
Characterization of Immunoactive and Immunotolerant CD4+ T Cells in Breast Cancer by Measuring Activity of Signaling Pathways That Determine Immune Cell Function


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Signaling Pathway Activity Analysis of Preclinical and Clinical Studies
2.2. General Rules for the Interpretation of the Signal Transduction Pathway Activity Scores
- (1)
- On the same sample, the log2 odds pathway activity scores cannot be compared between different signaling pathways, since each of the signaling pathways has its own range in the log2 odds activity scores;
- (2)
- The log2 odds range for the pathway activity (minimum–maximum activities) may vary, depending on the cell type. Once the range is defined using samples with a known pathway activity, on every new sample, the absolute value can be directly interpreted against that reference. If the range is not defined, only differences in the log2 odds activity score between samples can be interpreted;
- (3)
- The pathway activity scores are highly quantitative, and even small differences in log2 odds can be reproducible and meaningful;
- (4)
- A negative log2 odds ratio does not mean that the pathway is inactive.
2.3. Microarray Data Source and Quality Control
2.4. Statistics
2.5. Defining a Threshold for the Abnormal Pathway Activity in Blood CD4+ T Cell Samples
3. Results
3.1. Measuring Activity of Signal Transduction Pathways, Using Validated Assays
3.2. Signaling Pathway Activities in Resting and Activated CD4+ T Cells, in CD4+-Derived Th1 and Th2 Cells, and in CD4+ Treg Cells
3.3. Immune-Suppressive Effect of the Cancer Cell SN on the Activated CD4+ T Cells
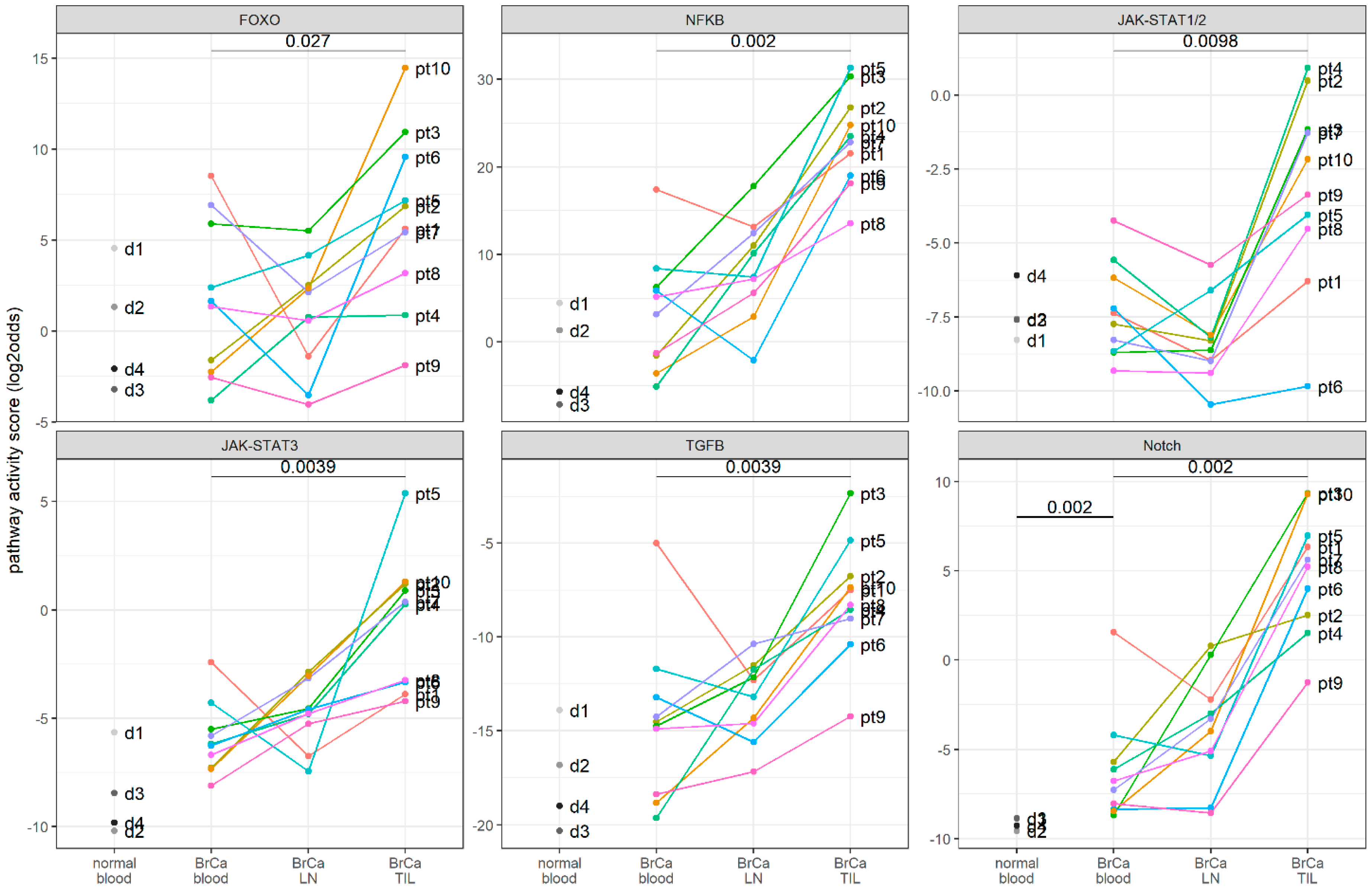
3.4. Clinical Study: Signaling Pathway Activity in CD4+ T Cells Derived from Blood, Lymph Nodes, and Breast Cancer Tissue Samples of Patients with Primary Breast Cancer
3.5. A Threshold for the Abnormal Pathway Activity in Blood CD4+ T Cell Samples
4. Discussion
5. Limitations of the Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Han, X. Anti–PD-1/PD-L1 Therapy of Human Cancer: Past, Present, and Future. J. Clin. Invest. 2015, 125, 3384–3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emens, L.A. Breast Cancer Immunotherapy: Facts and Hopes. Clin. Cancer Res. 2018, 24, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Hendry, S.; Salgado, R.; Gevaert, T.; Russell, P.A.; John, T.; Thapa, B.; Christie, M.; van de Vijver, K.; Estrada, M.V.; Gonzalez-Ericsson, P.I.; et al. Assessing Tumor-Infiltrating Lymphocytes in Solid Tumors: A Practical Review for Pathologists and Proposal for a Standardized Method From the International Immunooncology Biomarkers Working Group: Part 1: Assessing the Host Immune Response, TILs in Invasive Breast Carcinoma and Ductal Carcinoma In Situ, Metastatic Tumor Deposits and Areas for Further Research. Adv. Anat. Pathol. 2017, 24, 235–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendry, S.; Salgado, R.; Gevaert, T.; Russell, P.A.; John, T.; Thapa, B.; Christie, M.; van de Vijver, K.; Estrada, M.V.; Gonzalez-Ericsson, P.I.; et al. Assessing Tumor-Infiltrating Lymphocytes in Solid Tumors: A Practical Review for Pathologists and Proposal for a Standardized Method from the International Immuno-Oncology Biomarkers Working Group: Part 2: TILs in Melanoma, Gastrointestinal Tract Carcinomas, Non-Small Cell Lung Carcinoma and Mesothelioma, Endometrial and Ovarian Carcinomas, Squamous Cell Carcinoma of the Head and Neck, Genitourinary Carcinomas, and Primary Brain Tumors. Adv. Anat. Pathol. 2017, 24, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Solinas, C.; Aiello, M.; De Silva, P.; Gu-Trantien, C.; Migliori, E.; Willard-Gallo, K. Targeting PD-1 in Cancer: Biological Insights with a Focus on Breast Cancer. Crit. Rev. Oncol. Hematol. 2019, 142, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Stenström, J.; Hedenfalk, I.; Hagerling, C. Regulatory T Lymphocyte Infiltration in Metastatic Breast Cancer-an Independent Prognostic Factor That Changes with Tumor Progression. Breast Cancer Res. 2021, 23, 27. [Google Scholar] [CrossRef]
- Plitas, G.; Konopacki, C.; Wu, K.; Bos, P.D.; Morrow, M.; Putintseva, E.V.; Chudakov, D.M.; Rudensky, A.Y. Regulatory T Cells Exhibit Distinct Features in Human Breast Cancer. Immunity 2016, 45, 1122–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.M.; Patterson, S.J.; Levings, M.K. The Role of the PI3K Signaling Pathway in CD4+ T Cell Differentiation and Function. Front. Immunol. 2012, 3, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platanias, L.C. Mechanisms of Type-I- and Type-II-Interferon-Mediated Signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.; Ghosh, S. NF-ΚB: Roles and Regulation In Different CD4+ T Cell Subsets. Immunol. Rev. 2013, 252, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.O.; Wan, Y.Y.; Flavell, R.A. T Cell-Produced Transforming Growth Factor-Β1 Controls T Cell Tolerance and Regulates Th1- and Th17-Cell Differentiation. Immunity 2007, 26, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Goodman, W.A.; Young, A.B.; McCormick, T.S.; Cooper, K.D.; Levine, A.D. Stat3 Phosphorylation Mediates Resistance of Primary Human T Cells to Regulatory T Cell Suppression. J. Immunol. 2011, 186, 3336–3345. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Pal, S.K.; Reckamp, K.; Figlin, R.A.; Yu, H. STAT3: A Target to Enhance Antitumor Immune Response. Curr. Top. Microbiol. Immunol. 2011, 344, 41–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaegh, W.; van Ooijen, H.; Inda, M.A.; Hatzis, P.; Versteeg, R.; Smid, M.; Martens, J.; Foekens, J.; van de Wiel, P.; Clevers, H.; et al. Selection of Personalized Patient Therapy through the Use of Knowledge-Based Computational Models That Identify Tumor-Driving Signal Transduction Pathways. Cancer Res. 2014, 74, 2936–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ooijen, H.; Hornsveld, M.; Veen, C.D.; Velter, R.; Dou, M.; Verhaegh, W.; Burgering, B.; van de Stolpe, A. Assessment of Functional PI3K Pathway Activity in Cancer Tissue Using FOXO Target Gene Expression in a Knowledge-Based Computational Model. Am. J. Pathol. 2018, 188, 1956–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Stolpe, A.; Holtzer, L.; van Ooijen, H.; de Inda, M.A.; Verhaegh, W. Enabling Precision Medicine by Unravelling Disease Pathophysiology: Quantifying Signal Transduction Pathway Activity across Cell and Tissue Types. Sci. Rep. 2019, 9, 1603. [Google Scholar] [CrossRef] [Green Version]
- Van de Stolpe, A. Quantitative Measurement of Functional Activity of the PI3K Signaling Pathway in Cancer. Cancers 2019, 11, 293. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, W.; Verhaegh, W.; Holtzer, L.; van de Stolpe, A. Measurement of Cellular Immune Response to Viral Infection and Vaccination. Front. Immunol. 2020, 11, 575074. [Google Scholar] [CrossRef]
- Gu-Trantien, C.; Loi, S.; Garaud, S.; Equeter, C.; Libin, M.; de Wind, A.; Ravoet, M.; Le Buanec, H.; Sibille, C.; Manfouo-Foutsop, G.; et al. CD4+ Follicular Helper T Cell Infiltration Predicts Breast Cancer Survival. J. Clin. Invest. 2013, 123, 2873–2892. [Google Scholar] [CrossRef]
- Canté-Barrett, K.; Holtzer, L.; van Ooijen, H.; Hagelaar, R.; Cordo’, V.; Verhaegh, W.; van de Stolpe, A.; Meijerink, J.P.P. A Molecular Test for Quantifying Functional Notch Signaling Pathway Activity in Human Cancer. Cancers 2020, 12, 3142. [Google Scholar] [CrossRef]
- van de Stolpe, A.; Verhaegh, W.; Blay, J.-Y.; Ma, C.X.; Pauwels, P.; Pegram, M.; Prenen, H.; De Ruysscher, D.; Saba, N.F.; Slovin, S.F.; et al. RNA Based Approaches to Profile Oncogenic Pathways From Low Quantity Samples to Drive Precision Oncology Strategies. Front. Genet. 2021, 11, 598118. [Google Scholar] [CrossRef] [PubMed]
- GEO Database. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Gds/ (accessed on 24 June 2020).
- Kanduri, K.; Tripathi, S.; Larjo, A.; Mannerström, H.; Ullah, U.; Lund, R.; Hawkins, R.D.; Ren, B.; Lähdesmäki, H.; Lahesmaa, R. Identification of Global Regulators of T-Helper Cell Lineage Specification. Genome Med. 2015, 7, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Chen, H.; Probst-Kepper, M.; Geffers, R.; Eifes, S.; del Sol, A.; Schughart, K.; Zeng, A.-P.; Balling, R. PLAU Inferred from a Correlation Network Is Critical for Suppressor Function of Regulatory T Cells. Mol. Syst. Biol. 2012, 8, 624. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.L.; Miller, C.J. Simpleaffy: A BioConductor Package for Affymetrix Quality Control and Data Analysis. Bioinformatics 2005, 21, 3683–3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heber, S.; Sick, B. Quality Assessment of Affymetrix GeneChip Data. OMICS 2006, 10, 358–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parman, C.; Halling, C.; Gentleman, R. AffyQCReport: QC Report Generation for AffyBatch Objects, R Package Version 1.58.0. 2018.
- Gautier, L.; Cope, L.; Bolstad, B.M.; Irizarry, R.A. Affy--Analysis of Affymetrix GeneChip Data at the Probe Level. Bioinformatics 2004, 20, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Hillmer, E.J.; Zhang, H.; Li, H.S.; Watowich, S.S. STAT3 Signaling in Immunity. Cytokine Growth Factor Rev. 2016, 31, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fung, M.M.; Rohwer, F.; McGuire, K.L. IL-2 Activation of a PI3K-Dependent STAT3 Serine Phosphorylation Pathway in Primary Human T Cells. Cell. Signal. 2003, 15, 625–636. [Google Scholar] [CrossRef]
- Campbell, J.D.; Cook, G.; Robertson, S.E.; Fraser, A.; Boyd, K.S.; Gracie, J.A.; Franklin, I.M. Suppression of IL-2-Induced T Cell Proliferation and Phosphorylation of STAT3 and STAT5 by Tumor-Derived TGF Beta Is Reversed by IL-15. J. Immunol. 2001, 167, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Rudd, C.E.; Schneider, H. Unifying Concepts in CD28, ICOS and CTLA4 Co-Receptor Signalling. Nat. Rev. Immunol. 2003, 3, 544–556. [Google Scholar] [CrossRef]
- Riley, J.L. PD-1 Signaling in Primary T Cells. Immunol. Rev. 2009, 229, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-C. The Non-Canonical NF-ΚB Pathway in Immunity and Inflammation. Nat. Rev. Immunol. 2017, 17, 545–558. [Google Scholar] [CrossRef]
- Asano, N.; Watanabe, T.; Kitani, A.; Fuss, I.J.; Strober, W. Notch1 Signaling and Regulatory T Cell Function. J. Immunol. 2008, 180, 2796–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostroukhova, M.; Qi, Z.; Oriss, T.B.; Dixon-McCarthy, B.; Ray, P.; Ray, A. Treg-Mediated Immunosuppression Involves Activation of the Notch-HES1 Axis by Membrane-Bound TGF-Beta. J. Clin. Invest. 2006, 116, 996–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flavell, R.A.; Sanjabi, S.; Wrzesinski, S.H.; Licona-Limón, P. The Polarization of Immune Cells in the Tumour Environment by TGFβ. Nat. Rev. Immunol. 2010, 10, 554–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, R.V.; Chemnitz, J.M.; Frauwirth, K.A.; Lanfranco, A.R.; Braunstein, I.; Kobayashi, S.V.; Linsley, P.S.; Thompson, C.B.; Riley, J.L. CTLA-4 and PD-1 Receptors Inhibit T-Cell Activation by Distinct Mechanisms. Mol. Cell Biol. 2005, 25, 9543–9553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frigola, X.; Inman, B.A.; Lohse, C.M.; Krco, C.J.; Cheville, J.C.; Thompson, R.H.; Leibovich, B.; Blute, M.L.; Dong, H.; Kwon, E.D. Identification of a Soluble Form of B7-H1 That Retains Immunosuppressive Activity and Is Associated with Aggressive Renal Cell Carcinoma. Clin. Cancer Res. 2011, 17, 1915–1923. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Lang, J. Soluble PD-1 and PD-L1: Predictive and Prognostic Significance in Cancer. Oncotarget 2017, 8, 97671–97682. [Google Scholar] [CrossRef] [Green Version]
- Nagato, T.; Ohkuri, T.; Ohara, K.; Hirata, Y.; Kishibe, K.; Komabayashi, Y.; Ueda, S.; Takahara, M.; Kumai, T.; Ishibashi, K.; et al. Programmed Death-Ligand 1 and Its Soluble Form Are Highly Expressed in Nasal Natural Killer/T-Cell Lymphoma: A Potential Rationale for Immunotherapy. Cancer Immunol. Immunother. 2017, 66, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Finkelmeier, F.; Canli, Ö.; Tal, A.; Pleli, T.; Trojan, J.; Schmidt, M.; Kronenberger, B.; Zeuzem, S.; Piiper, A.; Greten, F.R.; et al. High Levels of the Soluble Programmed Death-Ligand (SPD-L1) Identify Hepatocellular Carcinoma Patients with a Poor Prognosis. Eur. J. Cancer 2016, 59, 152–159. [Google Scholar] [CrossRef]
- Yang, Y. Cancer Immunotherapy: Harnessing the Immune System to Battle Cancer. J. Clin. Invest. 2015, 125, 3335–3337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, E.; Massagué, J. Transforming Growth Factor-β Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef] [PubMed]
- Fisher, D.T.; Appenheimer, M.M.; Evans, S.S. The Two Faces of IL-6 in the Tumor Microenvironment. Semin. Immunol. 2014, 26, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frauwirth, K.A.; Riley, J.L.; Harris, M.H.; Parry, R.V.; Rathmell, J.C.; Plas, D.R.; Elstrom, R.L.; June, C.H.; Thompson, C.B. The CD28 Signaling Pathway Regulates Glucose Metabolism. Immunity 2002, 16, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Itoh, Y.; Saitoh, M.; Miyazawa, K. Smad3–STAT3 Crosstalk in Pathophysiological Contexts. Acta Biochim. Biophys Sin. 2018, 50, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haura, E.B.; Turkson, J.; Jove, R. Mechanisms of Disease: Insights into the Emerging Role of Signal Transducers and Activators of Transcription in Cancer. Nat. Clin. Pract. Oncol. 2005, 2, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-M.; Yu, C.-R.; Dambuza, I.; Marrero, B.; Egwuagu, C.E. STAT3 Protein Interacts with Class O Forkhead Transcription Factors in the Cytoplasm and Regulates Nuclear/Cytoplasmic Localization of FoxO1 and FoxO3a Proteins in CD4+ T Cells. J. Biol. Chem. 2012, 287, 30436–30443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohue, Y.; Nishikawa, H. Regulatory T (Treg) Cells in Cancer: Can Treg Cells Be a New Therapeutic Target? Cancer Sci. 2019, 110, 2080–2089. [Google Scholar] [CrossRef]
- Tanaka, A.; Sakaguchi, S. Targeting Treg Cells in Cancer Immunotherapy. Eur. J. Immunol. 2019, 49, 1140–1146. [Google Scholar] [CrossRef] [Green Version]
- Saleh, R.; Elkord, E. Acquired Resistance to Cancer Immunotherapy: Role of Tumor-Mediated Immunosuppression. Semin. Cancer Biol. 2020, 65, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Terranova-Barberio, M.; Pawlowska, N.; Dhawan, M.; Moasser, M.; Chien, A.J.; Melisko, M.E.; Rugo, H.; Rahimi, R.; Deal, T.; Daud, A.; et al. Exhausted T Cell Signature Predicts Immunotherapy Response in ER-Positive Breast Cancer. Nat. Commun. 2020, 11, 3584. [Google Scholar] [CrossRef] [PubMed]
- Ohkura, N.; Sakaguchi, S. Treg Cells Acquire New Directions, Cytokines Navigate. Immunity 2012, 37, 443–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippitz, B.E. Cytokine Patterns in Patients with Cancer: A Systematic Review. Lancet Oncol. 2013, 14, e218–e228. [Google Scholar] [CrossRef]


{kind=link}
| A. | Sample Category | Annotation Per Sample | FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch |
|---|---|---|---|---|---|---|---|---|
| CD4+ naive | replicate 1 | 10.3 | −9.6 | −7.7 | −7.3 | −10.9 | −5.0 | |
| replicate 2 | 12.4 | −8.2 | −8.2 | −8.1 | −10.9 | −3.4 | ||
| replicate 3 | 7.5 | −11.7 | −9.2 | −6.3 | −11.0 | −4.0 | ||
| CD4+ activated | replicate 1 | −3.8 | −1.8 | −2.8 | −3.4 | −18.8 | −4.2 | |
| replicate 2 | −3.5 | −1.7 | −4.4 | −2.5 | −18.9 | −3.9 | ||
| replicate 3 | −4.9 | −0.8 | −2.5 | −1.6 | −18.2 | −4.1 | ||
| B. | Sample Category | Annotation Per Sample | FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch |
| CD4+ activated + interleukin 12 (IL-12) | replicate 1 | −4.9 | 5.3 | −1.1 | −0.1 | −15.6 | −4.5 | |
| replicate 2 | −4.4 | 6.1 | −0.9 | −1.1 | −14.2 | −4.6 | ||
| replicate 3 | −4.8 | 5.2 | −1.3 | −0.3 | −14.6 | −3.9 | ||
| CD4+ activated + IL-4 | replicate 1 | 0.6 | −6.2 | −7.5 | −0.2 | −19.0 | −4.2 | |
| replicate 2 | −0.4 | −5.1 | −7.0 | 0.0 | −18.3 | −3.6 | ||
| replicate 3 | 0.2 | −5.1 | −5.9 | 0.6 | −18.5 | −4.0 | ||
| C. | Sample Category | Annotation Per Sample | FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch |
| Treg, in a resting state | −0.7 | −6.7 | −2.8 | −2.5 | −16.3 | −0.6 | ||
| Treg, in a suppressive state | 180 min | 8.9 | 13.3 | −5.1 | 17.0 | −9.8 | 3.6 | |
| 200 min | 10.6 | 12.4 | −4.7 | 17.1 | −8.9 | 3.3 | ||
| 220 min | 10.0 | 12.7 | −5.2 | 16.6 | −7.9 | 1.6 |
| Sample Category | Annotation per Sample | FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch | |
|---|---|---|---|---|---|---|---|---|
| Not activated | Control | 1 | −2.7 | −10.9 | −8.3 | −6.2 | −14.8 | −7.5 |
| 2 | −2.4 | −10.8 | −8.8 | −6.3 | −14.3 | −7.3 | ||
| 3 | −3.2 | −11.1 | −8.7 | −6.8 | −14.8 | −7.1 | ||
| + cancer tissue supernatant | tumor 1 | −2.6 | −9.1 | −8.3 | −7.5 | −10.1 | −6.8 | |
| tumor 2 | −2.4 | −11.5 | −8.4 | −8.3 | −10.3 | −7.2 | ||
| tumor 3 | −1.7 | −9.2 | −8.5 | −5.5 | −11.7 | −7.4 | ||
| tumor 4 | −2.0 | −11.2 | −8.7 | −7.5 | −13.2 | −7.8 | ||
| Activated (anti-CD3/anti-CD28) | Control | 1 | −7.5 | 3.5 | −6.2 | 6.0 | −15.8 | −4.2 |
| 2 | −7.2 | 2.3 | −5.9 | 5.2 | −16.4 | −4.3 | ||
| 3 | −7.5 | 3.1 | −6.0 | 5.0 | −17.0 | −4.2 | ||
| + cancer tissue supernatant | tumor 1 | 9.1 | −10.0 | −9.0 | −2.1 | −1.4 | −7.3 | |
| tumor 2 | 8.5 | −10.0 | −8.9 | −1.9 | −1.4 | −5.8 | ||
| tumor 3 | 3.5 | −4.4 | −7.0 | −1.1 | −6.5 | −4.8 | ||
| tumor 4 | 5.9 | −5.9 | −8.7 | −1.3 | −3.8 | −4.3 |
| Annotation per Sample | FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch | Age | ER a | PR b | HER2 c | Tumor Size (cm) d | LN Metastasis e | T cell Infiltrate f | Histological Grade g |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TIL pt 3 | 10.9 | 30.3 | −1.2 | 0.9 | −2.4 | 9.3 | 43 | + | + | − | 2.8 | 0/12 | minimal | 3 (SBR8) |
| TIL pt 1 | 5.6 | 21.5 | −6.3 | −3.9 | −7.5 | 6.3 | 51 | + | + | − | 1.8 | 0/16 | minimal | 2 (SBR6) |
| TIL pt 5 | 7.2 | 31.3 | −4.0 | 5.4 | −4.8 | 7.0 | 58 | + | + | − | 4 | 1/10 | minimal | 2 (SBR7) |
| TIL pt 2 | 6.9 | 26.8 | 0.5 | 1.2 | −6.8 | 2.5 | 56 | + | + | − | 1.8 | 1/19 | extensive | 2 (SBR7) |
| TIL pt 4 | 0.9 | 23.5 | 0.9 | 0.3 | −8.5 | 1.5 | 42 | + | + | + | 4.4 | 7/26 | extensive | 3 (SBR9) |
| TIL pt 10 | 14.5 | 24.8 | −2.2 | 1.3 | −7.4 | 9.3 | 61 | − | − | − | 3.5 | 0/11 | extensive | 3 (SBR9) |
| TIL pt 7 | 5.4 | 22.8 | −1.3 | 0.4 | −9.0 | 5.6 | 54 | − | − | − | 1.8 | 0/13 | extensive | 3 (SBR9) |
| TIL pt 6 | 9.6 | 19.0 | −9.8 | −3.3 | −10.4 | 4.0 | 74 | − | − | − | 0.4 M | 0/14 | minimal | 3 (SBR8) |
| TIL pt 8 | 3.2 | 13.6 | −4.5 | −3.2 | −8.3 | 5.2 | 68 | − | − | − | 2.7 | 0/20 | minimal | 3 (SBR8) |
| TIL pt 9 | −1.9 | 18.1 | −3.4 | −4.2 | −14.2 | −1.3 | 82 | − | − | − | 2.5 M | 20/22 | minimal | 3 (SBR8) |
| a Estrogen receptor 1 (ESR1); estrogen receptor alpha b Progesterone receptor. c CD340, HER-2/neu, ERBB2. d For multi-focal tumors, the size of the largest invasive foci is indicated followed by M for multifocal. e The number of positive LNs/number of examined LNs. f The extent of T cell infiltrate determined by immunohistochemistry (IHC; CD3 and CD4 labeling) g Scarff Bloom-Richardson staging score Breast cancer subtypes are color coded according to IHC staining: | ||||||||||||||
| ER+ HER2− | HER2+ | ER− HER2− | ||||||||||||
| FOXO | NFκB | JAK-STAT1/2 | JAK-STAT3 | TGFβ | Notch | |
|---|---|---|---|---|---|---|
| patient 1 | 8.5 | 17.4 | −7.4 | −2.4 | −5.0 | 1.6 |
| patient 2 | −1.6 | −1.5 | −7.7 | −7.3 | −14.5 | −5.7 |
| patient 3 | 5.9 | 6.3 | −8.7 | −5.5 | −14.7 | −8.7 |
| patient 4 | −3.8 | −5.1 | −5.6 | −6.2 | −19.6 | −6.1 |
| patient 5 | 2.4 | 8.4 | −8.7 | −4.3 | −11.7 | −4.2 |
| patient 6 | 1.7 | 5.9 | −7.2 | −6.3 | −13.2 | −8.4 |
| patient 7 | 6.9 | 3.2 | −8.3 | −5.8 | −14.3 | −7.3 |
| patient 8 | 1.3 | 5.2 | −9.3 | −6.7 | −14.9 | −6.8 |
| patient 9 | −2.5 | −1.3 | −4.2 | −8.1 | −18.4 | −8.0 |
| patient 10 | −2.2 | −3.6 | −6.2 | −7.3 | −18.8 | −8.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wesseling-Rozendaal, Y.; van Doorn, A.; Willard-Gallo, K.; van de Stolpe, A. Characterization of Immunoactive and Immunotolerant CD4+ T Cells in Breast Cancer by Measuring Activity of Signaling Pathways That Determine Immune Cell Function. Cancers 2022, 14, 490. https://doi.org/10.3390/cancers14030490
Wesseling-Rozendaal Y, van Doorn A, Willard-Gallo K, van de Stolpe A. Characterization of Immunoactive and Immunotolerant CD4+ T Cells in Breast Cancer by Measuring Activity of Signaling Pathways That Determine Immune Cell Function. Cancers. 2022; 14(3):490. https://doi.org/10.3390/cancers14030490
Chicago/Turabian StyleWesseling-Rozendaal, Yvonne, Arie van Doorn, Karen Willard-Gallo, and Anja van de Stolpe. 2022. "Characterization of Immunoactive and Immunotolerant CD4+ T Cells in Breast Cancer by Measuring Activity of Signaling Pathways That Determine Immune Cell Function" Cancers 14, no. 3: 490. https://doi.org/10.3390/cancers14030490