Sensitive Detection of Ciguatoxins Using a Neuroblastoma Cell-Based Assay with Voltage-Gated Potassium Channel Inhibitors


Abstract
:1. Introduction
2. Results
2.1. Optimization of O, V, and KV Channel Inhibitors’ (4-AP and TEA-Cl) Concentrations
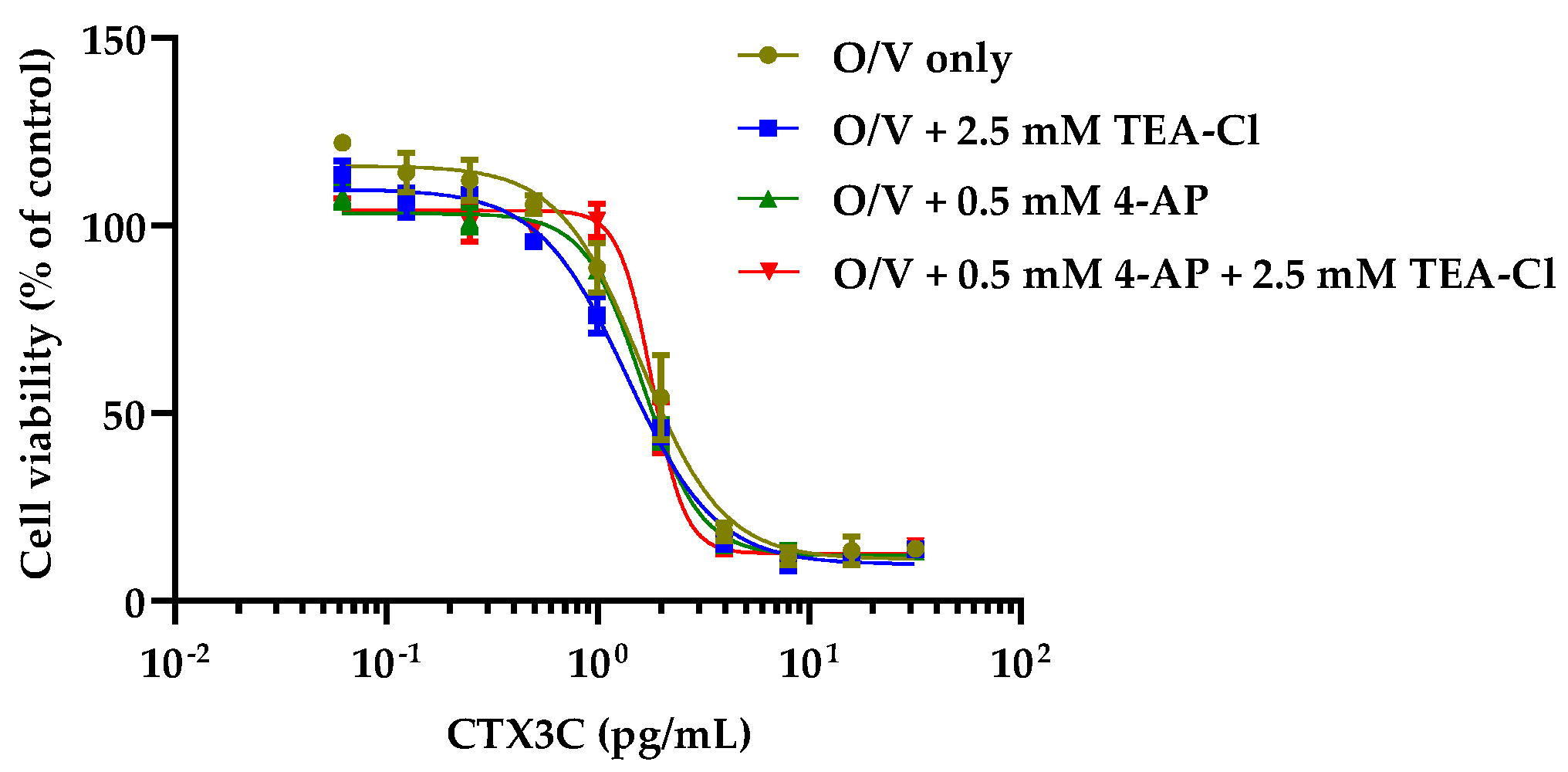
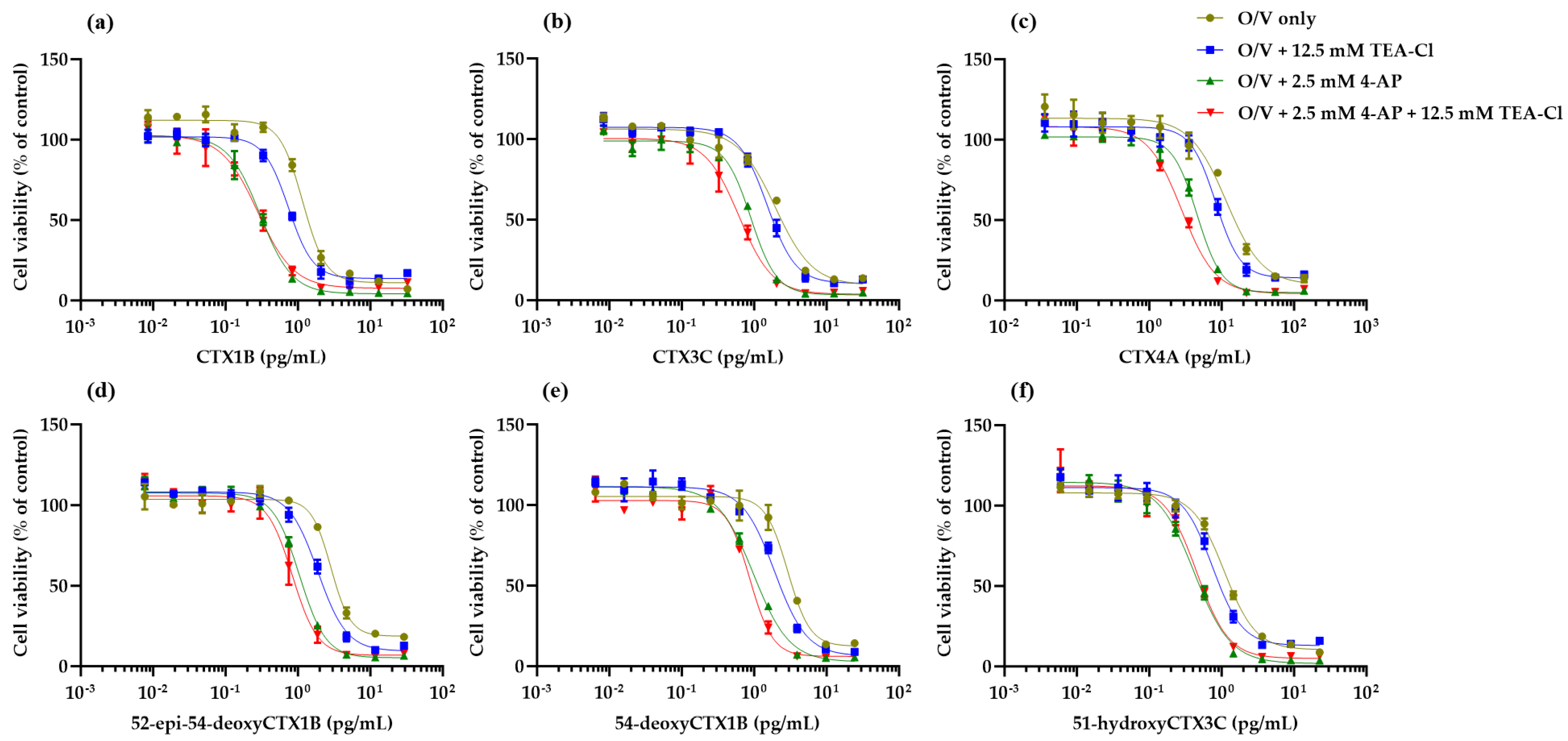
2.2. Cytotoxicity of CTX3C against N2a Cells in the Presence of KV Channel Inhibitors
2.3. Cytotoxicity of CTX Congeners against N2a Cells in the Coexistence of O, V, and KV Channel Inhibitors
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Reference CTXs
5.3. N2a Assay Using KV Channel Inhibitors
5.3.1. Passaging of the Cell Line
5.3.2. Optimization of O, V, and KV Channel Inhibitors’ (4-AP and TEA-Cl) Concentrations
5.3.3. Cytotoxicity of CTX Congeners against N2a Cells in the Presence of KV Channel Inhibitors and the Coexistence of O, V, and KV Channel Inhibitors
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.M.; Yasumoto, T. Isolation and structure of ciguatoxin-4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem. 1996, 60, 2103–2105. [Google Scholar] [CrossRef] [PubMed]
- Laza-Martínez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a strain of Fukuyoa Paulensis (Dinophyceae) from the western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef] [PubMed]
- Leite, I.D.P.; Sdiri, K.; Taylor, A.; Viallon, J.; Gharbia, H.B.; Mafra Júnior, L.L.; Swarzenski, P.; Oberhaensli, F.; Darius, H.T.; Chinain, M.; et al. Experimental evidence of ciguatoxin accumulation and depuration in carnivorous lionfish. Toxins 2021, 13, 564. [Google Scholar] [CrossRef] [PubMed]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A public health perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Revel, T.; Cruchet, P.; Viallon, J.; Gatti, C.M.I.; Sibat, M.; Hess, P.; Chinain, M. Deep-Water fish are potential vectors of ciguatera poisoning in the Gambier Islands, French Polynesia. Mar. Drugs 2021, 19, 644. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Picot, S.; Ung, A.; Viallon, J.; Gaertner-Mazouni, N.; Sibat, M.; Amzil, Z.; Chinain, M. Evidence of the bioaccumulation of ciguatoxins in giant clams (Tridacna maxima) exposed to Gambierdiscus spp. cells. Harmful Algae 2016, 57, 78–87. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.I.I.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological investigations on the sea urchin Tripneustes gratilla (Toxopneustidae, echinoid) from Anaho bay (Nuku Hiva, French Polynesia): Evidence for the presence of Pacific ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.i.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Tectus niloticus (Tegulidae, gastropod) as a novel vector of ciguatera poisoning: Detection of Pacific ciguatoxins in toxic samples from Nuku Hiva island (French Polynesia). Toxins 2017, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Kibler, S.R.; Tester, P.A.; Kunkel, K.E.; Moore, S.K.; Litaker, R.W. Effects of ocean warming on growth and distribution of dinoflagellates associated with ciguatera fish poisoning in the Caribbean. Ecol. Model. 2015, 316, 194–210. [Google Scholar] [CrossRef]
- Costa, P.R.; Estévez, P.; Soliño, L.; Castro, D.; Rodrigues, S.M.; Timoteo, V.; Leao-Martins, J.M.; Santos, C.; Gouveia, N.; Diogène, J.; et al. An update on ciguatoxins and CTX-like toxicity in fish from different trophic levels of the Selvagens islands (NE Atlantic, Madeira, Portugal). Toxins 2021, 13, 580. [Google Scholar] [CrossRef]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; de Falco, G.; Campàs, M.; Diogène, J. Assessment of cytotoxicity in ten strains of Gambierdiscus australes from Macaronesian Islands by Neuro-2a cell-based assays. J. Appl. Phycol. 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Epelboin, L.; Pérignon, A.; Hossen, V.; Vincent, R.; Krys, S.; Caumes, E. Two clusters of ciguatera fish poisoning in Paris, France, related to tropical fish imported from the French Caribbean by travelers. J. Travel Med. 2014, 21, 397–402. [Google Scholar] [CrossRef]
- Friedemann, M. Ciguatera fish poisoning outbreaks from 2012 to 2017 in Germany caused by snappers from India, Indonesia, and Vietnam. J. Consum. Prot. Food Saf. 2019, 14, 71–80. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Spielmeyer, A.; Blaschke, V.; Bodi, D.; Kappenstein, O. Ciguatera poisoning in Europe: A traceback to Indian Ocean sourced snapper fish (Lutjanus Bohar). Food Control 2023, 151, 109799. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Spielmeyer, A.; Friedemann, M.; Kapp, K.; Schwank, U.; Kappenstein, O.; Bodi, D. Food safety risk in Germany from mislabeled imported fish: Ciguatera outbreak trace-back, toxin elucidation, and public health implications. Front. Mar. Sci. 2022, 9, 849857. [Google Scholar] [CrossRef]
- Tartaglione, L.; Loeffler, C.R.; Miele, V.; Varriale, F.; Varra, M.; Monti, M.; Varone, A.; Bodi, D.; Spielmeyer, A.; Capellacci, S.; et al. Dereplication of Gambierdiscus balechii extract by LC-HRMS and in vitro assay: First description of a putative ciguatoxin and confirmation of 44-Methylgambierone. Chemosphere 2023, 319, 137940. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Loeffler, C.R.; Kappenstein, O. Identical Ciguatoxin-3C group profiles in Lutjanus Bohar from the Pacific and Indian Oceans–Indicating the need to re-evaluate geographical CTX classifications. Front. Mar. Sci. 2022, 9, 937438. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of ciguatoxins leads to species-specific toxin profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef]
- Li, J.; Mak, Y.L.; Chang, Y.H.; Xiao, C.; Chen, Y.M.; Shen, J.; Wang, Q.; Ruan, Y.; Lam, P.K.S. Uptake and depuration kinetics of Pacific ciguatoxins in orange-spotted grouper (Epinephelus coioides). Environ. Sci. Technol. 2020, 54, 4475–4483. [Google Scholar] [CrossRef]
- Yasumoto, T. The chemistry and biological function of natural marine toxins. Chem. Rec. 2001, 1, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Vernoux, J.-P.; Brereton, I.M. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Estevez, P.; Oses Prieto, J.; Burlingame, A.; Gago Martinez, A. Characterization of the ciguatoxin profile in fish samples from the eastern Atlantic Ocean using capillary liquid chromatography-high resolution mass spectrometry. Food Chem. 2023, 418, 135960. [Google Scholar] [CrossRef]
- Kryuchkov, F.; Robertson, A.; Miles, C.O.; Mudge, E.M.; Uhlig, S. LC–HRMS and chemical derivatization strategies for the structure elucidation of Caribbean ciguatoxins: Identification of C-CTX-3 and -4. Mar. Drugs 2020, 18, 182. [Google Scholar] [CrossRef]
- Mudge, E.M.; Miles, C.O.; Ivanova, L.; Uhlig, S.; James, K.S.; Erdner, D.L.; Fæste, C.K.; McCarron, P.; Robertson, A. Algal ciguatoxin identified as source of ciguatera poisoning in the Caribbean. Chemosphere 2023, 330, 138659. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Diogène, J.; Reverté, L.; Rambla-Alegre, M.; Del Rió, V.; De La Iglesia, P.; Campàs, M.; Palacios, O.; Flores, C.; Caixach, J.; Ralijaona, C.; et al. Identification of ciguatoxins in a shark involved in a fatal food poisoning in the Indian Ocean. Sci. Rep. 2017, 7, 8240. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Handbook Fish and Fishery Products Hazard and Control Guidance June 2022 Edition; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2022. [Google Scholar]
- European Commission. Commission regulation (EU) No 15/2011 of 10 January 2011 Amending Regulation (EC) No 2074/2005 as Regards Recognised Testing Methods for Detecting Marine Biotoxins in Live Bivalve Molluscs. Off. J. Eur. Union 2011, L6, 3–6. [Google Scholar]
- Dechraoui, M.Y.B.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.R.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- Nagae, M.; Igarashi, T.; Mizukoshi, K.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Development and validation of an LC-MS/MS method for the ultra-trace analysis of Pacific ciguatoxins in fish. J. AOAC Int. 2021, 104, 1272–1281. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Sato, T.; Hirama, M.; Fujii, I. Highly sensitive and practical fluorescent sandwich ELISA for ciguatoxins. Anal. Chem. 2018, 90, 7318–7324. [Google Scholar] [CrossRef]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.F.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016, 11, e0153348. [Google Scholar] [CrossRef] [PubMed]
- Yokozeki, T.; Hama, Y.; Fujita, K.; Igarashi, T.; Hirama, M.; Tsumuraya, T. Evaluation of relative potency of calibrated ciguatoxin congeners by near-infrared fluorescent receptor binding and neuroblastoma cell-based assays. Toxicon 2023, 230, 107161. [Google Scholar] [CrossRef]
- Díaz-Asencio, L.; Clausing, R.J.; Rañada, M.L.; Alonso-Hernández, C.M.; Dechraoui Bottein, M.Y. A radioligand receptor binding assay for ciguatoxin monitoring in environmental samples: Method development and determination of quality control criteria. J. Environ. Radioact. 2018, 192, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the neuroblastoma cell-based assay (CBA-N2a) for the improved detection of marine toxins active on voltage gated sodium channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Bodi, D.; Tartaglione, L.; Dell’Aversano, C.; Preiss-Weigert, A. Improving in vitro ciguatoxin and brevetoxin detection: Selecting neuroblastoma (Neuro-2a) cells with lower sensitivity to ouabain and veratridine (OV-LS). Harmful Algae 2021, 103, 101994. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, S.; Gaiani, G.; Tsumuraya, T.; Hirama, M.; Turquet, J.; Sagristà, N.; Rambla-Alegre, M.; Flores, C.; Caixach, J.; Diogène, J.; et al. Addressing the analytical challenges for the detection of ciguatoxins using an electrochemical biosensor. Anal. Chem. 2020, 92, 4858–4865. [Google Scholar] [CrossRef]
- Alkassar, M.; Leonardo, S.; Diogène, J.; Campàs, M. Immobilisation of Neuro-2a cells on electrodes and electrochemical detection of MTT formazan crystals to assess their viability. Bioelectrochemistry 2022, 148, 108274. [Google Scholar] [CrossRef]
- Gaiani, G.; Leonardo, S.; Tudó, À.; Toldrà, A.; Rey, M.; Andree, K.B.; Tsumuraya, T.; Hirama, M.; Diogène, J.; O’Sullivan, C.K.; et al. Rapid detection of ciguatoxins in Gambierdiscus and Fukuyoa with immunosensing tools. Ecotoxicol. Environ. Saf. 2020, 204, 111004. [Google Scholar] [CrossRef] [PubMed]
- Gaiani, G.; Cucchi, F.; Toldrà, A.; Andree, K.B.; Rey, M.; Tsumuraya, T.; O’Sullivan, C.K.; Diogène, J.; Campàs, M. Electrochemical biosensor for the dual detection of Gambierdiscus australes and Gambierdiscus excentricus in field samples. First report of G. excentricus in the Balearic Islands. Sci. Total. Environ. 2022, 806, 150915. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on marine biotoxins in shellfish–Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8, 1627. [Google Scholar] [CrossRef]
- Darius, H.T.; Revel, T.; Viallon, J.; Sibat, M.; Cruchet, P.; Longo, S.; Hardison, D.R.; Holland, W.C.; Tester, P.A.; Litaker, R.W.; et al. Comparative study on the performance of three detection methods for the quantification of Pacific ciguatoxins in French Polynesian strains of Gambierdiscus polynesiensis. Mar. Drugs 2022, 20, 348. [Google Scholar] [CrossRef]
- Bidard, J.N.; Vijverberg, H.P.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem. 1984, 259, 8353–8357. [Google Scholar] [CrossRef] [PubMed]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of sodium channel toxins: Directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [CrossRef]
- Yasumoto, T.; Fukui, M.; Sasaki, K.; Sugiyama, K. Determinations of marine toxins in foods. J. AOAC Int. 1995, 78, 574–582. [Google Scholar] [CrossRef]
- Nicolas, J.; Hendriksen, P.J.M.; Gerssen, A.; Bovee, T.F.H.; Rietjens, I.M.C.M. Marine neurotoxins: State of the art, bottlenecks, and perspectives for mode of action based methods of detection in seafood. Mol. Nutr. Food Res. 2014, 58, 87–100. [Google Scholar] [CrossRef]
- Castro, D.; Manger, R.; Vilariño, O.; Gago-Martínez, A. Evaluation of matrix issues in the applicability of the Neuro-2a cell based assay on the detection of CTX in fish samples. Toxins 2020, 12, 308. [Google Scholar] [CrossRef]
- Campàs, M.; Leonardo, S.; Rambla-Alegre, M.; Sagristà, N.; Vaya, R.; Diogène, J.; Torréns, M.; Fragoso, A. Cyclodextrin polymer clean-up method for the detection of ciguatoxins in fish with cell-based assays. Food Chem. 2023, 401, 134196. [Google Scholar] [CrossRef]
- Hidalgo, J.; Liberona, J.L.; Molgó, J.; Jaimovich, E. Pacific Ciguatoxin-1b effect over Na+ and K+ currents, inositol 1,4,5-triphosphate content and intracellular Ca2+ signals in cultured rat myotubes. Br. J. Pharmacol. 2002, 137, 1055–1062. [Google Scholar] [CrossRef]
- Birinyi-Strachan, L.C.; Gunning, S.J.; Lewis, R.J.; Nicholson, G.M. Block of voltage-gated potassium channels by Pacific Ciguatoxin-1 contributes to increased neuronal excitability in rat sensory neurons. Toxicol. Appl. Pharmacol. 2005, 204, 175–186. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic polyether modulators of voltage-gated Iion Channel function. Mar. Drugs 2006, 4, 82–118. [Google Scholar] [CrossRef]
- Hossen, V.; Soliño, L.; Leroy, P.; David, E.; Velge, P.; Dragacci, S.; Krys, S.; Flores Quintana, H.; Diogène, J. Contribution to the risk characterization of ciguatoxins: LOAEL estimated from eight ciguatera fish poisoning events in Guadeloupe (French West Indies). Environ. Res. 2015, 143, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.; Murray, S.A.; Zammit, A.; Edwards, A.W. Management of ciguatoxin risk in Eastern Australia. Toxins 2017, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Raposo-Garcia, S.; Cao, A.; Costas, C.; Louzao, M.C.; Vilariño, N.; Vale, C.; Botana, L.M. Mouse N2a Neuroblastoma Assay: Uncertainties and Comparison with Alternative Cell-Based Assays for ciguatoxin Detection. Mar. Drugs 2023, 21, 590. [Google Scholar] [CrossRef] [PubMed]
- LePage, K.T.; Dickey, R.W.; Gerwick, W.H.; Jester, E.L.; Murray, T.F. On the use of Neuro-2a neuroblastoma cells versus intact neurons in primary culture for neurotoxicity studies. Crit. Rev. Neurobiol. 2005, 17, 27–50. [Google Scholar] [CrossRef] [PubMed]
- Berman, F.W.; Murray, T.F. Brevetoxin-induced autocrine excitotoxicity is associated with manifold routes of Ca2+ influx. J. Neurochem. 2000, 74, 1443–1451. [Google Scholar] [CrossRef]
- Alonso, E.; Vale, C.; Sasaki, M.; Fuwa, H.; Konno, Y.; Perez, S.; Vieytes, M.R.; Botana, L.M. Calcium oscillations induced by gambierol in cerebellar granule cells. J. Cell. Biochem. 2010, 110, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Raposo-Garcia, S.; Costas, C.; Louzao, M.C.; Vale, C.; Botana, L.M. Synergistic effect of environmental food pollutants: Pesticides and marine biotoxins. Sci. Total Environ. 2023, 858, 160111. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol: A new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
- Alonso, E.; Fuwa, H.; Vale, C.; Suga, Y.; Goto, T.; Konno, Y.; Sasaki, M.; LaFerla, F.M.; Vieytes, M.R.; Giménez-Llort, L.; et al. Design and synthesis of skeletal analogues of gambierol: Attenuation of amyloid-β and tau pathology with voltage-gated potassium channel and N-Methyl-d-Aspartate receptor implications. J. Am. Chem. Soc. 2012, 134, 7467–7479. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Cui, Y.; Busse, E.; Mehrotra, S.; Rainier, J.D.; Murray, T.F. Gambierol inhibition of voltage-gated potassium channels augments spontaneous Ca2+ oscillations in cerebrocortical neurons. J. Pharmacol. Exp. Ther. 2014, 350, 615–623. [Google Scholar] [CrossRef]
- Ito, E.; Suzuki-Toyota, F.; Toshimori, K.; Fuwa, H.; Tachibana, K.; Satake, M.; Sasaki, M. Pathological effects on mice by gambierol, possibly one of the ciguatera toxins. Toxicon 2003, 42, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Murata, M.; Torigoe, K.; Satake, M.; Yasumoto, T. Gambieric acids, new potent antifungal substances with unprecedented polyether structures from a marine dinoflagellate Gambierdiscus toxicus. J. Org. Chem. 1992, 57, 5448–5453. [Google Scholar] [CrossRef]
- Tudó, À.; Rambla-Alegre, M.; Flores, C.; Sagristà, N.; Aguayo, P.; Reverté, L.; Campàs, M.; Gouveia, N.; Santos, C.; Andree, K.B.; et al. Identification of new CTX analogues in fish from the Madeira and Selvagens archipelagos by Neuro-2a CBA and LC-HRMS. Mar. Drugs 2022, 20, 236. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Yasumoto, T. Quantification of representative ciguatoxins in the Pacific using quantitative nuclear magnetic resonance spectroscopy. Mar. Drugs 2017, 15, 309. [Google Scholar] [CrossRef]
- Yamashita, S.; Takeuchi, K.; Koyama, T.; Inoue, M.; Hayashi, Y.; Hirama, M. Practical route to the left wing of CTX1B and total syntheses of CTX1B and 54-deoxyCTX1B. Chemistry 2015, 21, 2621–2628. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Congeners | O/V Only (pg/mL) | O/V + TEA-Cl (pg/mL) | O/V + 4-AP (pg/mL) | O/V + 4-AP + TEA-Cl (pg/mL) |
|---|---|---|---|---|
| CTX1B | 1.16 ± 0.09 | 0.74 ± 0.05 *** | 0.30 ± 0.03 *** | 0.28 ± 0.04 *** |
| CTX3C | 1.96 ± 0.28 | 1.52 ± 0.11 * | 0.90 ± 0.07 *** | 0.62 ± 0.07 *** |
| CTX4A | 11.7 ± 1.88 | 8.27 ± 0.78 ** | 4.59 ± 0.27 *** | 2.82 ± 0.26 *** |
| 52-epi-54-deoxyCTX1B | 2.86 ± 0.23 | 1.88 ± 0.15 *** | 1.05 ± 0.06 *** | 0.84 ± 0.10 *** |
| 54-deoxyCTX1B | 2.91 ± 0.29 | 1.91 ± 0.20 *** | 0.98 ± 0.07 *** | 0.87 ± 0.09 *** |
| 51-hydroxyCTX3C | 1.09 ± 0.07 | 0.75 ± 0.08 *** | 0.42 ± 0.04 *** | 0.45 ± 0.06 *** |
| Congeners | O/V Only | O/V + TEA-Cl | O/V + 4-AP | O/V + 4-AP + TEA-Cl | TEFs [44] |
|---|---|---|---|---|---|
| CTX1B | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| CTX3C | 0.6 | 0.5 | 0.3 | 0.5 | 0.2 |
| CTX4A | 0.10 | 0.09 | 0.07 | 0.10 | 0.1 |
| 52-epi-54-deoxyCTX1B | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 |
| 54-deoxyCTX1B | 0.4 | 0.4 | 0.3 | 0.3 | 0.3 |
| 51-hydroxyCTX3C | 1.1 | 1.0 | 0.7 | 0.6 | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yokozeki, T.; Kawabata, M.; Fujita, K.; Hirama, M.; Tsumuraya, T. Sensitive Detection of Ciguatoxins Using a Neuroblastoma Cell-Based Assay with Voltage-Gated Potassium Channel Inhibitors. Toxins 2024, 16, 118. https://doi.org/10.3390/toxins16030118
Yokozeki T, Kawabata M, Fujita K, Hirama M, Tsumuraya T. Sensitive Detection of Ciguatoxins Using a Neuroblastoma Cell-Based Assay with Voltage-Gated Potassium Channel Inhibitors. Toxins. 2024; 16(3):118. https://doi.org/10.3390/toxins16030118
Chicago/Turabian StyleYokozeki, Toshiaki, Madoka Kawabata, Kazuhiro Fujita, Masahiro Hirama, and Takeshi Tsumuraya. 2024. "Sensitive Detection of Ciguatoxins Using a Neuroblastoma Cell-Based Assay with Voltage-Gated Potassium Channel Inhibitors" Toxins 16, no. 3: 118. https://doi.org/10.3390/toxins16030118