

Polymer Kernels as Compact Carriers for Suspended Cardiomyocytes
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
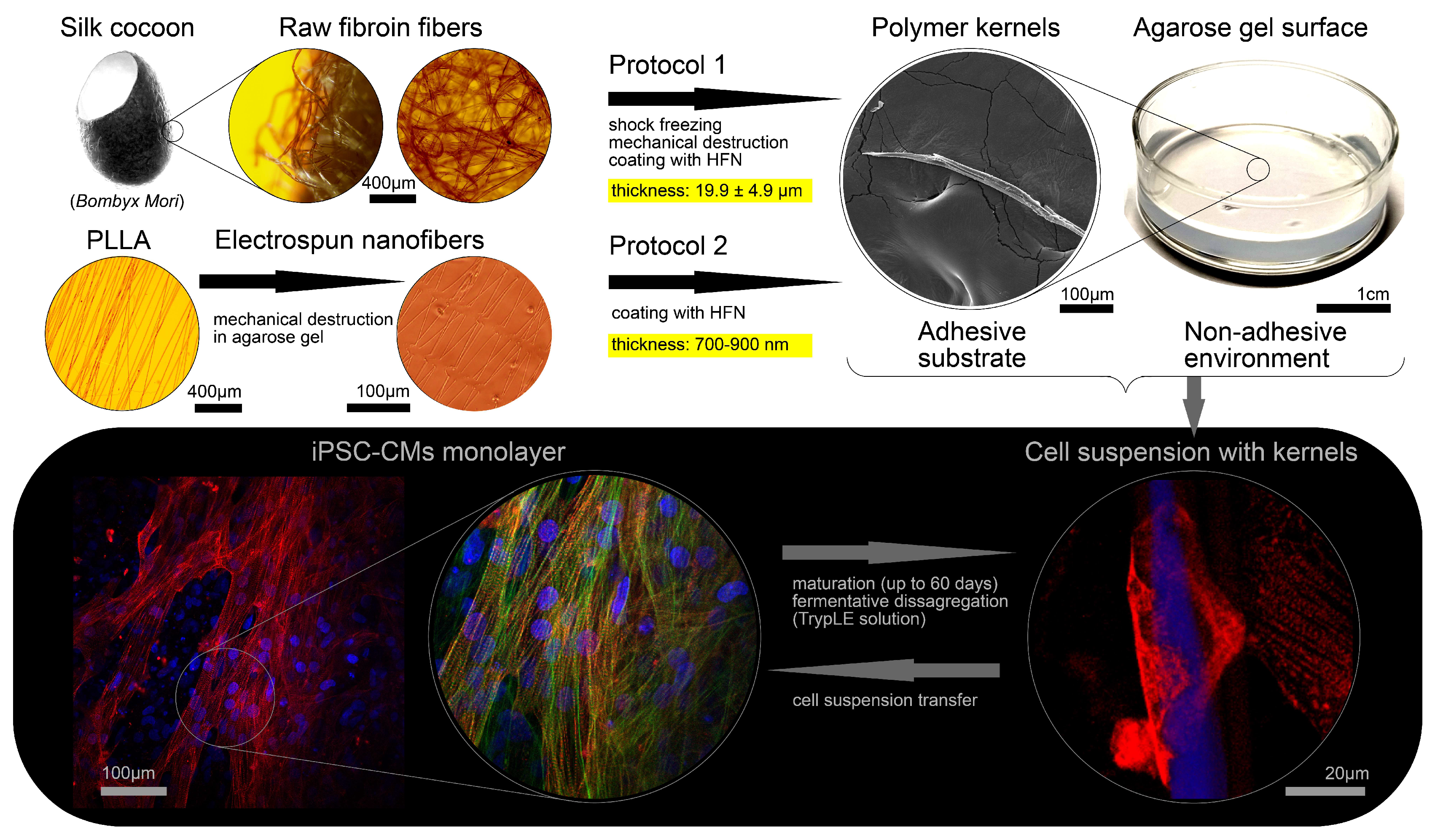
2.1. The Processing of Silkworm Cocoons and Cell Cultivation on Microcarriers (Protocol 1)
2.2. Electrospinning of Aligned Polymer Fibers (Protocol 2)
2.3. Cell Transfer Procedure
2.4. Optical Mapping Protocol
2.5. Immunofluorescent Staining
2.6. Data Processing and Statistical Analysis
2.7. Cell Culture and Ethical Approval
3. Results
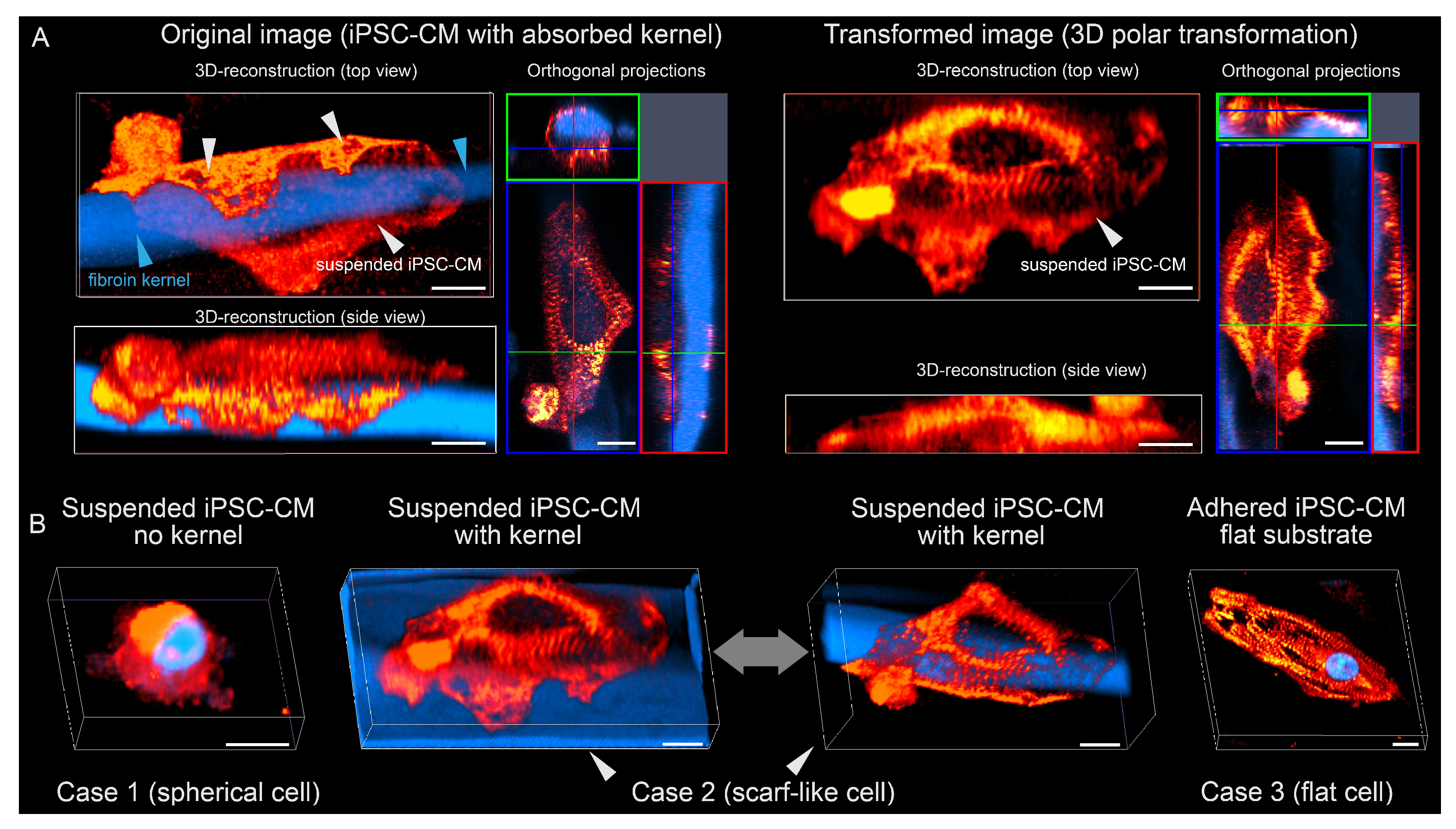
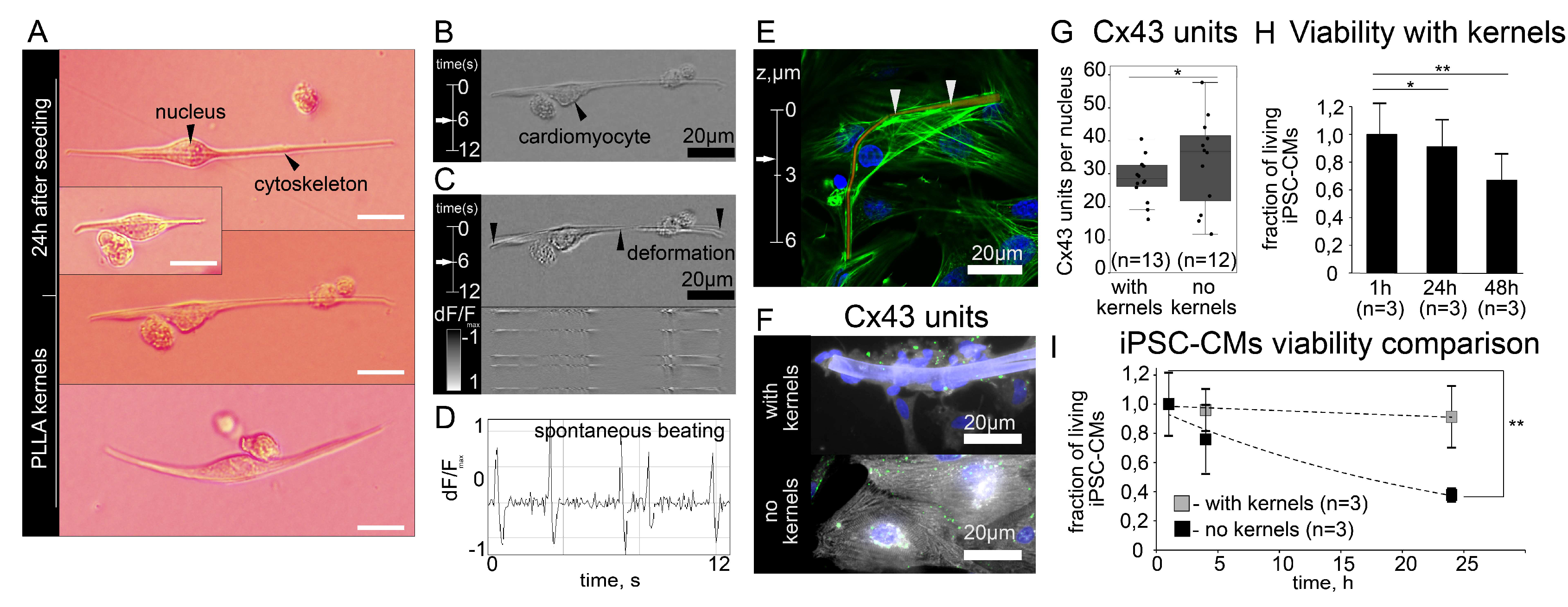
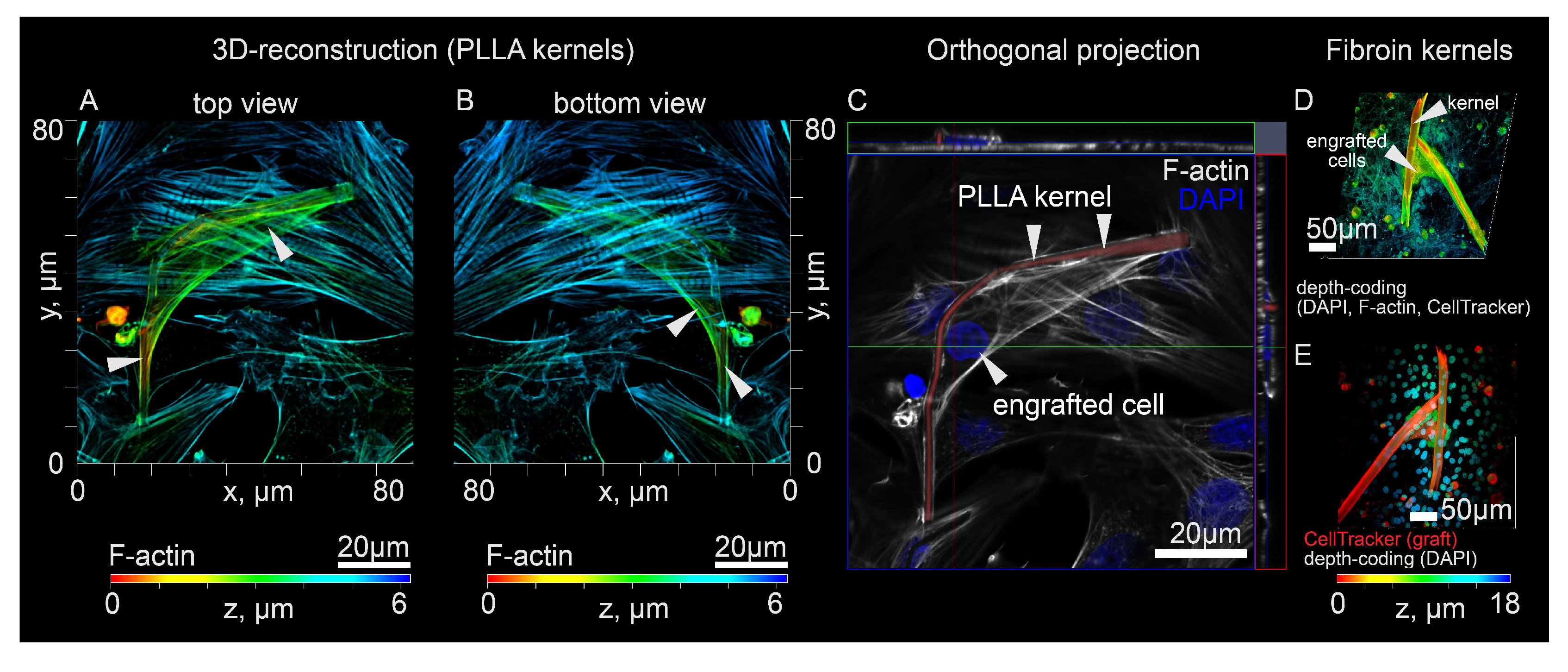
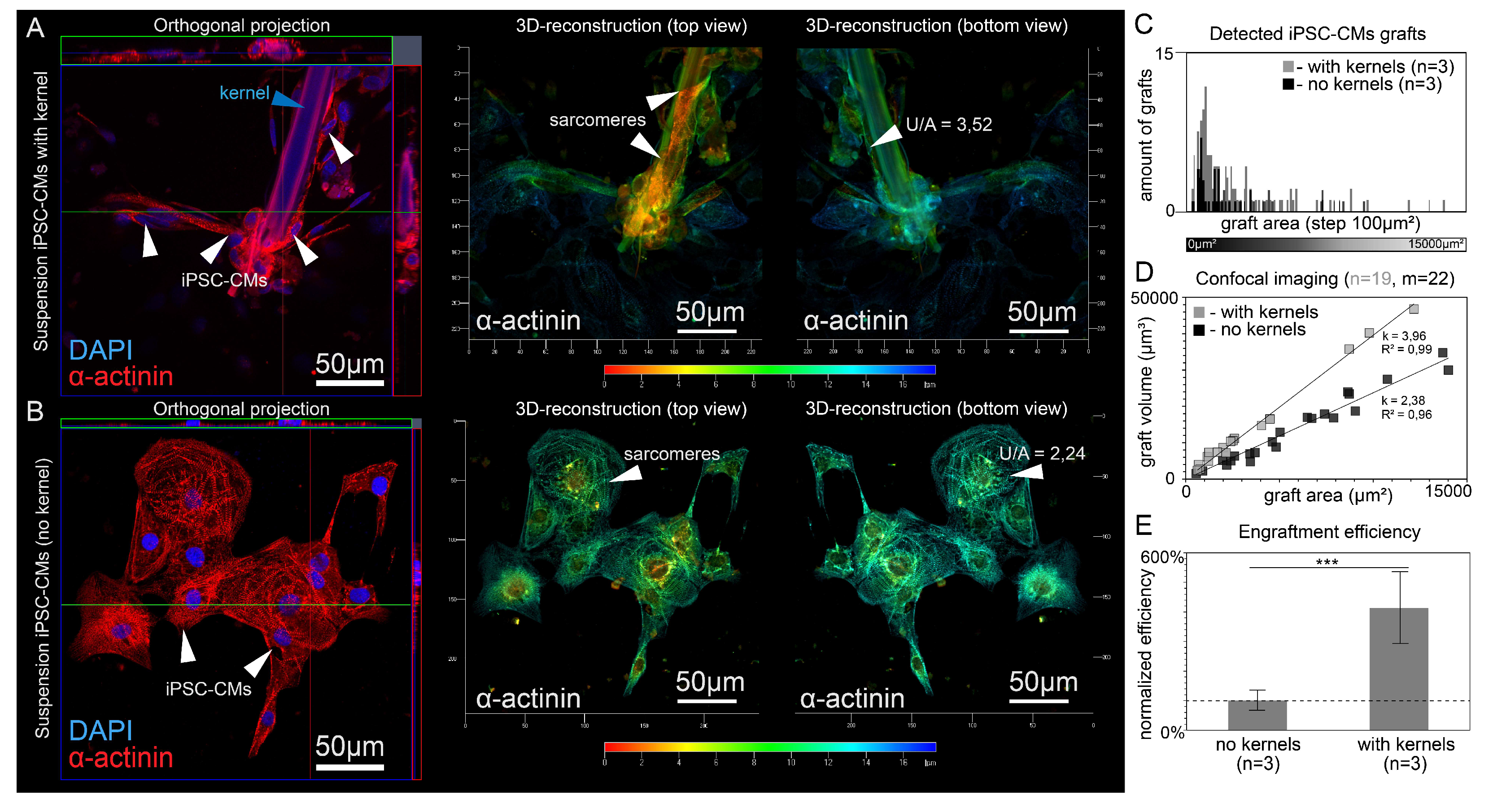
3.1. Polymer Kernels for Intracellular Organization Recovery
3.2. Co-Culturing of Excitable Cells on Polymer Kernels with a Monolayer of Cardiomyocytes
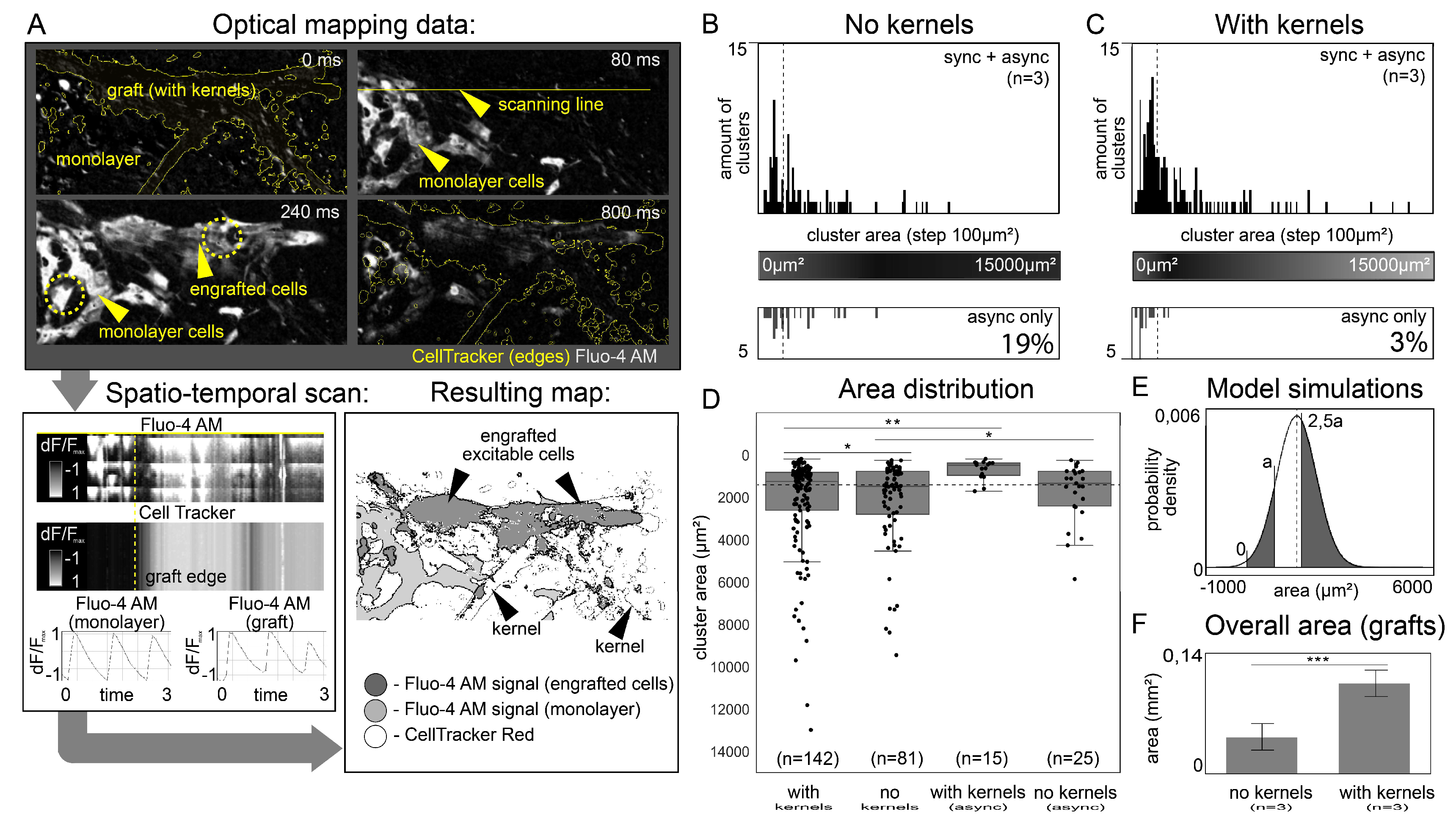
3.3. Evidence of Electrical Coupling between Isolated iPSC-CMs and Excitable Monolayer in Optical Mapping Experiments and Statistical Validation
3.4. Analysis of the Assumptions Underlying the Optical Mapping Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
References
- Kadota, S.; Shiba, Y. Pluripotent stem cell-derived cardiomyocyte transplantation for heart disease treatment. Curr. Cardiol. Rep. 2019, 21, 73. [Google Scholar] [CrossRef] [PubMed]
- Bizy, A.; Klos, M. Optimizing the use of iPSC-CMs for cardiac regeneration in animal models. Animals 2020, 10, 1561. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; Wada, Y.; Ichimura, H.; Tanaka, Y.; Ogasawara, T.; Okada, K.; Shiba, N.; Sakamoto, K.; et al. Allogeneic transplantation of iPS cell-derived cardiomyocytes regenerates primate hearts. Nature 2016, 538, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Kishino, Y.; Fujita, J.; Tohyama, S.; Okada, M.; Tanosaki, S.; Someya, S.; Fukuda, K. Toward the realization of cardiac regenerative medicine using pluripotent stem cells. Inflamm. Regen. 2020, 40, 1. [Google Scholar] [CrossRef] [PubMed]
- Romagnuolo, R.; Masoudpour, H.; Porta-Sánchez, A.; Qiang, B.; Barry, J.; Laskary, A.; Qi, X.; Massé, S.; Magtibay, K.; Kawajiri, H.; et al. Human Embryonic Stem Cell-Derived Cardiomyocytes Regenerate the Infarcted Pig Heart but Induce Ventricular Tachyarrhythmias. Stem Cell Rep. 2019, 12, 967–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, S.; Miki, K.; Takaki, T.; Okubo, C.; Hatani, T.; Chonabayashi, K.; Nishikawa, M.; Takei, I.; Oishi, A.; Narita, M.; et al. Enhanced engraftment, proliferation and therapeutic potential in heart using optimized human iPSC-derived cardiomyocytes. Sci. Rep. 2016, 6, 19111. [Google Scholar]
- Joseph, K.J.; Liang, A.; Weinberg, S.; Trayanova, N. Computational modeling of aberrant electrical activity following remuscularization with intramyocardially injected pluripotent stem cell-derived cardiomyocytes. J. Mol. Cell. Cardiol. 2022, 162, 97–109. [Google Scholar]
- Karakikes, I.; Ameen, M.; Termglinchan, V.; Wu, J. Human induced pluripotent stem cell–derived cardiomyocytes: Insights into molecular, cellular, and functional phenotypes. Circ. Res. 2015, 117, 80–88. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Ye, L. Maturation of pluripotent stem cell-derived cardiomyocytes: A critical step for drug development and cell therapy. J. Cardiovasc. Transl. Res. 2018, 11, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Kwon, C. Regulation of cardiomyocyte maturation during critical perinatal window. J. Physiol. 2020, 598, 2941–2956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, C.S.; Fukunishi, T.; Zhang, H.; Huang, C.Y.; Nashed, A.; Blazeski, A.; DiSilvestre, D.; Vricella, L.; Conte, J.; Tung, L.; et al. Biomaterial-free three-dimensional bioprinting of cardiac tissue using human induced pluripotent stem cell derived cardiomyocytes. Sci. Rep. 2017, 7, 4566. [Google Scholar] [CrossRef] [PubMed]
- Kawatou, M.; Masumoto, H.; Fukushima, H.; Morinaga, G.; Sakata, R.; Ashihara, T.; Yamashita, J.K. Modelling Torsade de Pointes arrhythmias in vitro in 3D human iPS cell-engineered heart tissue. Nat. Commun. 2017, 8, 1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slotvitsky, M.M.; Tsvelaya, V.A.; Podgurskaya, A.D.; Agladze, K.I. Formation of an electrical coupling between differentiating cardiomyocytes. Sci. Rep. 2020, 10, 7774. [Google Scholar] [CrossRef] [PubMed]
- Orlova, Y.; Magome, N.; Liu, L.; Chen, Y.; Agladze, K. Electrospun nanofibers as a tool for architecture control in engineered cardiac tissue. Biomaterials 2011, 32, 5615–5624. [Google Scholar] [CrossRef] [PubMed]
- Moisenovich, M.M.; Pustovalova, O.; Shackelford, J.; Vasiljeva, T.V.; Druzhinina, T.V.; Kamenchuk, Y.A.; Guzeev, V.V.; Sokolova, O.S.; Bogush, V.G.; Debabov, V.G.; et al. Tissue regeneration in vivo within recombinant spidroin 1 scaffolds. Biomaterials 2012, 33, 3887–3898. [Google Scholar] [CrossRef]
- Lowery, J.; Datta, N.; Rutledge, G. Effect of fiber diameter, pore size and seeding method on growth of human dermal fibroblasts in electrospun poly (ε-caprolactone) fibrous mats. Biomaterials 2010, 31, 491–504. [Google Scholar] [CrossRef]
- Blakeney, B.A.; Tambralli, A.; Anderson, J.M.; Andukuri, A.; Lim, D.-J.; Dean, D.R.; Jun, H.-W. Cell infiltration and growth in a low density, uncompressed three-dimensional electrospun nanofibrous scaffold. Biomaterials 2011, 32, 1583–1590. [Google Scholar] [CrossRef] [Green Version]
- Teplenin, A.; Krasheninnikova, A.; Agladze, N.; Sidoruk, K.; Agapova, O.; Agapov, I.; Bogush, V.; Agladze, K. Functional Analysis of the Engineered Cardiac Tissue Grown on Recombinant Spidroin Fiber Meshes. PLoS ONE 2015, 10, e0121155. [Google Scholar] [CrossRef]
- Shadrin, I.Y.; Allen, B.W.; Qian, Y.; Jackman, C.P.; Carlson, A.L.; Juhas, M.E.; Bursac, N. Cardiopatch platform enables maturation and scale-up of human pluripotent stem cell-derived engineered heart tissues. Nat. Commun. 2017, 8, 1825. [Google Scholar] [CrossRef] [Green Version]
- Balashov, V.; Efimov, A.; Agapova, O.; Pogorelov, A.; Agapov, I.; Agladze, K. High resolution 3D microscopy study of cardiomyocytes on polymer scaffold nanofibers reveals formation of unusual sheathed structure. Acta Biomater. 2018, 68, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Podgurskaya, A.D.; Tsvelaya, V.A.; Slotvitsky, M.M.; Dementyeva, E.V.; Valetdinova, K.R.; Agladze, K.I. The use of iPSC-derived cardiomyocytes and optical mapping for erythromycin arrhythmogenicity testing. Cardiovasc. Toxicol. 2019, 19, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Dementyeva, E.V.; Medvedev, S.P.; Kovalenko, V.R.; Vyatkin, Y.V.; Kretov, E.I.; Slotvitsky, M.M.; Shtokalo, D.N.; Pokushalov, E.A.; Zakian, S.M. Applying Patient-Specific Induced Pluripotent Stem Cells to Create a Model of Hypertrophic Cardiomyopathy. Biochemistry 2019, 84, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Podgurskaya, A.D.; Slotvitsky, M.M.; Tsvelaya, V.A.; Frolova, S.R.; Romanova, S.G.; Balashov, V.A.; Agladze, K.I. Cyclophosphamide arrhythmogenicitytesting using human-induced pluripotent stem cell-derived cardiomyocytes. Sci. Rep. 2021, 11, 2336. [Google Scholar] [CrossRef]
- Kudryashova, N.; Tsvelaya, V.; Agladze, K.; Panfilov, A. Virtual cardiac monolayers for electrical wave propagation. Sci. Rep. 2017, 7, 7887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudryashova, N.; Nizamieva, A.; Tsvelaya, V.; Panfilov, A.V.; Agladze, K.I. Self-organization of conducting pathways explains electrical wave propagation in cardiac tissues with high fraction of non-conducting cells. PLoS Comput. Biol. 2019, 15, e1006597. [Google Scholar] [CrossRef] [Green Version]
- Agladze, N.; Halaidych, O.; Tsvelaya, V.; Bruegmann, T.; Kilgus, C.; Sasse, P.; Agladze, K. Synchronization of excitable cardiac cultures of different origin. Biomater. Sci. 2017, 5, 1777–1785. [Google Scholar] [CrossRef]
- Sperelakis, N. An Electric Field Mechanism for Transmission of Excitation Between Myocardial Cells. Circ. Res. 2002, 91, 985–987. [Google Scholar] [CrossRef]
- Kléber, A.G.; Rudy, Y. Basic Mechanisms of Cardiac Impulse Propagation and Associated Arrhythmias. Physiol. Rev. 2004, 84, 431–488. [Google Scholar] [CrossRef] [Green Version]
- Kohl, P.; Gourdie, R. Fibroblast-myocyte electrotonic coupling: Does it occur in native cardiac tissue? J. Mol. Cell. Cardiol. 2014, 70, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Vepari, C.; Kaplan, D. Silk as a biomaterial. Prog. Polym. Sci. 2007, 32, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.-D.; Cheng, Y.; Teng, C.-P.; Khin, Y.-W.; Loh, X.-J.; Tee, S.-Y.; Low, M.; Ye, E.; Yu, H.-D.; Zhang, Y.-W.; et al. Structures, mechanical properties and applications of silk fibroin materials. Prog. Polym. Sci. 2015, 46, 86–110. [Google Scholar] [CrossRef]
- Herron, T.J.; Da Rocha, A.M.; Campbell, K.F.; Ponce-Balbuena, D.; Willis, C.; Guerrero-Serna, G.; Liu, Q.; Klos, M.; Musa, H.; Zarzoso, M.; et al. Extracellular Matrix–Mediated Maturation of Human Pluripotent Stem Cell–Derived Cardiac Monolayer Structure and Electrophysiological Function. Circ. Arrhythmia Electrophysiol. 2016, 9, e003638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCain, M.; Lee, H.; Aratyn-Schaus, Y.; Kléber, A.; Parker, K. Cooperative coupling of cell-matrix and cell–cell adhesions in cardiac muscle. Proc. Natl. Acad. Sci. USA 2012, 109, 9881–9886. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Neidig, L.E.; Yang, X.; Weber, G.J.; El-Nachef, D.; Tsuchida, H.; Dupras, S.; Kalucki, F.A.; Jayabalu, A.; Futakuchi-Tsuchida, A.; et al. Pharmacologic therapy for engraftment arrhythmia induced by transplantation of human cardiomyocytes. Stem Cell Rep. 2021, 16, 2473–2487. [Google Scholar] [CrossRef] [PubMed]
- Tapa, S.; Wang, L.; Stuart, S.D.F.; Wang, Z.; Jiang, Y.; Habecker, B.A.; Ripplinger, C.M. Adrenergic supersensitivity and impaired neural control of cardiac electrophysiology following regional cardiac sympathetic nerve loss. Sci. Rep. 2020, 10, 18801. [Google Scholar] [CrossRef]
- Paci, M.; Pölönen, R.-P.; Cori, D.; Penttinen, K.; Aalto-Setälä, K.; Severi, S.; Hyttinen, J. Automatic Optimization of an in silico Model of Human iPSC Derived Cardiomyocytes Recapitulating Calcium Handling Abnormalities. Front. Physiol. 2018, 9, 709. [Google Scholar] [CrossRef] [Green Version]
- Lesman, A.; Gepstein, L.; Levenberg, S. Cell Tri-Culture for Cardiac Vascularization. In Cardiac Tissue Engineering; Humana Press: New York, NY, USA, 2014; pp. 131–137. [Google Scholar]
- MacQueen, L.A.; Sheehy, S.P.; Chantre, C.O.; Zimmerman, J.F.; Pasqualini, F.S.; Liu, X.; Goss, J.A.; Campbell, P.H.; Gonzalez, G.M.; Park, S.-J.; et al. A tissue-engineered scale model of the heart ventricle. Nat. Biomed. Eng. 2018, 2, 930–941. [Google Scholar] [CrossRef]
- Entcheva, E.; Kay, M.W. Cardiac optogenetics: A decade of enlightenment. Nat. Rev. Cardiol. 2021, 18, 349–367. [Google Scholar] [CrossRef]
- Mitrea, B.G.; Caldwell, B.J.; Pertsov, A.M. Imaging electrical excitation inside the myocardial wall. Biomed. Opt. Express 2011, 2, 620–633. [Google Scholar] [CrossRef] [Green Version]
- Tarr, M.; Sperelakis, N. Weak electrotonic interaction between contiguous cardiac cells. Am. J. Physiol. 1964, 207, 691–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperelakis, N. Combined electric field and gap junctions on propagation of action potentials in cardiac muscle and smooth muscle in pspice simulation. J. Electrocardiol. 2003, 36, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Slotvitsky, M.; Tsvelaya, V.; Frolova, S.; Dementyeva, E.; Agladze, K. Arrhythmogenicity test based on a human-induced pluripotent stem cell (iPSC)-derived cardiomyocyte layer. Toxicol. Sci. 2019, 168, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Balashov, V.; Gorbunov, V.; Guria, K.; Agladze, K. Muscular thin films for label-free mapping of excitation propagation in cardiac tissue. Ann. Biomed. Eng. 2020, 48, 2425–2437. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slotvitsky, M.; Berezhnoy, A.; Scherbina, S.; Rimskaya, B.; Tsvelaya, V.; Balashov, V.; Efimov, A.E.; Agapov, I.; Agladze, K. Polymer Kernels as Compact Carriers for Suspended Cardiomyocytes. Micromachines 2023, 14, 51. https://doi.org/10.3390/mi14010051
Slotvitsky M, Berezhnoy A, Scherbina S, Rimskaya B, Tsvelaya V, Balashov V, Efimov AE, Agapov I, Agladze K. Polymer Kernels as Compact Carriers for Suspended Cardiomyocytes. Micromachines. 2023; 14(1):51. https://doi.org/10.3390/mi14010051
Chicago/Turabian StyleSlotvitsky, Mikhail, Andrey Berezhnoy, Serafima Scherbina, Beatrisa Rimskaya, Valerya Tsvelaya, Victor Balashov, Anton E. Efimov, Igor Agapov, and Konstantin Agladze. 2023. "Polymer Kernels as Compact Carriers for Suspended Cardiomyocytes" Micromachines 14, no. 1: 51. https://doi.org/10.3390/mi14010051