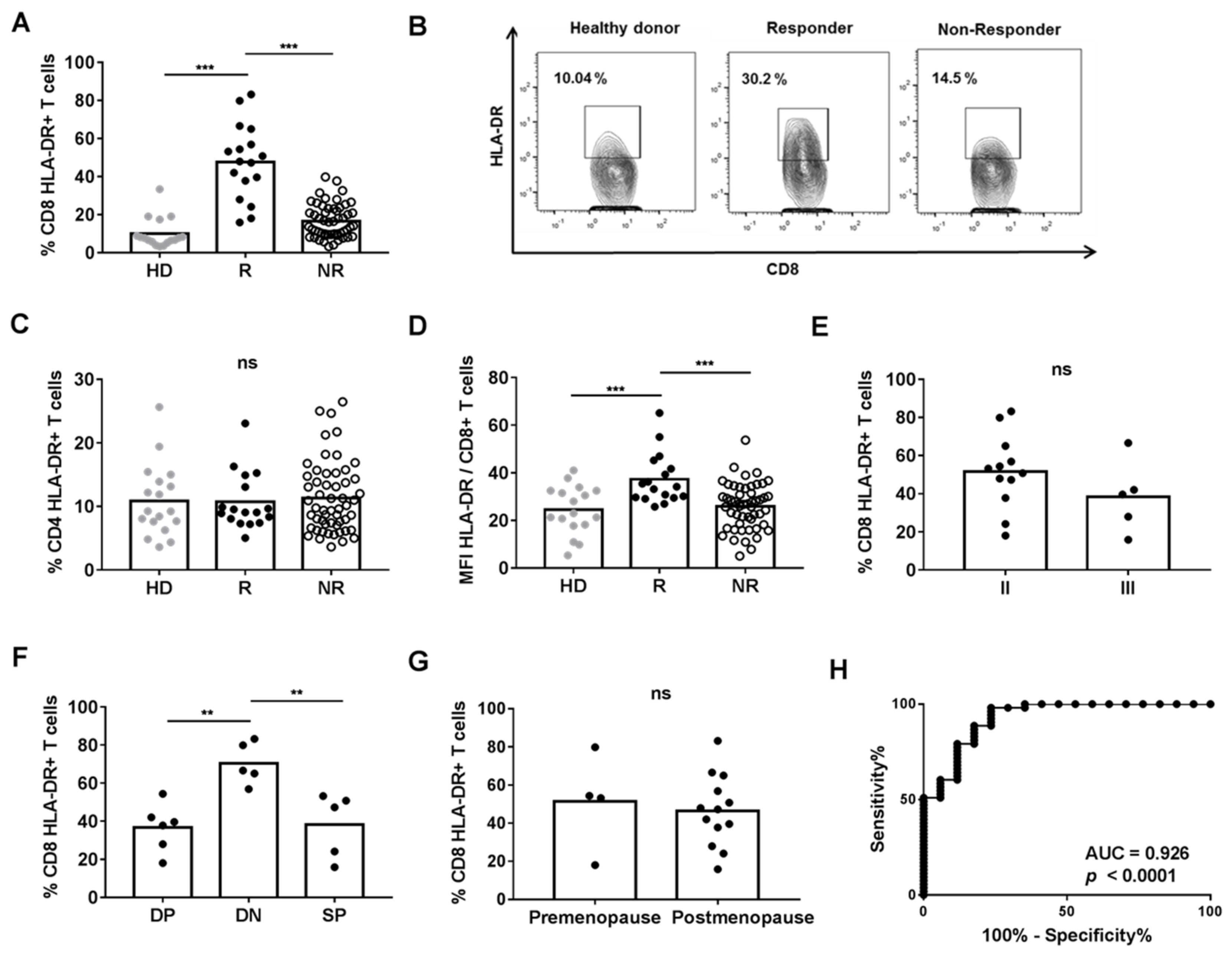
3.1. HLA-DR Expression on CD8+ T Cells Was Increased in R Patients
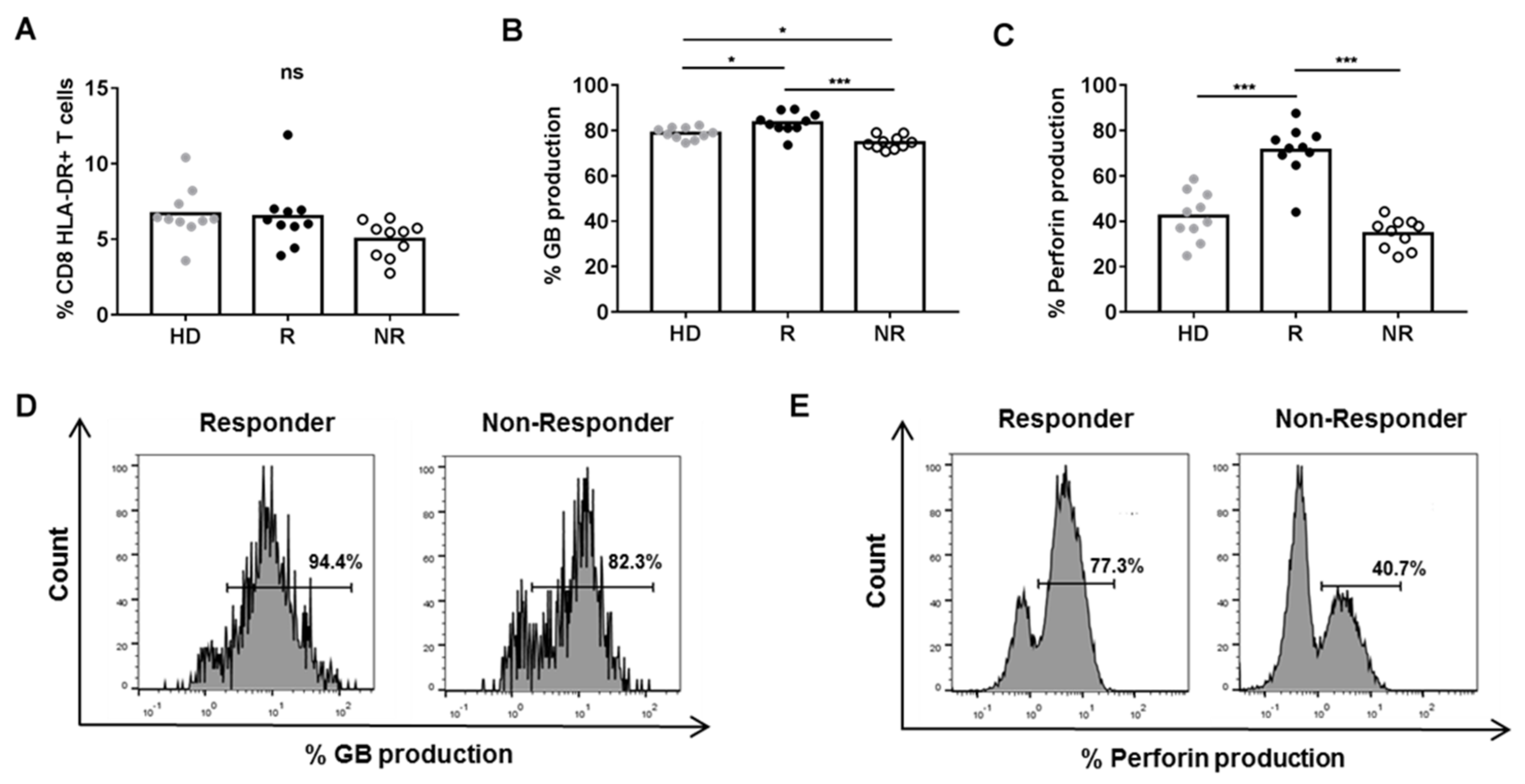
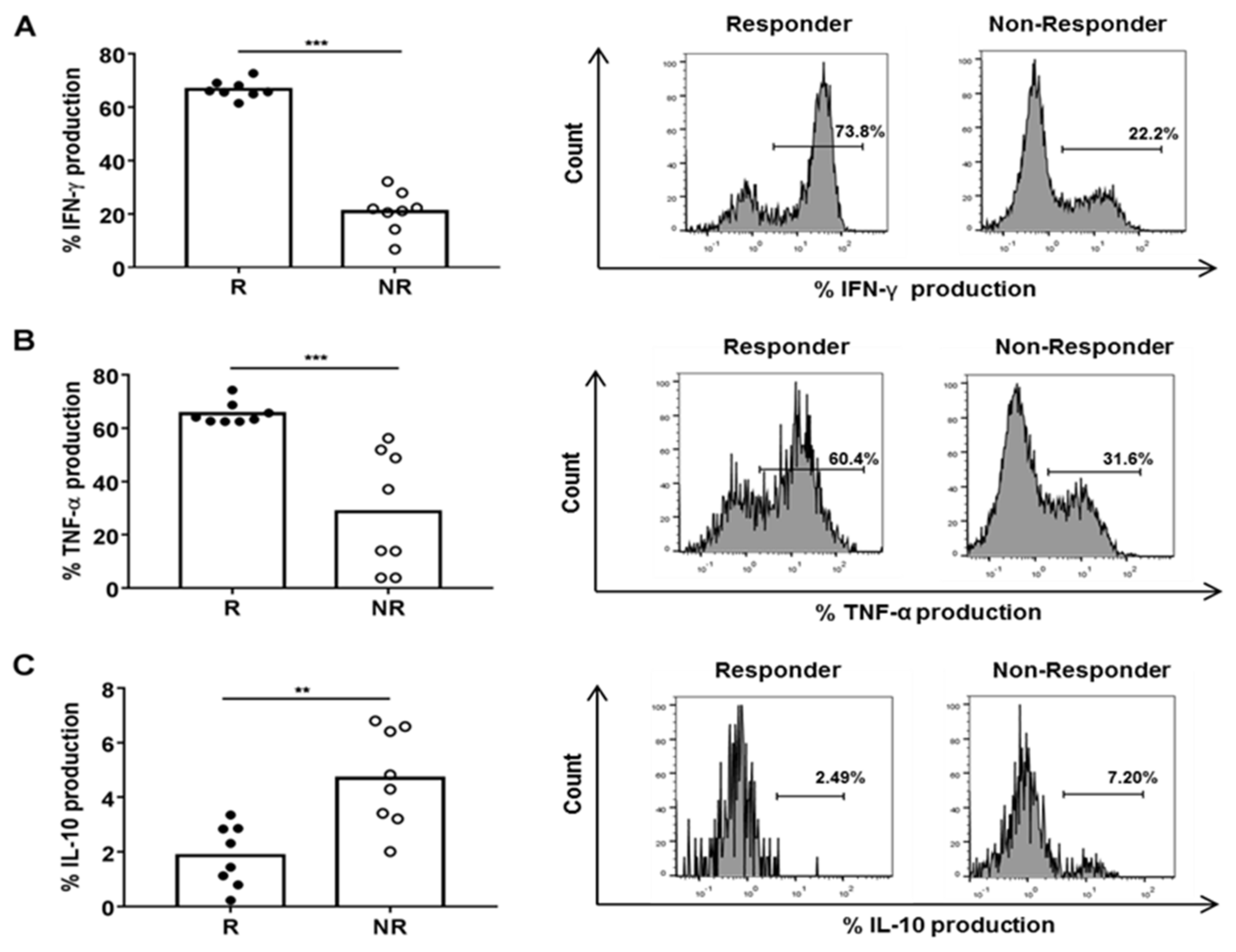
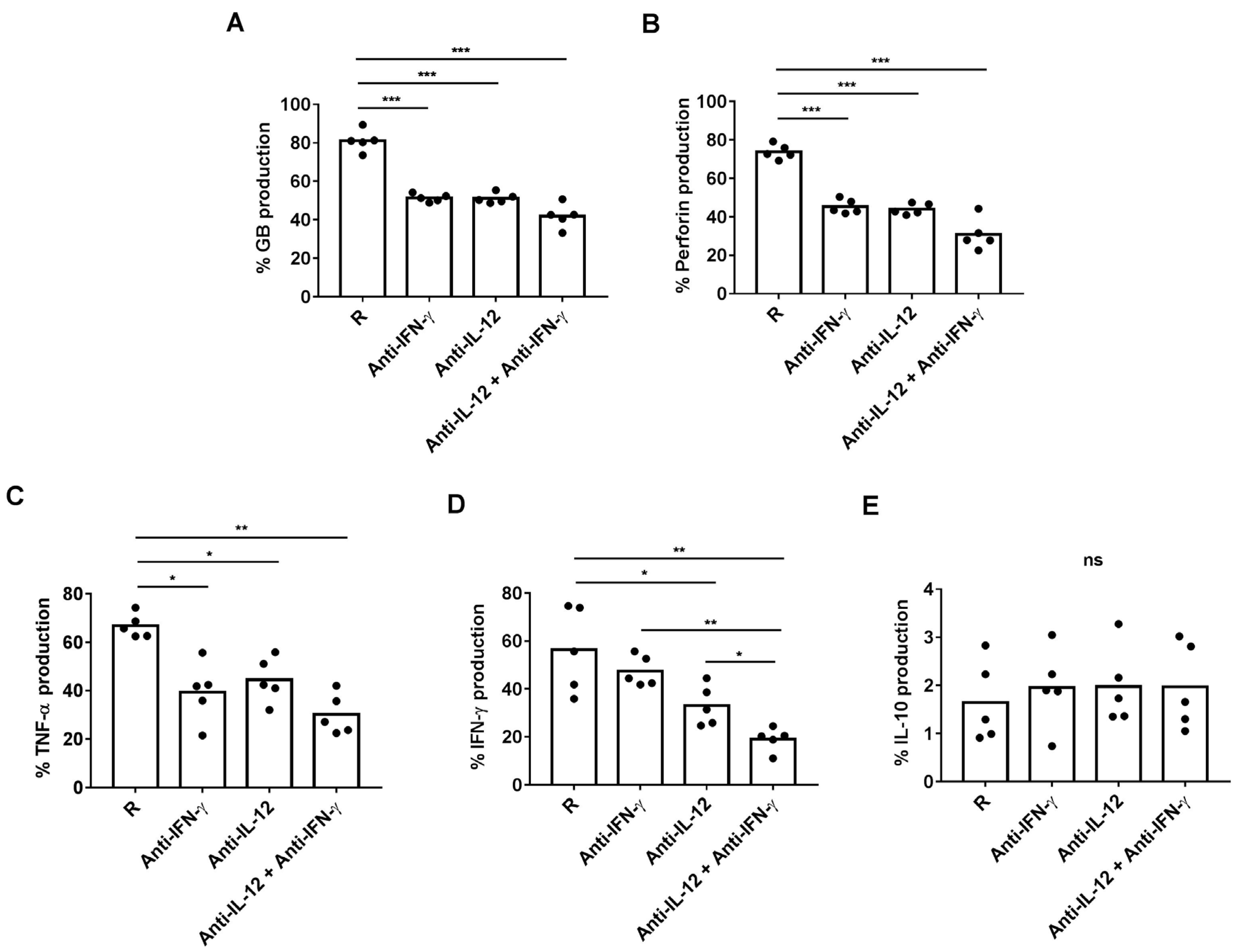
3.2. Plasma from R Patients Increased Cytotoxic and Pro-Inflammatory Activity of CD8+ HLA-DR+ T Cells
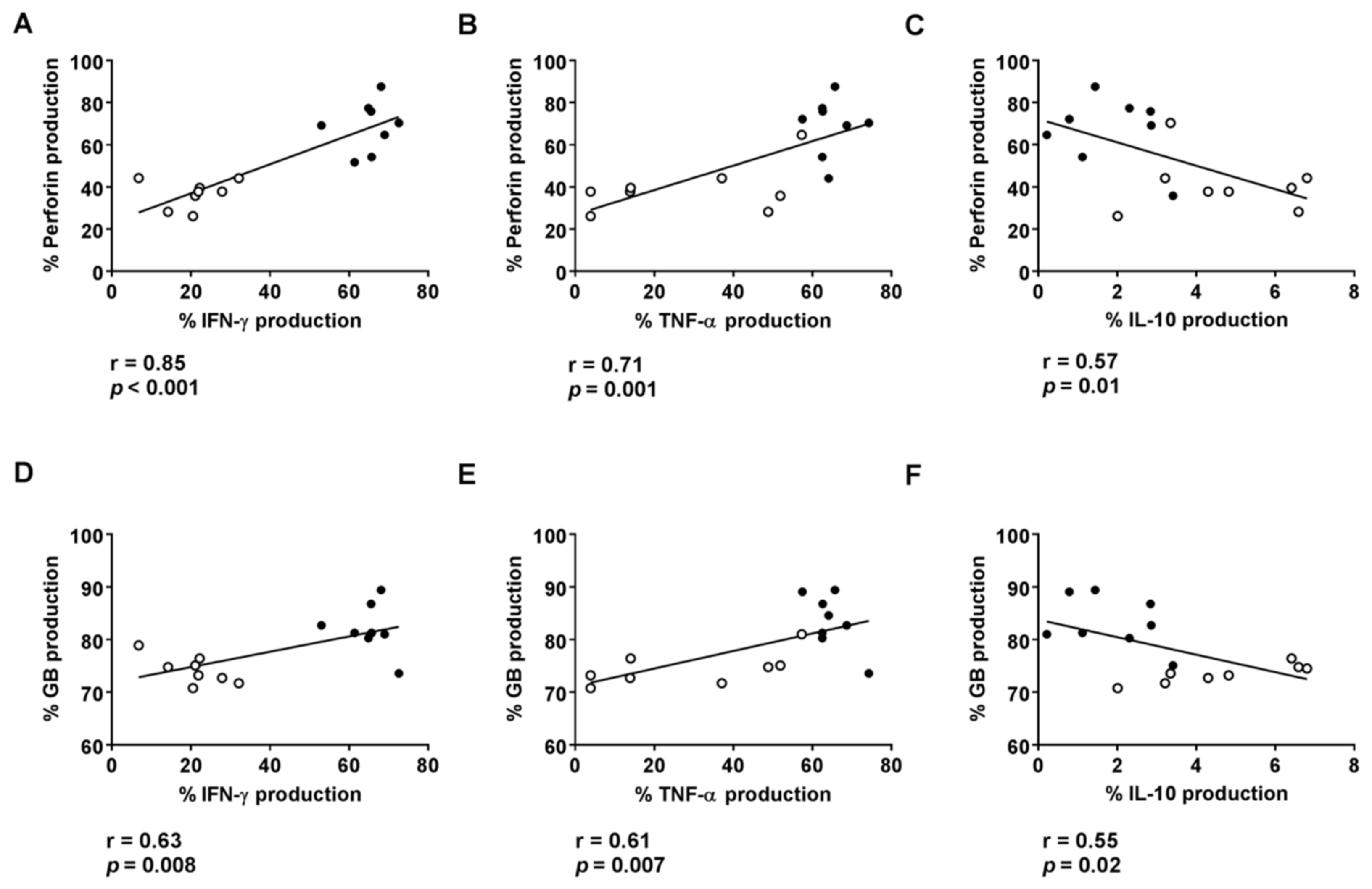
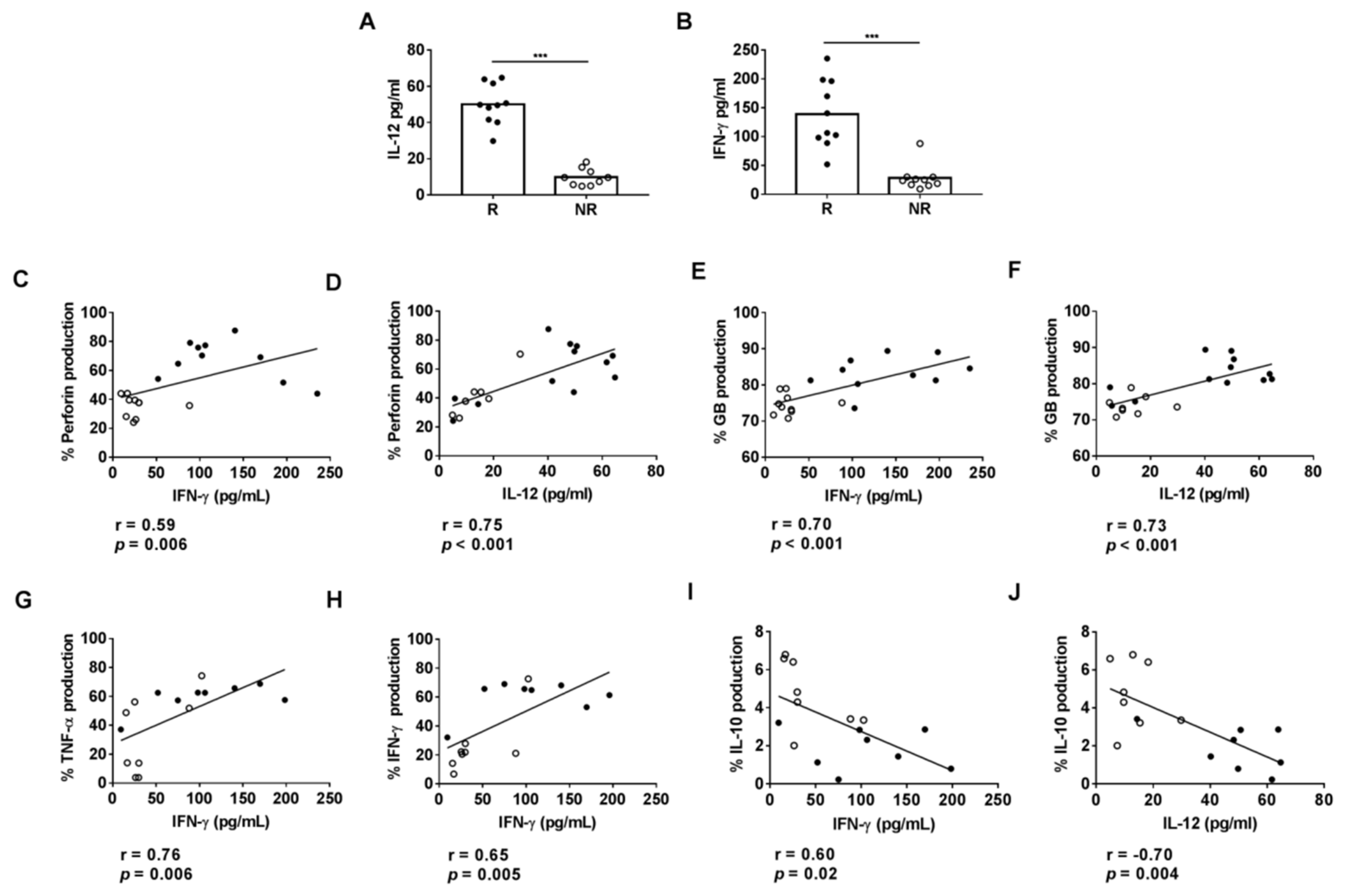
3.3. Plasmatic Levels of IL-12 and IFN-γ Are Associated with Cytotoxic Activity on HLA-DR CD8+ T Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- König, L.; Mairinger, F.D.; Hoffmann, O.; Bittner, A.-K.; Schmid, K.W.; Kimmig, R.; Kasimir-Bauer, S.; Bankfalvi, A. Dissimilar patterns of tumor-infiltrating immune cells at the invasive tumor front and tumor center are associated with response to neoadjuvant chemotherapy in primary breast cancer. BMC Cancer 2019, 19, 120. [Google Scholar] [CrossRef] [PubMed]
- Oshi, M.; Asaoka, M.; Tokumaru, Y.; Yan, L.; Matsuyama, R.; Ishikawa, T.; Endo, I.; Takabe, K. CD8 T Cell Score as a Prognostic Biomarker for Triple Negative Breast Cancer. Int. J. Mol. Sci. 2020, 21, 6968. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Seo, J.-Y.; Ahn, J.-H.; Ahn, S.-H.; Gong, G. Tumor-Associated Lymphocytes Predict Response to Neoadjuvant Chemotherapy in Breast Cancer Patients. J. Breast Cancer 2013, 16, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladoire, S.; Arnould, L.; Apetoh, L.; Coudert, B.; Martin, F.; Chauffert, B.; Fumoleau, P.; Ghiringhelli, F. Pathologic Complete Response to Neoadjuvant Chemotherapy of Breast Carcinoma Is Associated with the Disappearance of Tumor-Infiltrating Foxp3+ Regulatory T Cells. Clin. Cancer Res. 2008, 14, 2413–2420. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, D.P.; Jacinto, A.; Borralho, P.; Braga, S.; Cabral, M.G. HLA-DR in Cytotoxic T Lymphocytes Predicts Breast Cancer Patients’ Response to Neoadjuvant Chemotherapy. Front. Immunol. 2018, 9, 2605. [Google Scholar] [CrossRef] [Green Version]
- Tewari, M.; Krishnamurthy, A.; Shukla, H.S. Predictive markers of response to neoadjuvant chemotherapy in breast cancer. Surg. Oncol. 2008, 17, 301–311. [Google Scholar] [CrossRef]
- Okano, M.; Oshi, M.; Butash, A.L.; Asaoka, M.; Katsuta, E.; Peng, X.; Qi, Q.; Yan, L.; Takabe, K. Estrogen Receptor Positive Breast Cancer with High Expression of Androgen Receptor has Less Cytolytic Activity and Worse Response to Neoadjuvant Chemotherapy but Better Survival. Int. J. Mol. Sci. 2019, 20, 2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieci, M.V.; Mathieu, M.C.; Guarneri, V.; Conte, P.; Delaloge, S.; Andre, F.; Goubar, A. Prognostic and predictive value of tumor-infiltrating lymphocytes in two phase III randomized adjuvant breast cancer trials. Ann. Oncol. 2015, 26, 1698–1704. [Google Scholar] [CrossRef]
- García-Martínez, E.; Gil, G.L.; Benito, A.C.; González-Billalabeitia, E.; Conesa, M.A.V.; García, T.G.; García-Garre, E.; Vicente, V.; De La Peña, F.A. Tumor-infiltrating immune cell profiles and their change after neoadjuvant chemotherapy predict response and prognosis of breast cancer. Breast Cancer Res. 2014, 16, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demaria, S.; Formenti, S.C. Role of T lymphocytes in tumor response to radiotherapy. Front. Oncol. 2012, 2, 95. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, M.; Sasano, H.; Tamaki, K.; Hirakawa, H.; Takahashi, Y.; Nakagawa, S.; Watanabe, G.; Tada, H.; Suzuki, A.; Ohuchi, N.; et al. Prognostic significance of tumor-infiltrating CD8+ and FOXP3+ lymphocytes in residual tumors and alterations in these parameters after neoadjuvant chemotherapy in triple-negative breast cancer: A retrospective multicenter study. Breast Cancer Res. 2015, 17, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.-Y.; Wu, C.Y.-J.; Fang, J.-H.; Chen, H.-C.; Feng, L.-Y.; Huang, C.-Y.; Wei, K.-C.; Fang, J.-Y.; Lin, C.-Y. Functional Change of Effector Tumor-Infiltrating CCR5+CD38+HLA-DR+CD8+ T Cells in Glioma Microenvironment. Front. Immunol. 2019, 10, 2395. [Google Scholar] [CrossRef] [PubMed]
- Predina, J.D.; Haas, A.R.; Martinez, M.; O’Brien, S.; Moon, E.K.; Woodruff, P.; Stadanlick, J.; Corbett, C.; Frenzel-Sulyok, L.; Bryski, M.G.; et al. Neoadjuvant Gene-Mediated Cytotoxic Immunotherapy for Non-Small-Cell Lung Cancer: Safety and Immunologic Activity. Mol. Ther. 2021, 29, 658–670. [Google Scholar] [CrossRef]
- Gonzalez, S.M.; Taborda, N.A.; Rugeles, M.T. Role of Different Subpopulations of CD8+ T Cells during HIV Exposure and Infection. Front. Immunol. 2017, 8, 936. [Google Scholar] [CrossRef]
- Perdomo-Celis, F.; Velilla, P.A.; Taborda, N.A.; Rugeles, M.T. An altered cytotoxic program of CD8+ T-cells in HIV-infected patients despite HAART-induced viral suppression. PLoS ONE 2019, 14, e0210540. [Google Scholar] [CrossRef]
- Schmidt, M.E.; Knudson, C.J.; Hartwig, S.M.; Pewe, L.L.; Meyerholz, D.; Langlois, R.A.; Harty, J.; Varga, S.M. Memory CD8 T cells mediate severe immunopathology following respiratory syncytial virus infection. PLoS Pathog. 2018, 14, e1006810. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Liu, C.; Fu, R.; Wang, H.; Wang, J.; Liu, X.; Feng, L.; Li, L.; Liu, H.; Wang, H.; et al. CD8+HLA-DR+ T cells are increased in patients with severe aplastic anemia. Mol. Med. Rep. 2014, 10, 1252–1258. [Google Scholar] [CrossRef] [Green Version]
- Rea, I.M.; McNerlan, S.E.; Alexander, H.D. CD69, CD25, and HLA-DR activation antigen expression on CD3+ lymphocytes and relationship to serum TNF-alpha, IFN-gamma, and sIL-2R levels in aging. Exp Gerontol. 1999, 34, 79–93. [Google Scholar] [CrossRef]
- Guarneri, V.; Dieci, M.; Bisagni, G.; Frassoldati, A.; Bianchi, G.V.; De Salvo, G.; Orvieto, E.; Urso, L.; Pascual, T.; Pare, L.; et al. De-escalated therapy for HR+/HER2+ breast cancer patients with Ki67 response after 2-week letrozole: Results of the PerELISA neoadjuvant study. Ann. Oncol. 2019, 30, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, K.; Patel, T.; Niravath, P.; Rodriguez, A.; Darcourt, J.; Belcheva, A.; Boone, T.; Ensor, J.; Chang, J. Targeting mTOR and DNA repair pathways in residual triple negative breast cancer post neoadjuvant chemotherapy. Sci. Rep. 2021, 11, 82. [Google Scholar] [CrossRef]
- Zamora, C.; Cantó, E.; Nieto, J.C.; Bardina, J.; Diaz-Torné, C.; Moya, P.; Magallares, B.; Ortiz, M.A.; Julià, G.; Juarez, C.; et al. Binding of Platelets to Lymphocytes: A Potential Anti-Inflammatory Therapy in Rheumatoid Arthritis. J. Immunol. 2017, 198, 3099–3108. [Google Scholar] [CrossRef] [PubMed]
- Lebossé, F.; Gudd, C.; Tunc, E.; Singanayagam, A.; Nathwani, R.; Triantafyllou, E.; Pop, O.; Kumar, N.; Mukherjee, S.; Hou, T.Z.; et al. CD8+ T cells from patients with cirrhosis display a phenotype that may contribute to cirrhosis-associated immune dysfunction. EBioMedicine 2019, 49, 258–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, L.D.; Khayumbi, J.; Ongalo, J.; Sasser, L.E.; Tonui, J.; Campbell, A.; Odhiambo, F.H.; Ouma, S.G.; Alter, G.; Gandhi, N.R.; et al. Distinct Human NK Cell Phenotypes and Functional Responses to Mycobacterium tuberculosis in Adults from TB Endemic and Non-endemic Regions. Front. Cell. Infect. Microbiol. 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Däubener, W.; Wanagat, N.; Pilz, K.; Seghrouchni, S.; Fischer, H.G.; Hadding, U. A new, simple, bioassay for human IFN-γ. J. Immunol. Methods 1994, 168, 39–47. [Google Scholar] [CrossRef]
- Jiang, X.; Orr, B.A.; Kranz, D.M.; Shapiro, D.J. Estrogen Induction of the Granzyme B Inhibitor, Proteinase Inhibitor 9, Protects Cells against Apoptosis Mediated by Cytotoxic T Lymphocytes and Natural Killer Cells. Endocrinology 2006, 147, 1419–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, S.; Lécuroux, C.; Sáez-Cirión, A.; Pancino, G.; Girault, I.; Versmisse, P.; Boufassa, F.; Taulera, O.; Sinet, M.; Lambotte, O.; et al. Potential Role for HIV-Specific CD38−/HLA-DR+ CD8+ T Cells in Viral Suppression and Cytotoxicity in HIV Controllers. PLoS ONE 2014, 9, e101920. [Google Scholar] [CrossRef] [PubMed]
- Tassi, E.; Grazia, G.; Vegetti, C.; Bersani, I.; Bertolini, G.; Molla, A.; Baldassari, P.; Andriani, F.; Roz, L.; Sozzi, G.; et al. Early Effector T Lymphocytes Coexpress Multiple Inhibitory Receptors in Primary Non–Small Cell Lung Cancer. Cancer Res. 2016, 77, 851–861. [Google Scholar] [CrossRef] [Green Version]
- Noble, A.; Giorgini, A.; Leggat, J.A. Cytokine-induced IL-10–secreting CD8 T cells represent a phenotypically distinct suppressor T-cell lineage. Blood 2006, 107, 4475–4483. [Google Scholar] [CrossRef]
- Holling, T.M.; Schooten, E.; Elsen, P.J.V.D. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.C.; Matthews, S.; Yap, G.S. IL-12 Signaling Drives CD8+T Cell IFN-γ Production and Differentiation of KLRG1+Effector Subpopulations during Toxoplasma gondii Infection. J. Immunol. 2008, 180, 5935–5945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtsinger, J.M.; Valenzuela, J.O.; Agarwal, P.; Lins, D.C.; Mescher, M.F. Cutting Edge: Type I IFNs Provide a Third Signal to CD8 T Cells to Stimulate Clonal Expansion and Differentiation. J. Immunol. 2005, 174, 4465–4469. [Google Scholar] [CrossRef] [Green Version]
- Starbeck-Miller, G.; Badovinac, V.; Barber, D.L.; Harty, J.T. Cutting Edge: Expression of FcγRIIB Tempers Memory CD8 T Cell Function In Vivo. J. Immunol. 2014, 192, 35–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottinghaus, E.K.; Vesosky, B.; Turner, J. Interleukin-12 is sufficient to promote antigen-independent interferon-γ production by CD8 T cells in old mice. Immunology 2009, 128, e679–e690. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.J.; Ornelles, D.A.; Mitchell, L.M.; Brzoza-Lewis, K.L.; Hiltbold, E.M. IL-12 Produced by Dendritic Cells Augments CD8+ T Cell Activation through the Production of the Chemokines CCL1 and CCL17. J. Immunol. 2008, 181, 8576–8584. [Google Scholar] [CrossRef] [Green Version]
- Tough, D.F.; Zhang, X.; Sprent, J. An IFN-gamma-dependent pathway controls stimulation of memory phenotype CD8+ T cell turnover in vivo by IL-12, IL-18, and IFN-gamma. J Immunol. 2001, 166, 6007–6011. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lin, W.; Chen, H.; Xu, Z.; Ye, Y.; Chen, M. Dual-target IL-12-containing nanoparticles enhance T cell functions for cancer immunotherapy. Cell. Immunol. 2020, 349, 1040–1042. [Google Scholar] [CrossRef]
- Ahn, H.J.; Maruo, S.; Tomura, M.; Mu, J.; Hamaoka, T.; Nakanishi, K.; Clark, S.; Kurimoto, M.; Okamura, H.; Fujiwara, H. A mechanism underlying synergy between IL-12 and IFN-gamma-inducing factor in enhanced production of IFN-gamma. J. Immunol. 1997, 159, 2125–2131. [Google Scholar] [PubMed]
- Zhang, X.; Starnbach, M.N. An Excess of the Proinflammatory Cytokines IFN-γ and IL-12 Impairs the Development of the Memory CD8+T Cell Response toChlamydia trachomatis. J. Immunol. 2015, 195, 1665–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marana, H.R.C.; da Silva, J.S.; de Andrade, J.M. NK Cell Activity in the Presence of IL-12 Is a Prognostic Assay to Neoadjuvant Chemotherapy in Cervical Cancer. Gynecol. Oncol. 2000, 78, 318–323. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Characteristics | Classification | Value |
|---|---|---|
| Median age (range), years | 58 (36–84) | |
| Menopause status, n (%) | Pre-menopause | 21 (28.7%) |
| Post-menopause | 52 (71.2%) | |
| Hormone receptor, n (%) | ER | |
| + | 56 (76.7%) | |
| − | 17 (23.2%) | |
| PR | ||
| − | 47 (64.3%) | |
| + | 26 (35.6%) | |
| Cancer stage, n (%) | II | 42 (57.5%) |
| III | 31 (35.6%) | |
| Ki67, median (range), % | 25% (2–85%) | |
| Dimension, median (range), mm | 45 (10–120) | |
| Axillary lymph node invasion status, % | Negative | 47.3% |
| Positive | 52.7% | |
| Residual cancer burden (RCB), % | 0-pCR | 14.9% |
| I | 8.1% | |
| II | 52.7% | |
| III | 23% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osuna-Gómez, R.; Arqueros, C.; Galano, C.; Mulet, M.; Zamora, C.; Barnadas, A.; Vidal, S. Effector Mechanisms of CD8+ HLA-DR+ T Cells in Breast Cancer Patients Who Respond to Neoadjuvant Chemotherapy. Cancers 2021, 13, 6167. https://doi.org/10.3390/cancers13246167
Osuna-Gómez R, Arqueros C, Galano C, Mulet M, Zamora C, Barnadas A, Vidal S. Effector Mechanisms of CD8+ HLA-DR+ T Cells in Breast Cancer Patients Who Respond to Neoadjuvant Chemotherapy. Cancers. 2021; 13(24):6167. https://doi.org/10.3390/cancers13246167
Chicago/Turabian StyleOsuna-Gómez, Rubén, Cristina Arqueros, Carla Galano, Maria Mulet, Carlos Zamora, Agustí Barnadas, and Silvia Vidal. 2021. "Effector Mechanisms of CD8+ HLA-DR+ T Cells in Breast Cancer Patients Who Respond to Neoadjuvant Chemotherapy" Cancers 13, no. 24: 6167. https://doi.org/10.3390/cancers13246167