The Effect of Environmental Factors on Mould Counts and AFB1 Toxin Production by Aspergillus flavus in Maize

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Kernel Infection
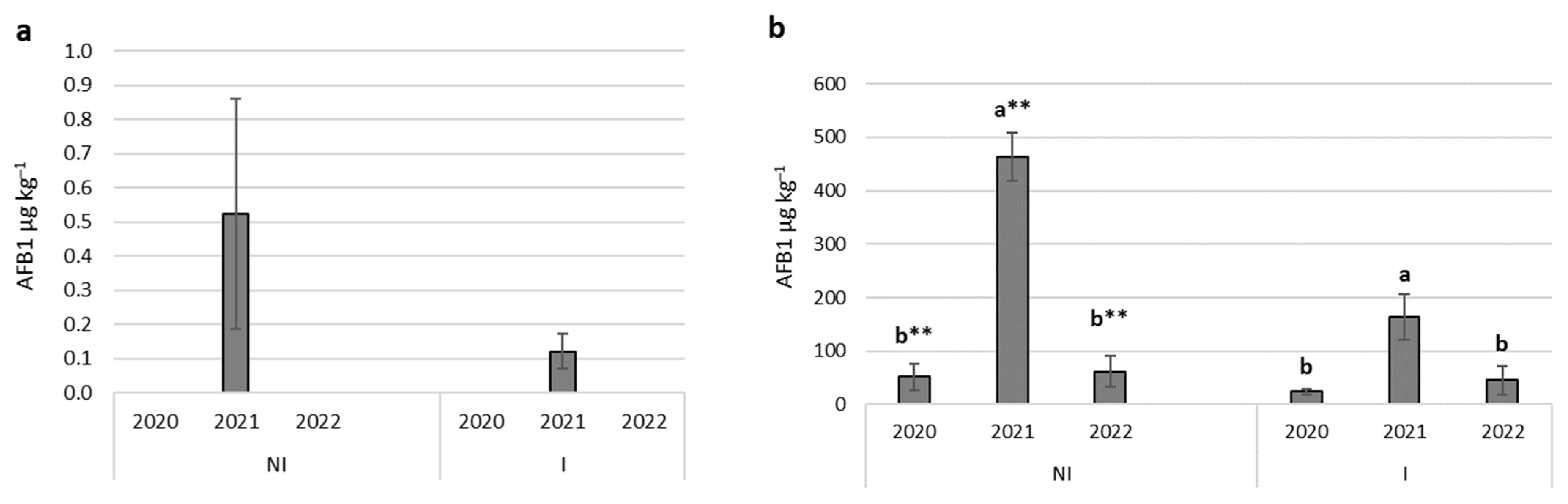
2.2. Aflatoxin Contamination
2.3. Seasonal Characteristics of Climatic Parameters
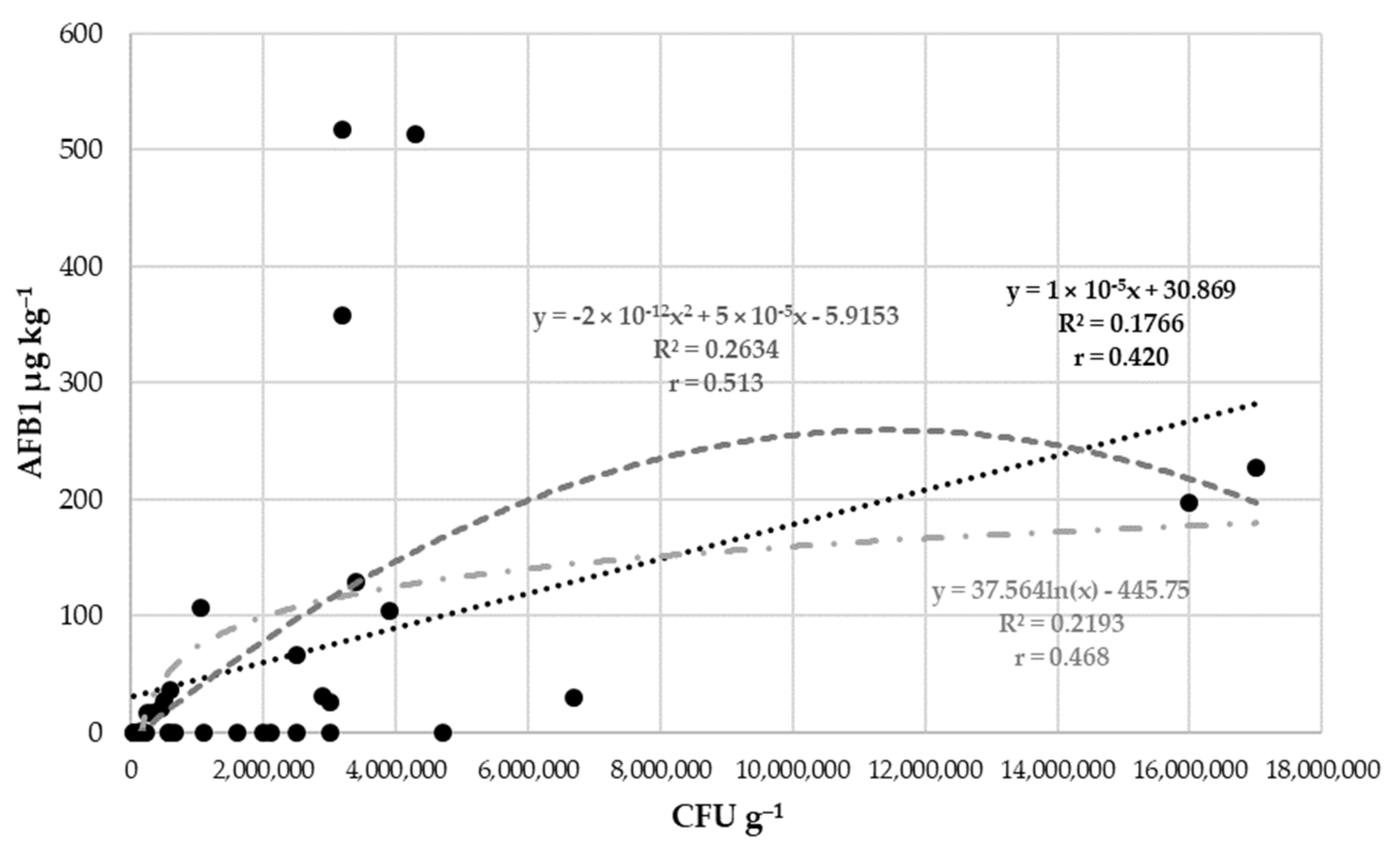
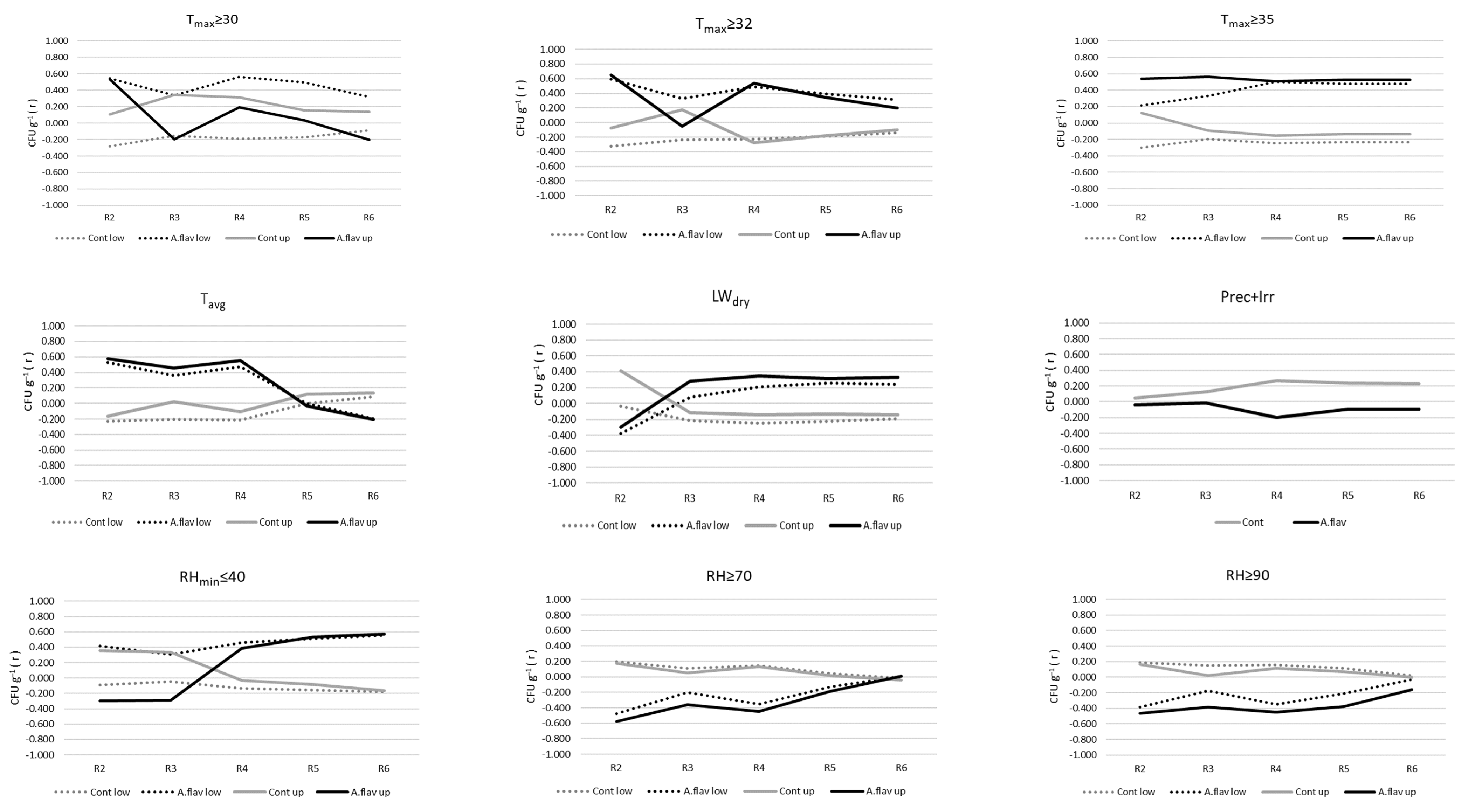
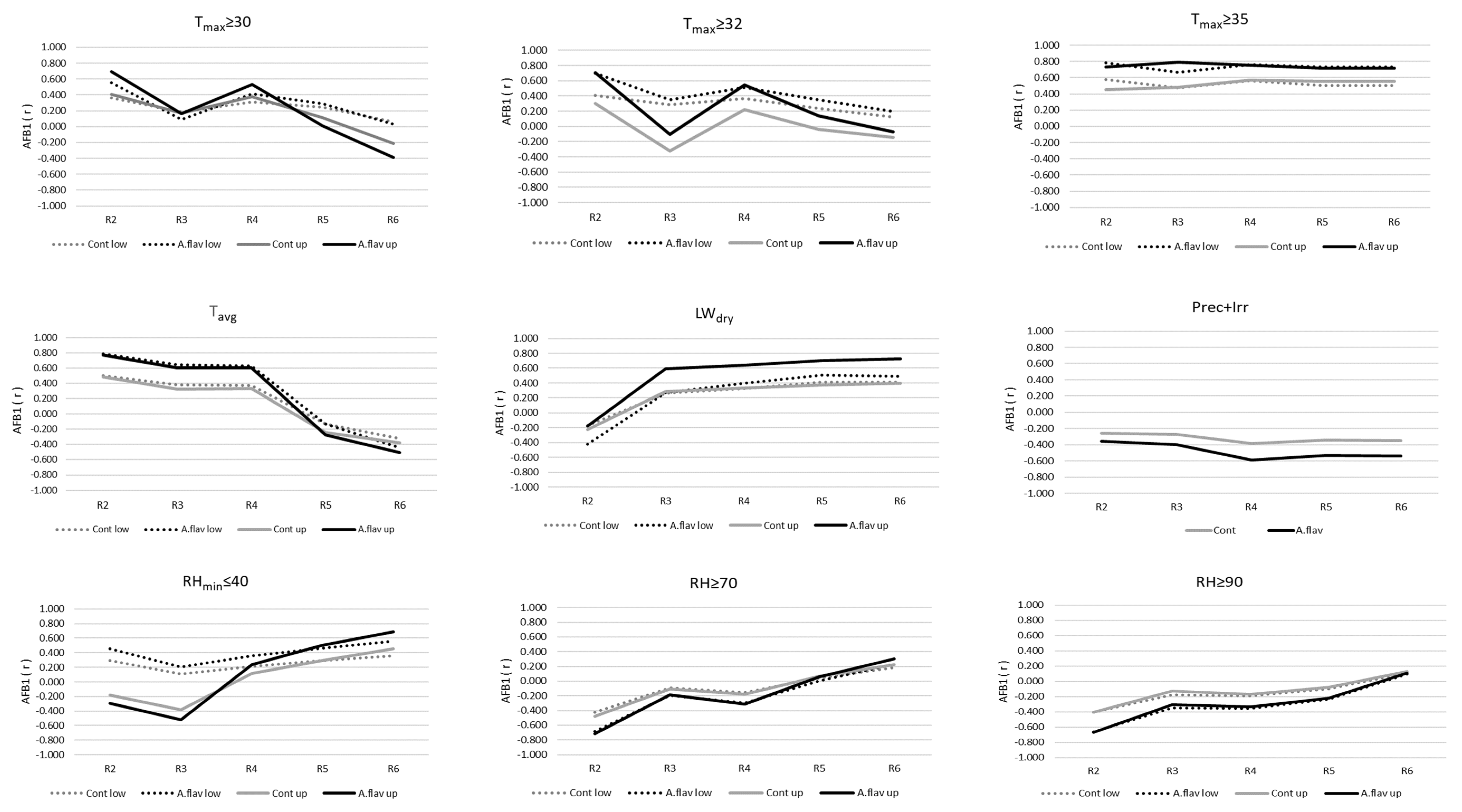
2.4. Relationships between Mould Colonisation, Toxin Production of A. flavus, and Environmental Parameters
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Field Experiment
5.2. Measurement of Environmental Parameters
5.2.1. Temperature and Humidity
5.2.2. Duration of Leaf Wetness
5.2.3. Precipitation
5.3. Post-Processing and Evaluation of the Data Series
5.3.1. Temperature and Relative Humidity
5.3.2. Leaf Wetness State
5.3.3. Precipitation
5.4. Aspergillus Flavus Artificial Contamination
5.5. Total Mould Count Determination
5.6. Detection of Aflatoxin B1 with HPLC-FD
5.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOStat. FAO Stat; FAO: Rome, Italy, 2021; Available online: http://www.fao.org/faostat (accessed on 2 December 2022).
- Hungarian Central Statistical Office, HCSO. Available online: https://www.ksh.hu/ (accessed on 11 November 2022).
- Pleadin, J.; Vulić, A.; Perši, N.; Škrivanko, M.; Capek, B.; Cvetnić, Ž. Aflatoxin B1 occurrence in maize sampled from Croatian farms and feed factories during 2013. Food Control 2014, 40, 286–291. [Google Scholar] [CrossRef]
- Orum, T.V.; Bigelow, D.M.; Cotty, P.J.; Nelson, M.R. Using predictions based on geostatistics to monitor trends in Aspergillus flavus strain composition. Phytopathology 1999, 89, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Pinke, Z.; Lövei, G.L. Increasing temperature cuts back crop yields in Hungary over the last 90 years. Glob. Chang. Biol. 2017, 23, 5426–5435. [Google Scholar] [CrossRef] [PubMed]
- Pinke, Z.; Decsi, B.; Kozma, Z.; Vári, Á.; Lövei, G.L. A spatially explicit analysis of wheat and maize yield sensitivity to changing groundwater levels in Hungary, 1961–2010. Sci. Total Environ. 2020, 715, 136555. [Google Scholar] [CrossRef] [PubMed]
- WHO. Evaluation of Certain Food Additives and Contaminants. In Forty-Ninth Report of the Joint FAO/WHO Expert Committee on Food Additives; WHO Technical Report Series 884; WHO: Geneva, Switzerland, 1999; ISBN 92-4-120884-8. [Google Scholar]
- Payne, G.A.; Widstorm, N.W. Aflatoxin in maize. Crit. Rev. Plant. Sci. 1992, 10, 423–440. [Google Scholar] [CrossRef]
- Varga, J.; Tóth, B.; Mesterhazy, A.; Teren, J.; Fazekas, B. Mycotoxigenic fungi and mycotoxins in foods and feeds in Hungary. In An Overview on Toxigenic Fungi and Mycotoxins in Europe; Logrieco, A., Visconti, A., Eds.; Kluwer Acedemic Publishers: Dordecht, The Netherlands, 2004; pp. 123–139. 252p, ISBN 1-4020-2645-5. [Google Scholar]
- Isakeit, T.; Murray, S.; Pekar, J. Influence of corn GMO traits for insect control on aflatoxin and fumonisin contamination in Texas. Phytopathology 2016, 106, 46. [Google Scholar]
- Marsh, S.F.; Payne, G.A. Preharvest infection of corn silks and kernels by Aspergillus flavus. Phytopathology 1984, 74, 1284–1289. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Locke, M.A. Spatial variability of Aspergillus flavus soil populations under different crops and corn grain colonisation and aflatoxins. Can. J. Bot. 2004, 82, 1768–1775. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Barbano, C.; Piva, G. Aflatoxin B1 contamination in maize related to the aridity index in North Italy. World Mycotoxin J. 2008, 1, 449–456. [Google Scholar] [CrossRef]
- Lahourar, A.; Marin, S.; Crespo-Sempere, A.; Said, S.; Sanchis, V. Effects of temperature, water activity and incubation time of fungal growth and aflatoxin B1 production by toxinogenic Aspergillus flavus isolates on sorghum seeds. Rev. Argent. Microbiol. 2016, 48, 78–85. [Google Scholar]
- Giorni, P.; Magan, N.; Battilani, P. Environmental factors modify carbon nutritional patterns and niche overlap between Aspergillus flavus and Fusarium verticillioides strains from maize. Int. J. Food Microbiol. 2009, 130, 213–218. [Google Scholar] [CrossRef]
- Sanchis, V.; Magan, N. Environmental conditions affecting mycotoxins. In Mycotoxins in Food; Magan, N., Olsen, M., Eds.; Woodhead Publishing: Cambridge, UK, 2004; pp. 174–189. [Google Scholar]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Aflatoxin in maize, a multifaceted answer of Aspergillus flavus governed by weather, host-plant and competitor fungi. J. Cereal Sci. 2016, 70, 256–262. [Google Scholar] [CrossRef]
- Rustom, I.Y.S. Aflatoxin in food and feed: Occurrence, legislation and inactivation by physical methods. Food Chem. 1997, 59, 57–67. [Google Scholar] [CrossRef]
- Farkas, Z.; Országh, E.; Engelhardt, T.; Csorba, S.; Kerekes, K.; Zentai, A.; Süth, M.; Nagy, A.; Miklós, G.; Molnár, K.; et al. A systematic review of the efficacy of interventions to control aflatoxins in the dairy production chain—Feed production and animal feeding interventions. Toxins 2022, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Camardo Leggieri, M.; Bertuzzi, T.; Pietri, A.; Battilani, P. Mycotoxin occurrence in maize produced in Northern Italy over the years 2009–2011: Focus on the role of crop related factors. Phytopathol. Mediterr. 2015, 54, 212–221. [Google Scholar]
- Hajnal, E.J.; Kos, J.; Krulj, J.; Krstovic, S.; Jajic, I.; Pezo, L.; Saric, B.; Nedeljkovic, N. Aflatoxins contamination of maize in Serbia: The impact of weather conditions in 2015. Food Addit. Contam. Part A 2017, 34, 1999–2010. [Google Scholar] [CrossRef]
- Baranyi, N.; Despot, D.J.; Palágyi, A.; Kiss, N.; Kocsubé, S.; Szekeres, A.; Kecskeméti, A.; Bencsik, O.; Vágvölgyi, C.; Klaric, M.S.; et al. Identification of Aspergillus species in Central Europe able to produce G-type aflatoxins. Acta Biol. Hung. 2015, 66, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Man, S.; Tofana, M.; Muste, S.; Paucean, A.; Birou, A. Natural occurrence of aflatoxins in wheat from central Transylvania. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca 2011, 68, 311–316. [Google Scholar]
- Girolami, F.; Barbarossa, A.; Badino, P.; Ghadiri, S.; Cavallini, D.; Zaghini, A.; Nebbia, C. Effects of turmeric powder on aflatoxin M1 and aflatoxicol excretion in milk from dairy cows exposed to aflatoxin B1 at EU maximum tolerable levels. Toxins 2022, 14, 430. [Google Scholar] [CrossRef]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–48. [Google Scholar] [CrossRef]
- Loi, M.; Logrieco, A.F.; Pusztahelyi, T.; Leiter, É.; Hornok, L.; Pócsi, I. Advanced Mycotoxin Control and Decontamination Techniques in View of an Increased Aflatoxin Risk in Europe Due to Climate Change. Front. Microbiol. 2023, 13, 1085891. [Google Scholar] [CrossRef] [PubMed]
- Dövényi-Nagy, T.; Rácz, C.; Molnár, K.; Bakó, K.; Szláma, Z.; Jóźwiak, Á.; Farkas, Z.; Pócsi, I.; Dobos, A.C. Pre-harvest modelling and mitigation of aflatoxins in maize in a changing climatic environment—A review. Toxins 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Toscano, P.; van der Fels-Kllerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Leggieri, M.C.; Toscano, P.; Battilani, P. Predicted aflatoxin B1 increase in Europe due to climate change: Actions and reactions at global level. Toxins 2021, 13, 292. [Google Scholar] [CrossRef]
- Szabo, B.; Toth, B.; Toldine Toth, E.; Varga, M.; Kovacs, N.; Varga, J.; Kocsube, S.; Palagyi, A.; Bagi, F.; Budakov, D.; et al. A new concept to secure food safety standards against Fusarium species and Aspergillus flavus and their toxins in maize. Toxins 2018, 10, 372. [Google Scholar] [CrossRef] [Green Version]
- Kocsubé, S.; Varga, J.; Szigeti, G.; Baranyi, N.; Suri, K.; Tóth, B.; Toldi, É.; Bartók, T.; Mesterházy, Á. Aspergillus species as mycotoxin producers in agricultural products in central Europe. Zb. Matice Srp. Za Prir. Nauk. 2013, 124, 13–25. [Google Scholar] [CrossRef]
- Dobolyi, C.S.; Sebők, F.; Varga, J.; Kocsubé, S.; Szigeti, G.; Baranyi, H.; Szécsi, Á.; Tóth, B.; Varga, M.; Kriszt, B.; et al. Occurance of aflatoxin producing Aspergillus flavus isolates in maize kernel in Hungary. Acta Aliment. 2013, 42, 451–459. [Google Scholar] [CrossRef]
- Kos, J.; Janić Hajnal, E.; Šarić, B.; Jovanov, P.; Mandić, A.; Ðuragić, O.; Kokić, B. Aflatoxins in maize harvested in the Republic of Serbia over the period 2012–2016. Food Addit. Contam. Part B 2018, 11, 246–255. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Szieberth, D.; Szabó, B.; Berényi, A.; Tóth, B. Mycotoxin Contamination of Maize (Zea mays, L.) Samples in Hungary, 2012–2017; Cereal Research Communications; Akadémiai Kiadó: Budapest, Hungary, 2022; Volume 50. [Google Scholar] [CrossRef]
- Virág, C.I.; Kutasy, E.T.; Lelesz, É.J.; Biró, G.; Tarcali, G.; Csüllög, K. Appearance of Aspergillus flavus and Aspergillus niger toxin-producing fungi on maize around Debrecen in 2019 and 2020. In Proceedings of the 44th Conference for Students of Agriculture and Veterinary Medicine with International Participation; Novi-Sad, Serbia, 15 December 2020, Proceedings Book; University of Novi Sad, Faculty of Agriculture: Novi Sad, Serbia, 2020; pp. 95–101. [Google Scholar]
- Magan, N.; Medina, A.; Aldred, D. Possible climate change effects on mycotoxin contamination of food crops pre- and post-harvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Paterson, R.; Lima, N. Further mycotoxin effects from climate change. Food Res. Int. 2011, 44, 2555–2566. [Google Scholar] [CrossRef] [Green Version]
- Kebede, H.; Abbas, H.K.; Fisher, K.D.; Bellaloui, N. Relationship between aflatoxin contamination and physiological responses of corn plants under drought and heat stress. Toxins 2012, 4, 1358–1403. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Chen, Z.-Y.; Lee, R.D.; Scully, B.T. Drought stress and preharvest aflatoxin contamination in agricultural commodity: Genetics, genomics and proteomics. J. Integr. Plant Biol. 2008, 50, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.K.; Duncan, H.E.; Payne, G.A.; Hamilton, P.B. Planting date, harvest date, and irrigation effects on infection and aflatoxin production by Aspergillus flavus in field corn. Phytopathology 1981, 66, 675–677. [Google Scholar]
- Abbas, H.K.; Wilkinson, J.R.; Zablotowicz, R.M.; Accinelli, R.M.; Abel, C.A.; Burns, H.A.; Weaver, M.A. Ecology of Aspergillus flavus, regulation of aflatoxin production, and management strategies to reduce aflatoxin contamination of corn. Toxin Rev. 2009, 28, 142–153. [Google Scholar] [CrossRef]
- Barontini, J.M.; Zanon, M.S.A.; Duretta, M.A.; Posse, A.M.R.; Torrico, A.K.; Monge, M.P.; Candela, R.E.; Chulze, S.N.; Pecci, M.P.G. Differential response of maize hybrids to field infection with Aspergillus flavus and aflatoxin accumulation in the Chaco Semi-arid region of Argentina. Crop Prot. 2022, 156, 105960. [Google Scholar] [CrossRef]
- Massomo, S.M.S. Aspergillus flavus and aflatoxin contamination in the maize value chain and what needs to be done in Tanzania. Sci. Afr. 2020, 10, e00606. [Google Scholar] [CrossRef]
- Camiletti, B.X.; Torrico, A.K.; Maurino, M.F.; Cristos, D.; Magnoli, C.; Lucini, E.I.; Pecci, M.P.G. Fungal screening and aflatoxin production by Aspergillus section Flavi isolated from pre-harvest maize ears grown in two Argentine regions. Crop Prot. 2017, 92, 41–48. [Google Scholar] [CrossRef]
- Burns, H.A.; Abbas, H.K. Effects of glufosinate–Ammonium and urea on aflatoxin and fumonisin levels in corn. Plant Health Prog. 2006, 7. [Google Scholar] [CrossRef] [Green Version]
- Diener, U.L.; Cole, R.J.; Sanders, T.H.; Payne, G.A.; Lee, L.S.; Klich, M.A. Epidemiology of aflatoxin, in formation by Aspergillus flavus. Annu. Rev. Phytopathol. 1987, 25, 240–270. [Google Scholar] [CrossRef]
- Windham, G.L.; Williams, W.P.; Hawkins, L.K.; Brooks, T.D. Effect of Aspergillus flavus inoculation methods and environmental conditions on aflatoxin accumulation in corn hybrids. Toxin Rev. 2009, 28, 70–78. [Google Scholar] [CrossRef]
- Marasas, W.F.O.; Gelderblom, W.C.A.; Shephard, G.S.; Vismer, H.F. Mycotoxins: A global problem. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J.F., Bandyopadhyay, R., Visconti, A., Eds.; Wallingford: Oxfordshire, UK, 2008; pp. 29–40. [Google Scholar]
- Shuttleworth, J.W.; Wallace, J.S. Evaporation from sparse crops—An energy combination theory. Q. J. R. Meteorol. Soc. 1985, 3, 839–855. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration. In Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage; FAO: Rome, Italy, 1998; Paper No. 56. [Google Scholar]
- Ritchie, S.W.; Hanway, J.J.; Benson, G.O.; Herman, J.C. How a corn plant develops. In Special Report No. 48; Iowa State University: Ames, IA, USA, 1993. [Google Scholar]
- Neild, R.E.; Newman, J.E. NCH-40 Growing Season Characteristics and Requirements in the Corn Belt. In National Corn Handbook; Cooperative Extension Service, Iowa State University: Ames, IA, USA, 1987. [Google Scholar]
- Abendroth, L.J.; Elmore, R.W.; Boyer, M.J.; Marlay, S.K. Corn Growth and Development. PMR 1009; Iowa State University Extension: Ames, IA, USA, 2011. [Google Scholar]
- Miedema, P. The effects of low temperature on Zea mays. Adv. Agron. 1982, 35, 93–128. [Google Scholar]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Global Change Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef] [PubMed]
- AG Clewer, A.G.; Scarisbrick, D.H. Practical Statistics and Experimental Design for Plant and Crop Science; John Wiley & Sons Ltd.: Chichester, UK, 2001. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CFU g−1 | |||||
|---|---|---|---|---|---|
| Treatment | Irrigation | Mean | SD | Min | Max |
| Control (non-inoculated) | NI | 692,555 b | 1,019,322 | 33,000 | 3,000,000 |
| I | 1,270,000 a | 1,439,237 | 30,000 | 4,700,000 | |
| A. flavus (inoculated) | NI | 2,865,555 b | 1,904,852 | 450,000 | 6,700,000 |
| I | 5,060,000 a | 6,288,750 | 240,000 | 17,000,000 | |
| AFB1 µg kg−1 | |||||
|---|---|---|---|---|---|
| Treatment | Irrigation | Mean | SD | Min | Max |
| Control (non-inoculated) | NI | 0.18 a | 0.41 | 0.00 | 1.30 |
| I | 0.04 b | 0.07 | 0.00 | 0.24 | |
| A.flavus (inoculated) | NI | 192.31 a ** | 201.38 | 20.28 | 517.49 |
| I | 77.42 b ** | 78.91 | 0.00 | 227.83 | |
| Start Date | End Date | Tavg | Tmax ≥ 30 | Tmax ≥ 32 | Tmax ≥ 35 | RHmin ≤ 40 | RH ≥ 70 dur. | RH ≥ 90 dur. | Prec. Sum | LW Dry | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | °C | number of days | day | day | mm | day | |||||
| R2 | 01. 08. | 07. 08. | 21.9 | 3 | 1 | 0 | 2 | 4.5 | 2.1 | 2.2 | 4.8 |
| R2–R3 | 01. 08. | 14. 08. | 22.5 | 10 | 5 | 0 | 4 | 8.6 | 4.5 | 2.2 | 9.5 |
| R2–R4 | 01. 08. | 23. 08. | 22.1 | 13 | 5 | 0 | 4 | 15.5 | 9.3 | 28.4 | 13.6 |
| R2–R5 | 01. 08. | 31. 08. | 22.0 | 16 | 7 | 0 | 8 | 19.8 | 11.3 | 28.6 | 19.3 |
| R2–R6 | 01. 08. | 12. 09. | 20.9 | 17 | 7 | 0 | 12 | 28.1 | 16.7 | 40.8 | 26.5 |
| 2021 | |||||||||||
| R2 | 27. 07 | 01. 08. | 25.7 | 6 | 5 | 1 | 3 | 2.6 | 1.1 | 2.2 | 4.6 |
| R2–R3 | 27. 07 | 09. 08. | 23.7 | 11 | 6 | 2 | 6 | 7.0 | 2.9 | 7.8 | 10.7 |
| R2–R4 | 27. 07 | 16. 08. | 24.0 | 17 | 11 | 5 | 13 | 10.3 | 4.1 | 8.8 | 16.4 |
| R2–R5 | 27. 07 | 27. 08. | 22.3 | 20 | 13 | 5 | 19 | 17.2 | 7.3 | 24.6 | 23.7 |
| R2–R6 | 27. 07 | 12. 09. | 20.2 | 20 | 13 | 5 | 28 | 26.8 | 13.6 | 35.8 | 32.8 |
| 2022 | |||||||||||
| R2 | 27. 07 | 02. 08. | 22.6 | 5 | 3 | 0 | 3 | 3.9 | 2.0 | 35.0 | 5.0 |
| R2–R3 | 27. 07 | 10. 08. | 23.0 | 10 | 7 | 1 | 8 | 6.2 | 2.7 | 35.0 | 9.3 |
| R2–R4 | 27. 07 | 17. 08. | 23.4 | 16 | 10 | 1 | 14 | 8.9 | 3.2 | 36.0 | 13.3 |
| R2–R5 | 27. 07 | 25. 08. | 23.5 | 21 | 14 | 2 | 17 | 13.5 | 5.5 | 40.2 | 16.7 |
| R2–R6 | 27. 07 | 02. 09. | 23.2 | 25 | 17 | 2 | 22 | 16.8 | 6.6 | 60.4 | 20.9 |
| Treatment | Canopy Level | Phenophase | Model | R | R2 | Predictors |
|---|---|---|---|---|---|---|
| A. flavus | lower | R2 | 1 | 0.594 | 0.353 | Tmax ≥ 32 |
| R4 | 1 | 0.559 | 0.313 | Tmax ≥ 30 | ||
| R5 | 1 | 0.511 | 0.261 | RHmin ≤ 40 | ||
| 2 | 0.677 | 0.459 | RHmin ≤ 40, RH ≥ 90 | |||
| R6 | 1 | 0.556 | 0.310 | RHmin ≤ 40 | ||
| upper | R2 | 1 | 0.647 | 0.418 | Tmax ≥ 32 | |
| R3 | 1 | 0.567 | 0.321 | Tmax ≥ 35 | ||
| R4 | 1 | 0.552 | 0.304 | Tavg | ||
| R5 | 1 | 0.538 | 0.289 | RHmin ≤ 40 | ||
| R6 | 1 | 0.574 | 0.33 | RHand ≤40 |
| Treatment | Canopy Level | Phenophase | Model | R | R2 | Predictors |
|---|---|---|---|---|---|---|
| Control | lower | R2 | 1 | 0.574 | 0.33 | Tmax ≥ 35 |
| R3 | 1 | 0.475 | 0.226 | Tmax ≥ 35 | ||
| R4 | 1 | 0.56 | 0.313 | Tmax ≥ 35 | ||
| R5 | 1 | 0.502 | 0.252 | Tmax ≥ 35 | ||
| R6 | 1 | 0.502 | 0.252 | Tmax ≥ 35 | ||
| upper | R2 | 1 | 0.488 | 0.238 | Tavg | |
| R3 | 1 | 0.483 | 0.233 | Tmax ≥ 35 | ||
| R4 | 1 | 0.569 | 0.324 | Tmax ≥ 35 | ||
| R5 | 1 | 0.553 | 0.306 | Tmax ≥ 35 | ||
| R6 | 1 | 0.553 | 0.306 | Tmax ≥ 35 | ||
| A. flavus | lower | R2 | 1 | 0.787 | 0.62 | Tavg |
| 2 | 0.844 | 0.712 | Tavg, Tmax ≥ 35 | |||
| R3 | 1 | 0.665 | 0.442 | Tmax ≥ 35 | ||
| 2 | 0.823 | 0.677 | Tmax ≥ 35, Prec+Irr | |||
| R4 | 1 | 0.763 | 0.582 | Tmax ≥ 35 | ||
| 2 | 0.834 | 0.695 | Tmax ≥ 35, Prec+Irr | |||
| R5 | 1 | 0.729 | 0.531 | Tmax ≥ 35 | ||
| 2 | 0.829 | 0.687 | Tmax ≥ 35, Prec+Irr | |||
| R6 | 1 | 0.729 | 0.531 | Tmax ≥ 35 | ||
| 2 | 0.833 | 0.693 | Tmax ≥ 35, Prec+Irr | |||
| upper | R2 | 1 | 0.774 | 0.6 | Tavg | |
| 2 | 0.834 | 0.695 | Tavg, RH ≥ 70 | |||
| R3 R4 | 1 | 0.788 | 0.622 | Tmax ≥ 35 | ||
| 1 | 0.753 | 0.567 | Tmax ≥ 35 | |||
| R5 | 1 | 0.72 | 0.519 | Tmax ≥ 35 | ||
| R6 | 1 | 0.723 | 0.522 | LW dry |
| Parameter | Temperature | Relative Humidity | Leaf Wetness | Precipitation |
|---|---|---|---|---|
| Sensor type | Onset HOBO RXW-THC-B-868 | Onset HOBO RXW-THC-B-868 | METER Env. PHYTOS-31 | Campbell Scientific ARG-100 |
| Number of sensors | 6 | 6 | 6 | 1 |
| Accuracy | ±0.2 °C | ±2.5% RH | - | ±4% |
| Resolution | 0.02 °C | 0.01% RH | 5 min | 0.2 mm/tip |
| Sampling frequency | 10 s | 10 s | 10 s | - |
| Logging interval | 5 min | 5 min | 5 min | 10 min |
| Averaging interval | 1 day | 1 day | 1 day (sum) | 1 day (sum) |
| Mounting height | 0.3 and 1.5 m | 0.3 and 1.5 m | 0.3 and 1.5 m | 1 m |
| Data logger applied | Onset Computer Corp. RX3000 | Onset Computer Corp. RX3000 | Campbell Scientific CR1000 | Campbell Scientific CR1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, K.; Rácz, C.; Dövényi-Nagy, T.; Bakó, K.; Pusztahelyi, T.; Kovács, S.; Adácsi, C.; Pócsi, I.; Dobos, A. The Effect of Environmental Factors on Mould Counts and AFB1 Toxin Production by Aspergillus flavus in Maize. Toxins 2023, 15, 227. https://doi.org/10.3390/toxins15030227
Molnár K, Rácz C, Dövényi-Nagy T, Bakó K, Pusztahelyi T, Kovács S, Adácsi C, Pócsi I, Dobos A. The Effect of Environmental Factors on Mould Counts and AFB1 Toxin Production by Aspergillus flavus in Maize. Toxins. 2023; 15(3):227. https://doi.org/10.3390/toxins15030227
Chicago/Turabian StyleMolnár, Krisztina, Csaba Rácz, Tamás Dövényi-Nagy, Károly Bakó, Tünde Pusztahelyi, Szilvia Kovács, Cintia Adácsi, István Pócsi, and Attila Dobos. 2023. "The Effect of Environmental Factors on Mould Counts and AFB1 Toxin Production by Aspergillus flavus in Maize" Toxins 15, no. 3: 227. https://doi.org/10.3390/toxins15030227