The Mycotoxin De-Epoxy-Deoxynivalenol (DOM-1) Increases Endoplasmic Reticulum Stress in Ovarian Theca Cells

Abstract
:1. Introduction
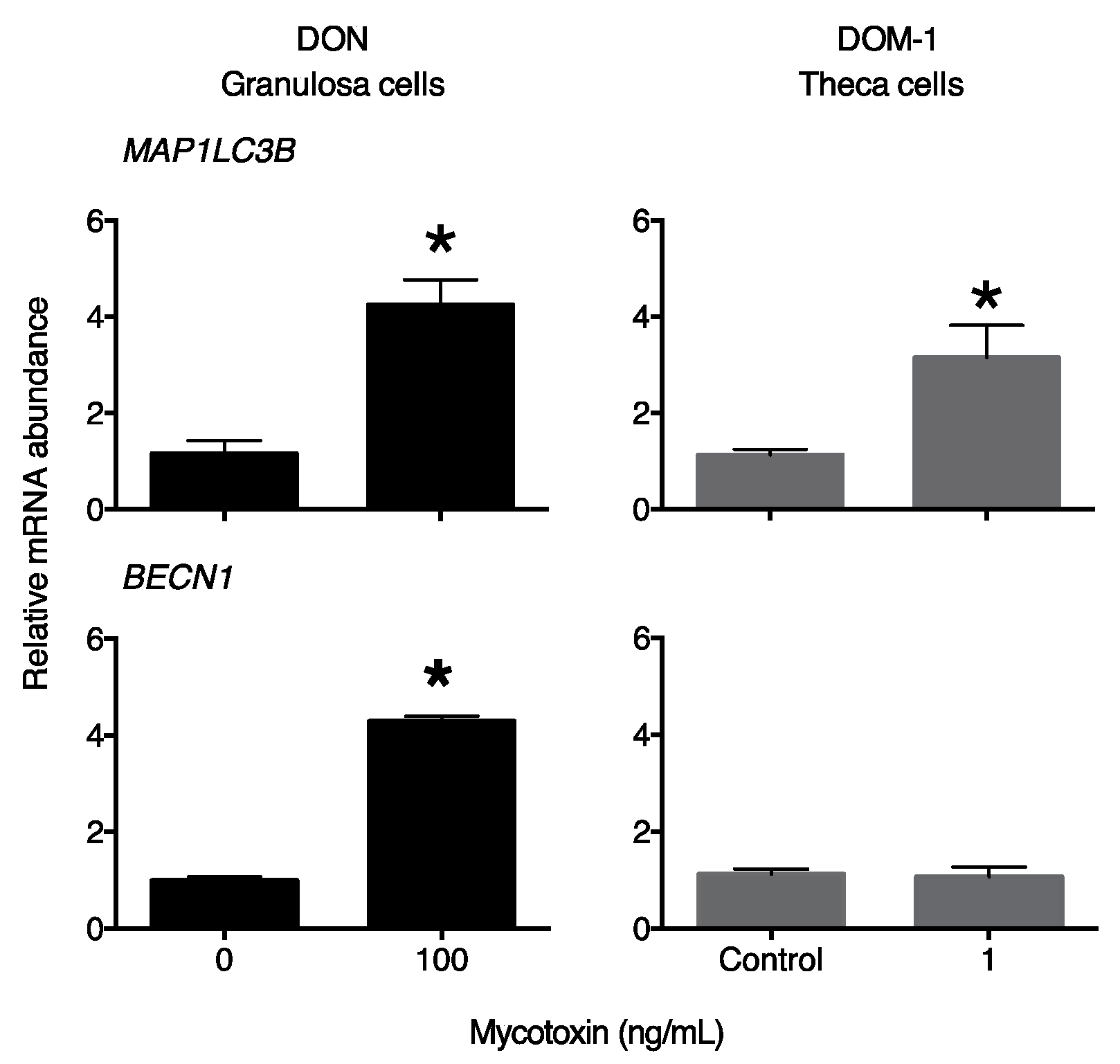
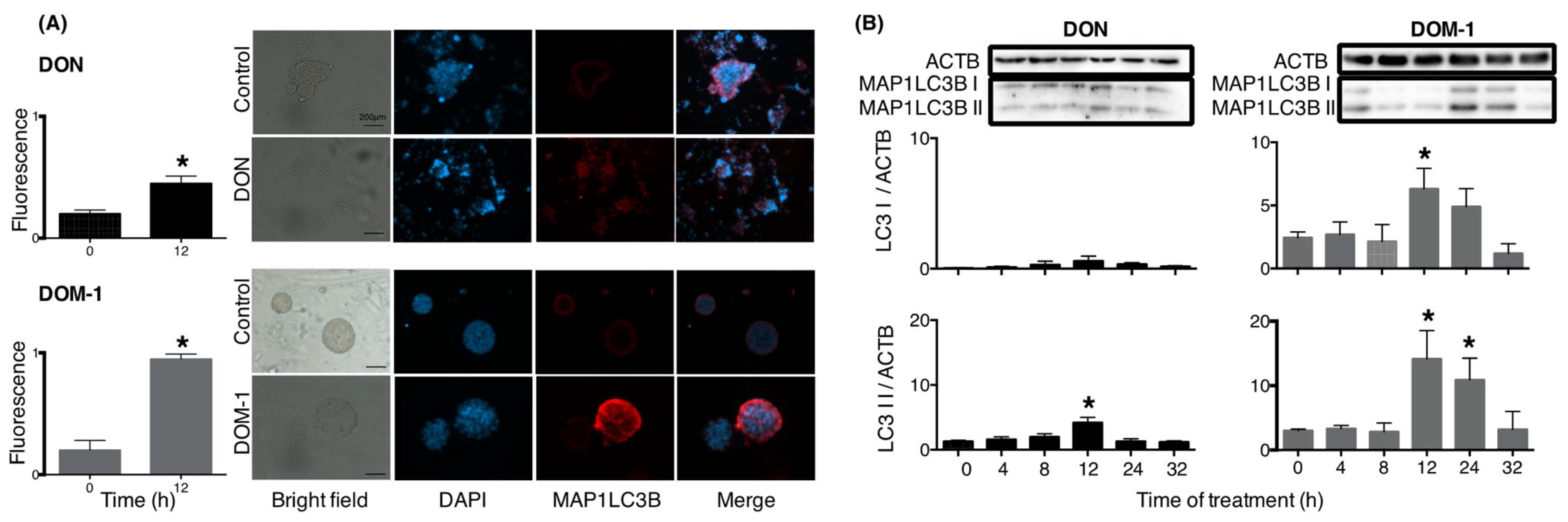
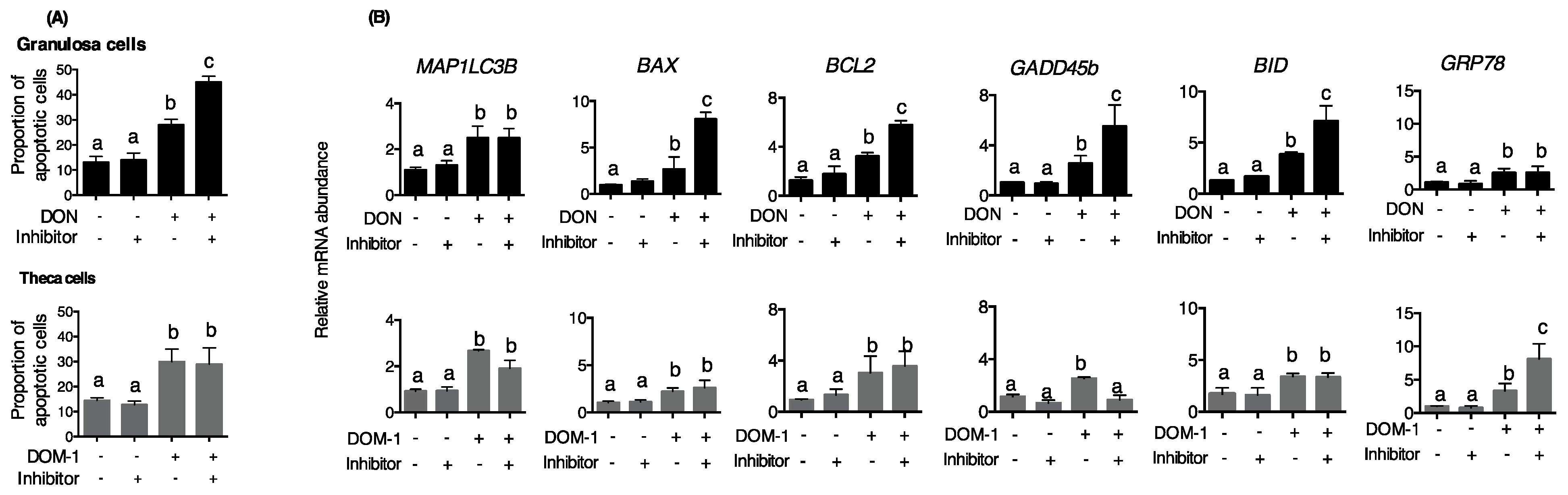
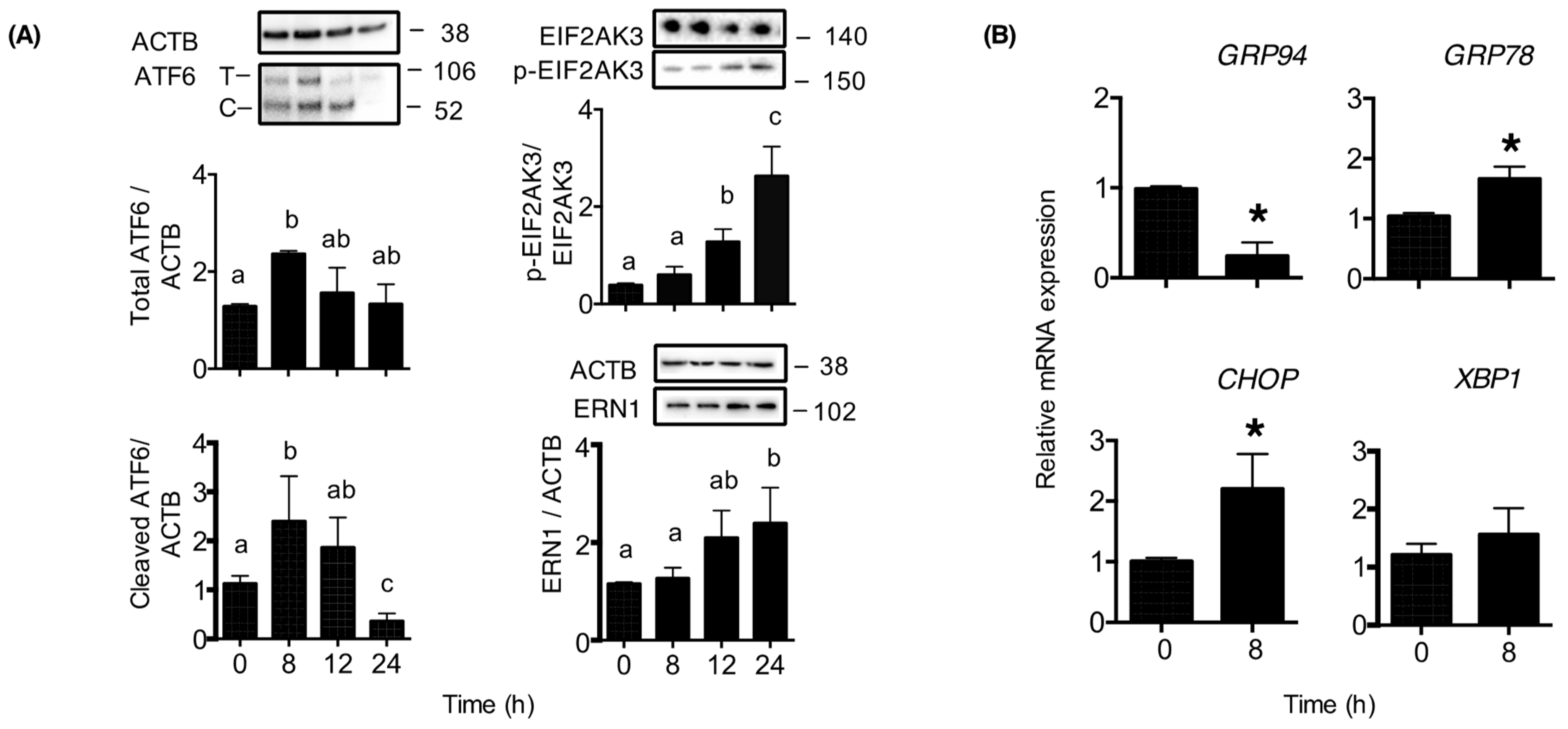
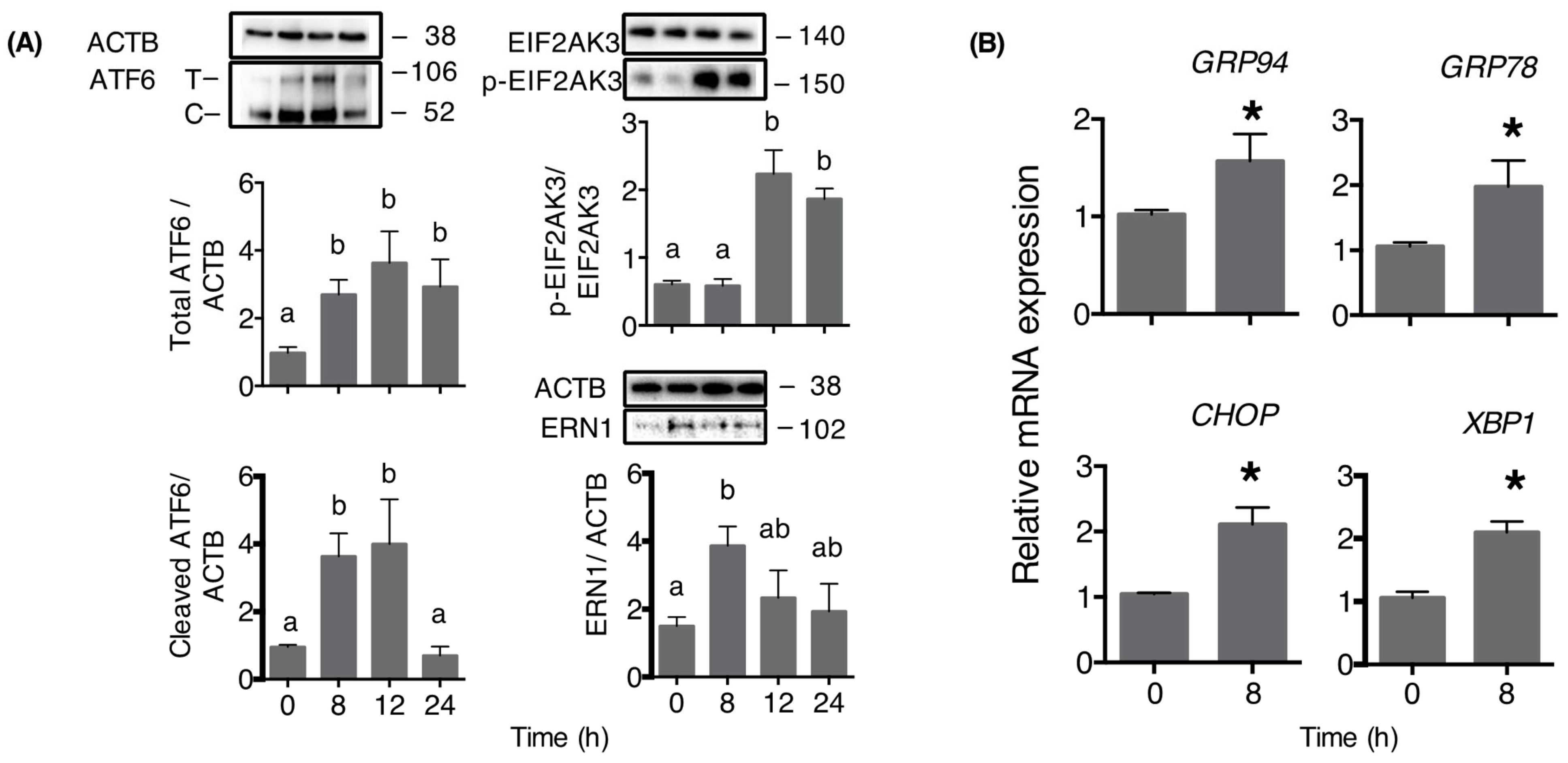
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Granulosa Cell Culture
5.2. Theca Cell Culture
5.3. Experimental Design
5.4. Total RNA Extraction and Real-Time PCR
5.5. Immunoblotting
5.6. Immunofluorescence
5.7. Flow Cytometry
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef] [PubMed]
- Twarużek, M.; Skrzydlewski, P.; Kosicki, R.; Grajewski, J. Mycotoxins survey in feed materials and feedingstuffs in years 2015–2020. Toxicon 2021, 202, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Pizzo, F.; Spicer, L.J.; Caloni, F. Fusarium mycotoxins: Effects on reproductive function in domestic animals—A review. Theriogenology 2013, 80, 557–564. [Google Scholar] [CrossRef]
- Rotter, B.A.; Thompson, B.K.; Lessard, M.; Trenholm, H.L.; Tryphonas, H. Influence of Low-Level Exposure to Fusarium Mycotoxins on Selected Immunological and Hematological Parameters in Young Swine1. Toxicol. Sci. 1994, 23, 117–124. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porc. Health Manag. 2016, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Alm, H.; Brüssow, K.P.; Torner, H.; Vanselow, J.; Tomek, W.; Dänicke, S.; Tiemann, U. Influence of Fusarium-toxin contaminated feed on initial quality and meiotic competence of gilt oocytes. Reprod. Toxicol. 2006, 22, 44–50. [Google Scholar] [CrossRef]
- Alm, H.; Greising, T.; Brussow, K.P.; Torner, H.; Tiemann, U. The influence of the mycotoxins deoxynivalenol and zearalenol on in vitro maturation of pig oocytes and in vitro culture of pig zygotes. Toxicol. In Vitro 2002, 16, 643–648. [Google Scholar] [CrossRef]
- Cortinovis, C.; Caloni, F.; Schreiber, N.B.; Spicer, L.J. Effects of fumonisin B1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology 2014, 81, 1042–1049. [Google Scholar] [CrossRef]
- Medvedova, M.; Kolesarova, A.; Capcarova, M.; Labuda, R.; Sirotkin, A.V.; Kovacik, J.; Bulla, J. The effect of deoxynivalenol on the secretion activity, proliferation and apoptosis of porcine ovarian granulosa cells in vitro. J. Environ. Sci. Health B 2011, 46, 213–219. [Google Scholar] [CrossRef]
- Guerrero-Netro, H.M.; Chorfi, Y.; Price, C.A. Effects of the mycotoxin deoxynivalenol on steroidogenesis and apoptosis in granulosa cells. Reproduction 2015, 149, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Pizzo, F.; Caloni, F.; Schreiber, N.B.; Cortinovis, C.; Spicer, L.J. In vitro effects of deoxynivalenol and zearalenone major metabolites alone and combined, on cell proliferation, steroid production and gene expression in bovine small-follicle granulosa cells. Toxicon 2016, 109, 70–83. [Google Scholar] [CrossRef]
- Zhou, H.-R.; Jia, Q.; Pestka, J.J. Ribotoxic stress response to the trichothecene deoxynivalenol in the macrophage involves the Src family kinase Hck. Toxicol. Sci. 2005, 85, 916–926. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.-R.; Lau, A.S.; Pestka, J.J. Role of double-stranded RNA-activated protein kinase R (PKR) in deoxynivalenol-induced ribotoxic stress response. Toxicol. Sci. 2003, 74, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam. A 2008, 25, 1128–1140. [Google Scholar] [CrossRef]
- Wu, Q.; Dohnal, V.; Huang, L.; Kuča, K.; Yuan, Z. Metabolic pathways of trichothecenes. Drug Metab. Rev. 2010, 42, 250–267. [Google Scholar] [CrossRef]
- Sundstøl Eriksen, G.; Pettersson, H.; Lundh, T. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.-P.F.L.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.-D.; et al. Microbial biotransformation of DON: Molecular basis for reduced toxicity. Sci. Rep. 2016, 6, 29105. [Google Scholar] [CrossRef] [Green Version]
- Dänicke, S.; Hegewald, A.-K.; Kahlert, S.; Kluess, J.; Rothkötter, H.J.; Breves, G.; Döll, S. Studies on the toxicity of deoxynivalenol (DON), sodium metabisulfite, DON-sulfonate (DONS) and de-epoxy-DON for porcine peripheral blood mononuclear cells and the Intestinal Porcine Epithelial Cell lines IPEC-1 and IPEC-J2, and on effects of DON and DONS on piglets. Food Chem. Toxicol. 2010, 48, 2154–2162. [Google Scholar] [CrossRef]
- Pierron, A.; Bracarense, A.P.F.L.; Cossalter, A.-M.; Laffitte, J.; Schwartz-Zimmermann, H.E.; Schatzmayr, G.; Pinton, P.; Moll, W.-D.; Oswald, I.P. Deepoxy-deoxynivalenol retains some immune-modulatory properties of the parent molecule deoxynivalenol in piglets. Arch. Toxicol. 2018, 92, 3381–3389. [Google Scholar] [CrossRef]
- Guerrero-Netro, H.M.; Barreta, M.H.; Costa, E.; Goetten, A.; Dupras, R.; Mills, L.; Koch, J.; Portela, V.M.; Price, C.A.; Chorfi, Y. Effects of the mycotoxin metabolite de-epoxy-deoxynivalenol (DOM-1) on embryo development and sperm motility in cattle. J. Appl. Toxicol. 2021, 41, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Netro, H.M.; Estienne, A.; Chorfi, Y.; Price, C.A. The mycotoxin metabolite deepoxy-deoxynivalenol increases apoptosis and decreases steroidogenesis in bovine ovarian theca cells. Biol. Reprod. 2017, 97, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, P.; Ron, D. The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humeau, J.; Leduc, M.; Cerrato, G.; Loos, F.; Kepp, O.; Kroemer, G. Phosphorylation of eukaryotic initiation factor-2α (eIF2α) in autophagy. Cell Death Dis. 2020, 11, 433. [Google Scholar] [CrossRef]
- B’chir, W.; Maurin, A.-C.; Carraro, V.; Averous, J.; Jousse, C.; Muranishi, Y.; Parry, L.; Stepien, G.; Fafournoux, P.; Bruhat, A. The eIF2α/ATF4 pathway is essential for stress-induced autophagy gene expression. Nucleic Acids Res. 2013, 41, 7683–7699. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Porter, K.; Parameswaran, N.; Bae, H.K.; Pestka, J.J. Role of GRP78/BiP Degradation and ER Stress in Deoxynivalenol-Induced Interleukin-6 Upregulation in the Macrophage. Toxicol. Sci. 2009, 109, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Katika, M.R.; Hendriksen, P.J.M.; Shao, J.; van Loveren, H.; Peijnenburg, A. Transcriptome analysis of the human T lymphocyte cell line Jurkat and human peripheral blood mononuclear cells exposed to deoxynivalenol (DON): New mechanistic insights. Toxicol. Appl. Pharmacol. 2012, 264, 51–64. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, K.; Jin, Y.; Fu, Y.; Wang, R.; Zhang, J. Protective Effects of Taraxasterol against Deoxynivalenol-Induced Damage to Bovine Mammary Epithelial Cells. Toxins 2022, 14, 211. [Google Scholar] [CrossRef]
- Islam, M.R.; Roh, Y.S.; Kim, J.; Lim, C.W.; Kim, B. Differential immune modulation by deoxynivalenol (vomitoxin) in mice. Toxicol. Lett. 2013, 221, 152–163. [Google Scholar] [CrossRef]
- Tang, Y.; Li, J.; Li, F.; Hu, C.-A.A.; Liao, P.; Tan, K.; Tan, B.; Xiong, X.; Liu, G.; Li, T.; et al. Autophagy protects intestinal epithelial Cells against Deoxynivalenol toxicity by alleviating oxidative stress via IKK signaling pathway. Free Radic. Biol. Med. 2015, 89, 944–951. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, Y.; Zhu, L.; Cao, L.; Xu, W.; Rahman, S.u.; Feng, S.; Li, Y.; Wu, J. Autophagy protects PC12 cells against deoxynivalenol toxicity via the Class III PI3K/beclin 1/Bcl-2 pathway. J. Cell. Physiol. 2020, 235, 7803–7815. [Google Scholar] [CrossRef]
- Yoshida, H.; Haze, K.; Yanagi, H.; Yura, T.; Mori, K. Identification of the cis-Acting Endoplasmic Reticulum Stress Response Element Responsible for Transcriptional Induction of Mammalian Glucose-regulated Proteins: Involvement of basic leucine zipper transcription factors. J. Biol. Chem. 1998, 273, 33741–33749. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.-H.; Iwakoshi Neal, N.; Glimcher Laurie, H. XBP-1 Regulates a Subset of Endoplasmic Reticulum Resident Chaperone Genes in the Unfolded Protein Response. Mol. Cell. Biol. 2003, 23, 7448–7459. [Google Scholar] [CrossRef] [Green Version]
- Rashid, H.-O.; Yadav, R.K.; Kim, H.-R.; Chae, H.-J. ER stress: Autophagy induction, inhibition and selection. Autophagy 2015, 11, 1956–1977. [Google Scholar] [CrossRef] [Green Version]
- Glister, C.; Richards, S.L.; Knight, P.G. Bone morphogenetic proteins (BMP) -4, -6, and -7 potently suppress basal and luteinizing hormone-induced androgen production by bovine theca interna cells in primary culture: Could ovarian hyperandrogenic dysfunction be caused by a defect in thecal BMP signaling? Endocrinology 2005, 146, 1883–1892. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5′→3′ |
|---|---|
| GRP78 | F: TGCGAAGCCCTATAGCTGAC R: AGTAGGTGGTACCCAGGTCG |
| GRP94 | F: TGCTGTGTGGAGAGGGAATG R: TCCTGTGACCACAATCCCAA |
| XBP1 | F: GCA GAG ACC AAG GGG AAT GG R: CTG CAG AGG TGC ACG TAG TC |
| MAP1LC3A | F: CCAGCAAAATCCCGGTGATAA R: TCATGTTGACATGGTCCGGG |
| GADD45 | F: TACGAGTCGGCCAAGCTGAT R: GTCCTCCTCTTCCTCGTCGAT |
| BAX | F: AACATGGAGCTGCAGAGGAT R: CAGTTGAAGTTGCCGTCAGA |
| BCL2 | F: ATGACTTCTCTCGGCGCTAC R: CTGAAGAGCTCCTCCACCAC |
| CHOP | F: GCACCAAGCATGAACAGTTG R: ATCGATGGTGGTTGGGTATG |
| BID | F: CTCCGTCCTGCTGCTCTTTC R: GTGGACGGCCTTCACCG |
| BECN1 | F: CCCAGCTGAAACCAGGAGAG R: GTGGACATCATCCTGGCTGG |
| ACTB | F: GGATGAGGCTCAGAGCAAGAGA R: TCGTCCCAGTTGGTGACGAT |
| H2AFZ | F: GCGGAATTCGAAATGGCTGG R: GGGAAACCGCCTTTGTCTTG |
| Name of Antibody | Manufacturer (Cat. No.) | Dilution | Blocking Solution | Incubation Time |
|---|---|---|---|---|
| β-actin (C4) | Santa Cruz (sc-47778 HRP, Dellas, TX, USA) | 1:5000 | Skim milk/TTBS | Overnight |
| LC3 | Novus Biologicals (NB100-2220, Englewood, CO, USA) | 1:1000 | TTBS | Overnight |
| EIF2AK3 | Cell Signaling (C33E10, Danvers, MA, USA) | 1:1000 | Skim milk | Overnight |
| phospho-EIF2AK3 | Cell signaling (T980 16F8) | 1:1000 | Skim milk | Overnight |
| ATF6α | Santa Cruz (SC-166659) | 2:1000 | Skim milk | 24 h |
| ERN1 | Bioss (SER726, Woburn, MA, USA) | 1:1000 | Skim milk | Overnight |
| Anti-mouse IgG, HRP conjugated | Calbiochem (402334, San Diego, CA, USA) | 1:10,000 | Skim milk/TTBS | 1 h |
| Anti-Rabbit IgG, HRP conjugate | Promega (W401B, Madison, WI, USA) | 1:10,000 | Skim milk/TTBS | 1 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Perea, A.D.; Guerrero-Netro, H.M.; Meza-Serrano, E.; Estienne, A.; Price, C.A. The Mycotoxin De-Epoxy-Deoxynivalenol (DOM-1) Increases Endoplasmic Reticulum Stress in Ovarian Theca Cells. Toxins 2023, 15, 228. https://doi.org/10.3390/toxins15030228
Reyes-Perea AD, Guerrero-Netro HM, Meza-Serrano E, Estienne A, Price CA. The Mycotoxin De-Epoxy-Deoxynivalenol (DOM-1) Increases Endoplasmic Reticulum Stress in Ovarian Theca Cells. Toxins. 2023; 15(3):228. https://doi.org/10.3390/toxins15030228
Chicago/Turabian StyleReyes-Perea, Angelica D., Hilda M. Guerrero-Netro, Europa Meza-Serrano, Anthony Estienne, and Christopher A. Price. 2023. "The Mycotoxin De-Epoxy-Deoxynivalenol (DOM-1) Increases Endoplasmic Reticulum Stress in Ovarian Theca Cells" Toxins 15, no. 3: 228. https://doi.org/10.3390/toxins15030228