Seven Years of Monitoring Susceptibility to Cry1Ab and Cry1F in Asian Corn Borer

 ,
,  , and
, and
Abstract
:1. Introduction
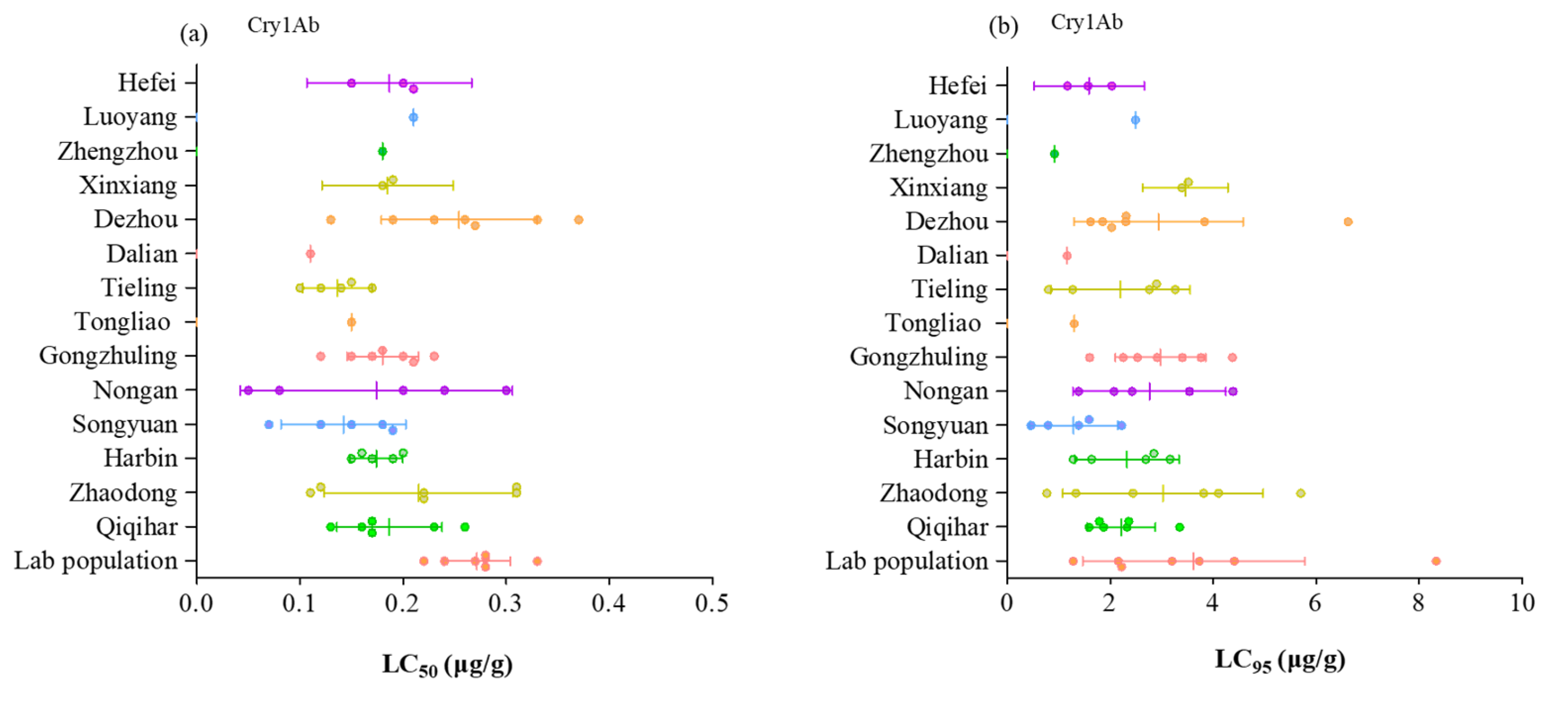
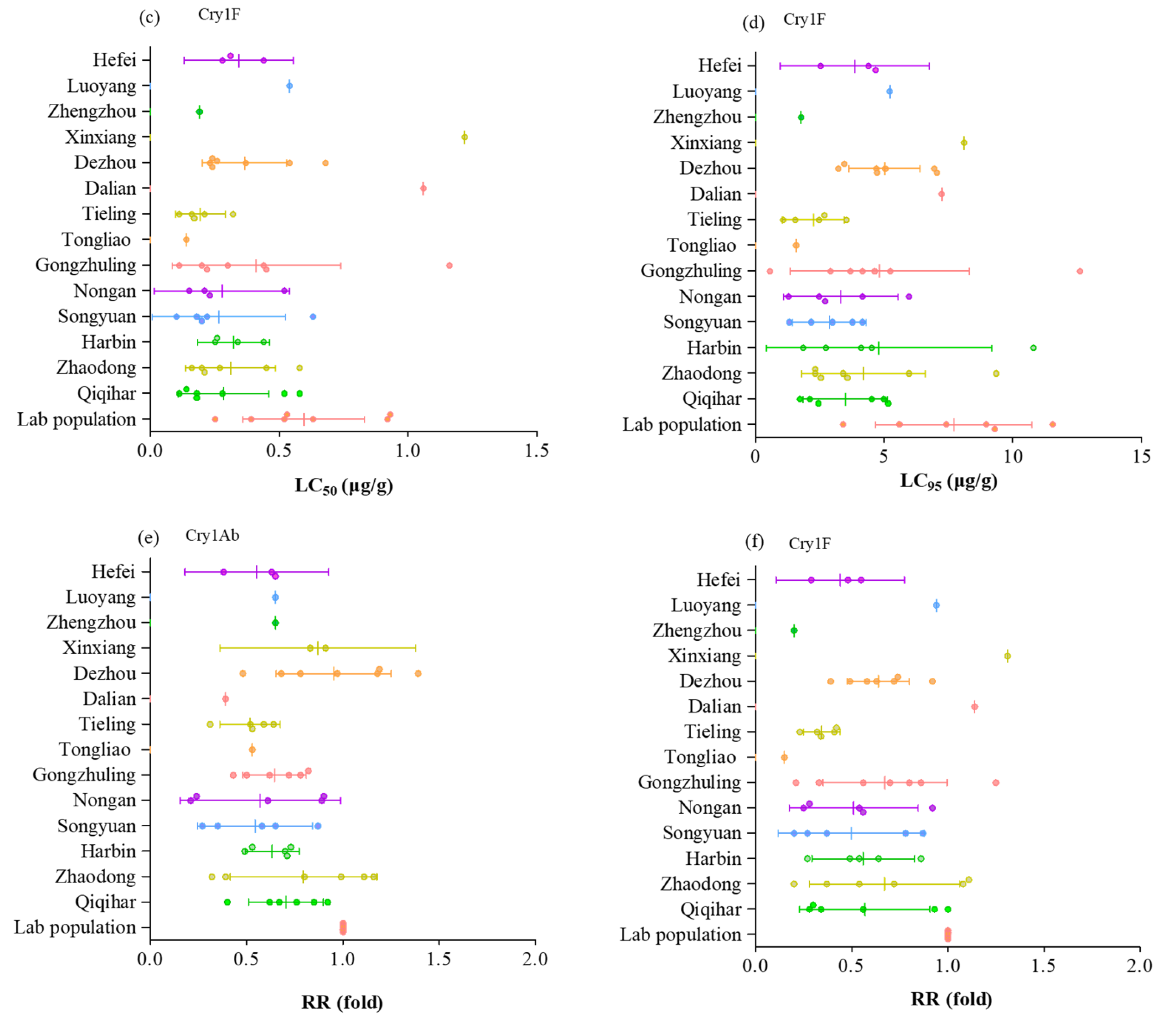
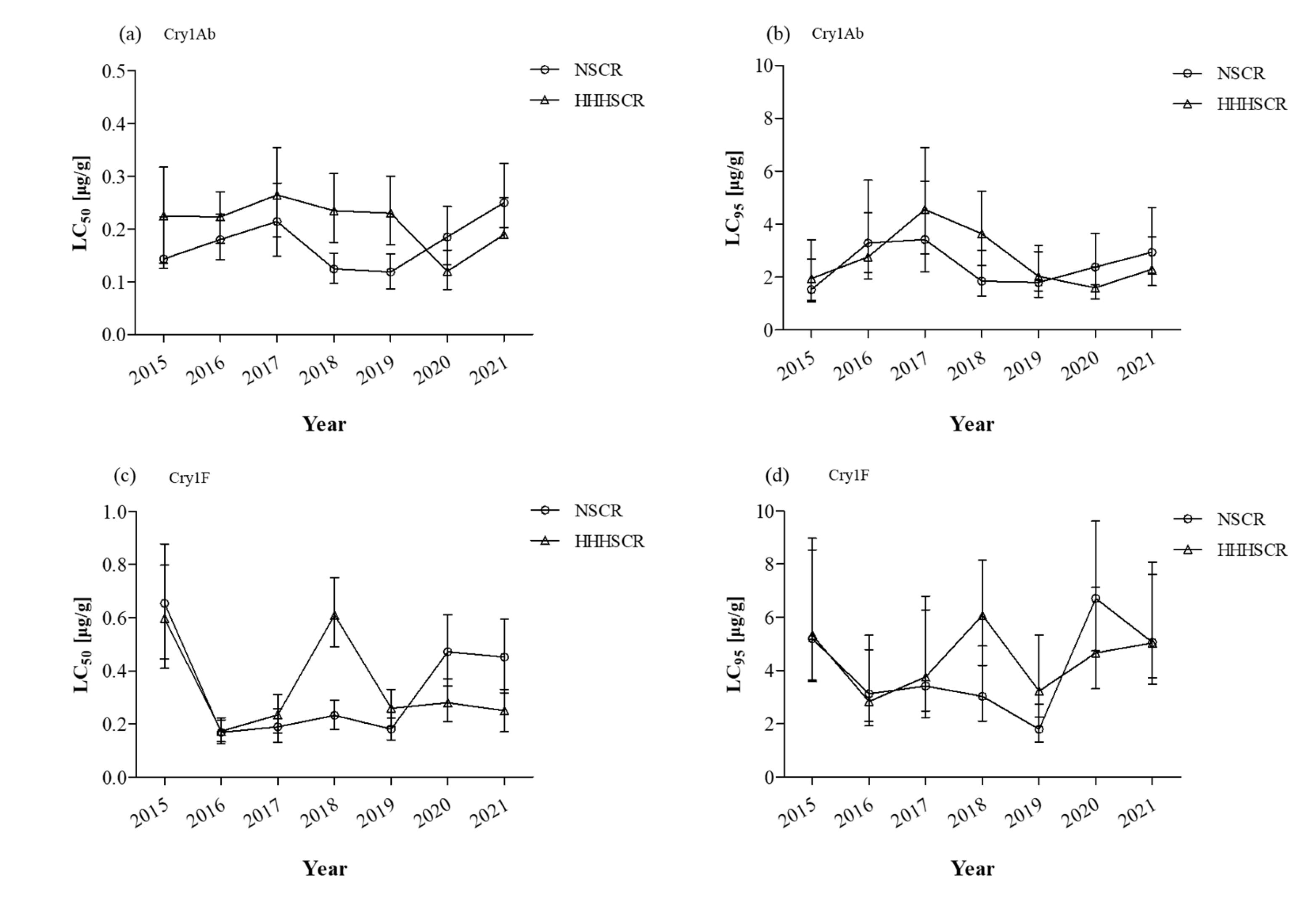
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Field Collections and Laboratory Rearing of O. furnacalis
5.2. Bt Toxins
5.3. Bioassays
5.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- ISAAA. Breaking Barriers with Breeding: A Primer on New Breeding Innovations for Food Security; ISAAA Brief No. 56; ISAAA: Ithaca, NY, USA, 2021. [Google Scholar]
- Yang, X.; Zhao, S.; Liu, B.; Gao, Y.; Hu, C.; Li, W.; Yang, Y.; Li, G.; Wang, L.; Yang, X.; et al. Bt maize can provide non-chemical pest control and enhance food safety in China. Plant Biotechnol. J. 2023, 21, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Shelton, A.M.; Zhao, J.Z.; Roush, R.T. Economic, Ecological, Food Safety, and Social Consequences of the Deployment of Bt Transgenic Plants. Annu. Rev. Èntomol. 2002, 47, 845–881. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wu, K.; Jiang, Y.; Guo, Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, M.G.; Yafuso, C.; Schmidt, C.; Huang, C.Y.; Rahman, M.; Olson, C.; Ellers-Kirk, C.; Orr, B.J.; Marsh, S.E.; Antilla, L.; et al. Farm-scale evaluation of transgenic cotton impacts on biodiversity, pesticide use, and yield. Proc. Natl. Acad. Sci. USA 2006, 16, 7571–7576. [Google Scholar] [CrossRef]
- Van, R.J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar]
- Ostrem, J.S.; Pan, Z.; Flexner, J.L.; Owens, E.; Binning, R.; Higgins, L.S. Monitoring Susceptibility of Western Bean Cutworm (Lepidoptera: Noctuidae) Field Populations to Bacillus thuringiensis Cry1F Protein. J. Econ. Èntomol. 2016, 109, 847–853. [Google Scholar] [CrossRef]
- Omoto, C.; Bernardi, O.; Salmeron, E.; Sorgatto, R.J.; Dourado, P.M.; Crivellari, A.; Carvalho, R.A.; Willse, A.; Martinelli, S.; Head, G.P. Field-evolved resistance to Cry1Ab maize by Spodoptera frugiperda in Brazil. Pest Manag. Sci. 2016, 72, 1727–1736. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop. Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Blanco, C.; Chiaravalle, W.; Dalla-Rizza, M.; Farias, J.; García-Degano, M.; Gastaminza, G.; Mota-Sánchez, D.; Murúa, M.; Omoto, C.; Pieralisi, B.; et al. Current situation of pests targeted by Bt crops in Latin America. Curr. Opin. Insect Sci. 2016, 15, 131–138. [Google Scholar] [CrossRef]
- Álvarez-Alfageme, F.; Devos, Y.; Camargo, A.M.; Arpaia, S.; Messéan, A. Managing resistance evolution to transgenic Bt maize in corn borers in Spain. Crit. Rev. Biotechnol. 2022, 42, 201–219. [Google Scholar] [CrossRef]
- ILSI [International Life Sciences Institute]. An Evaluation of Insect Resistance Management in Bt Field Corn: A Science-Based Framework for Risk Assessment and Risk Management; ILSI Press: Washington, DC, USA, 1998; p. 78. [Google Scholar]
- Siegfried, B.D.; Vaughn, T.T.; Spencer, T. Baseline Susceptibility of Western Corn Rootworm (Coleoptera: Crysomelidae) to Cry3Bb1 Bacillus thuringiensis Toxin. J. Econ. Èntomol. 2005, 98, 1320–1324. [Google Scholar] [CrossRef] [PubMed]
- Halliday, W.R.; Burham, K.P. Choosing the optimal diagnostic dose for monitoring insecticide resistance. J. Econ. Entomol. 1990, 83, 1151–1159. [Google Scholar] [CrossRef]
- Farias, J.R.; Horikoshi, R.J.; Santos, A.C.; Omoto, C. Geographical and Temporal Variability in Susceptibility to Cry1F Toxin from Bacillus thuringiensis in Spodoptera frugiperda (Lepidoptera: Noctuidae) Populations in Brazil. J. Econ. Èntomol. 2014, 107, 2182–2189. [Google Scholar] [CrossRef]
- Marçon, P.C.R.G.; Siegfried, B.D.; Spencer, T.; Hutchison, W.D. Development of Diagnostic Concentrations for Monitoring Bacillus thuringiensis Resistance in European Corn Borer (Lepidoptera: Crambidae). J. Econ. Èntomol. 2000, 93, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Nafus, D.M.; Schreiner, I.H. Review of the biology and control of the Asian corn borer, Ostrinia furnacalis (Lep: Pyralidae). Int. J. Pest Manag. 1991, 37, 41–56. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Lu, X.; He, K.L.; Zhou, D.R. Review of history, present situation and prospect of the Asian maize borer research in China. J. Shenyang Agric. Univ. 2000, 31, 402–412. [Google Scholar]
- Alcantara, E.; Atienza, M.M.; Camacho, L.; Parimi, S. Baseline susceptibility of Philippine Ostrinia furnacalis (Lepidoptera: Crambidae) populations to insecticidal Cry1A.105 and Cry2Ab2 proteins and validation of candidate diagnostic concentration for monitoring resistance. Biodiversitas 2021, 22, 956–960. [Google Scholar] [CrossRef]
- Le, D.K.; Le, Q.K.; Tran, T.T.H.; Nguyen, D.V.; Dao, T.H.; Nguyen, T.T.; Truong, X.L.; Nguyen, Q.C.; Pham, H.P.; Phan, T.T.T.; et al. Baseline susceptibility of Asian corn borer (Ostrinia furnacalis (Guenée)) populations in Vietnam to Cry1Ab insecticidal protein. J. Asia Pac. Èntomol. 2019, 22, 493–498. [Google Scholar] [CrossRef]
- Sun, D.D.; Quan, Y.D.; Wang, Y.Q.; Wang, Z.Y.; He, K.L. Resistance of transgenic Bt maize (Ruifeng125, DBN9936 &DBN9978) to Asian corn borer. Plant Protect. 2021, 47, 206–211. [Google Scholar]
- Chang, X.; Wang, W.; Shen, Z.C.; Ye, G.Y. Evaluation of transgenic cry1Ab/cry2Aj maize for its resistance to Ostrinia furnacalis. Acta Phytophy. Sin. 2013, 40, 339–344. [Google Scholar]
- Gujar, G.T.; Trisyono, Y.A.; Chen, M. Genetically Modified Crops in Asia Pacific; CSIRO Publishing: Melbourne, VIC, Australia, 2021; pp. 107–118. [Google Scholar]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defining Terms for Proactive Management of Resistance to Bt Crops and Pesticides. J. Econ. Èntomol. 2014, 107, 496–507. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Wang, Z.; Wen, L.; Bai, S.; Ma, X.; Yao, Z. Determination of baseline susceptibility to Cry1Ab protein for Asian corn borer (Lep., Crambidae). J. Appl. Èntomol. 2005, 129, 407–412. [Google Scholar] [CrossRef]
- Liu, X.; Liu, S.; Long, Y.; Wang, Y.; Zhao, W.; Shwe, S.M.; Wang, Z.; He, K.; Bai, S. Baseline Susceptibility and Resistance Allele Frequency in Ostrinia furnacalis in Relation to Cry1Ab Toxins in China. Toxins 2022, 14, 255. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Gore, J.; Leonard, B.R.; Jones, R.H. Influence of Agronomic Hosts on the Susceptibility of Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) to Genetically Engineered and Non-Engineered Cottons. Environ. Èntomol. 2003, 32, 103–110. [Google Scholar] [CrossRef]
- Zhang, H.; Tian, W.; Zhao, J.; Jin, L.; Yang, J.; Liu, C.; Yang, Y.; Wu, S.; Wu, K.; Cui, J.; et al. Diverse genetic basis of field-evolved resistance to Bt cotton in cotton bollworm from China. Proc. Natl. Acad. Sci. USA 2012, 109, 10275–10280. [Google Scholar] [CrossRef] [PubMed]
- Deans, C.; Behmer, S.T.; Sword, G. The impact of nutrition on susceptibility to Cry endotoxins in Helicoverpa zea (Lepidoptera: Noctuidae). In Proceedings of the Entomological Society of America Meeting 2014, Portland, OR, USA, 16–19 November 2014. [Google Scholar]
- Gujar, G.T.; Kalia, V.; Kumari, A.; Singh, B.P.; Mittal, A.; Nair, R.; Mohan, M. Helicoverpa armigera baseline susceptibility to Bacillus thuringiensis Cry toxins and resistance management for Bt cotton in India. J. Invertebr. Pathol. 2007, 95, 214–219. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Spencer, T.; Crespo, A.L.; Storer, N.P.; Head, G.P.; Owens, E.D.; Guyer, D. Ten Years of Bt Resistance Monitoring in the European Corn Borer: What We Know, What We Don’t Know, and What We Can Do Better. Am. Èntomol. 2007, 53, 208–214. [Google Scholar] [CrossRef]
- Chakroun, M.; Bel, Y.; Caccia, S.; Abdelkefi-Mesrati, L.; Escriche, B.; Ferré, J. Susceptibility of Spodoptera frugiperda and Sexigua to Bacillus thuringiensis Vip3Aa insecticidal protein. J. Invertebr. Pathol. 2012, 110, 334–339. [Google Scholar] [CrossRef]
- Lemes, A.R.N.; Davolos, C.C.; Legori, P.C.B.C.; Fernandes, O.; Ferré, J.; Lemos, M.V.F.; Desiderio, J.A. Synergism and Antagonism between Bacillus thuringiensis Vip3A and Cry1 Proteins in Heliothis virescens, Diatraea saccharalis and Spodoptera frugiperda. PLoS ONE 2014, 9, e107196. [Google Scholar] [CrossRef]
- Burd, A.D.; Gould, F.J.; Bradley, R.; Vanduyn, J.W.; Moar, W.J. Estimated frequency of non-recessive Bt resistance genes in bollworm, Helicoverpa zea (Bolddie) (Lepidoptera: Noctuidae) in eastern North Carolina. J. Econ. Entomol. 2003, 96, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.; Cayabyab, B.F.; Alcantara, E.P.; Huang, F.; He, K.; Nickerson, K.W.; Siegfried, B.D. Comparative binding of Cry1Ab and Cry1F Bacillus thuringiensis toxins to brush border membrane proteins from Ostrinia nubilalis, Ostrinia furnacalis and Diatraea saccharalis (Lepidoptera: Crambidae) midgut tissue. J. Invertebr. Pathol. 2013, 114, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Quan, Y.; Yang, J.; Shu, C.; Wang, Z.; Zhang, J.; Gatehouse, A.M.R.; Tabashnik, B.E.; He, K. Evolution of Asian Corn Borer Resistance to Bt Toxins Used Singly or in Pairs. Toxins 2019, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Zhou, D.R.; He, K.L. Studies on mass rearing of Asian corn borer: Development of a satisfactory non-agar semi-artificial diet and its use. Acta Phytophyl. Sin. 1999, 26, 324–328. [Google Scholar]
- Payton, M.E.; Greenstone, M.H.; Schenker, N. Overlapping confidence intervals or standard error intervals: What do they mean in terms of statistical significance? J. Insect Sci. 2003, 3, 34. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
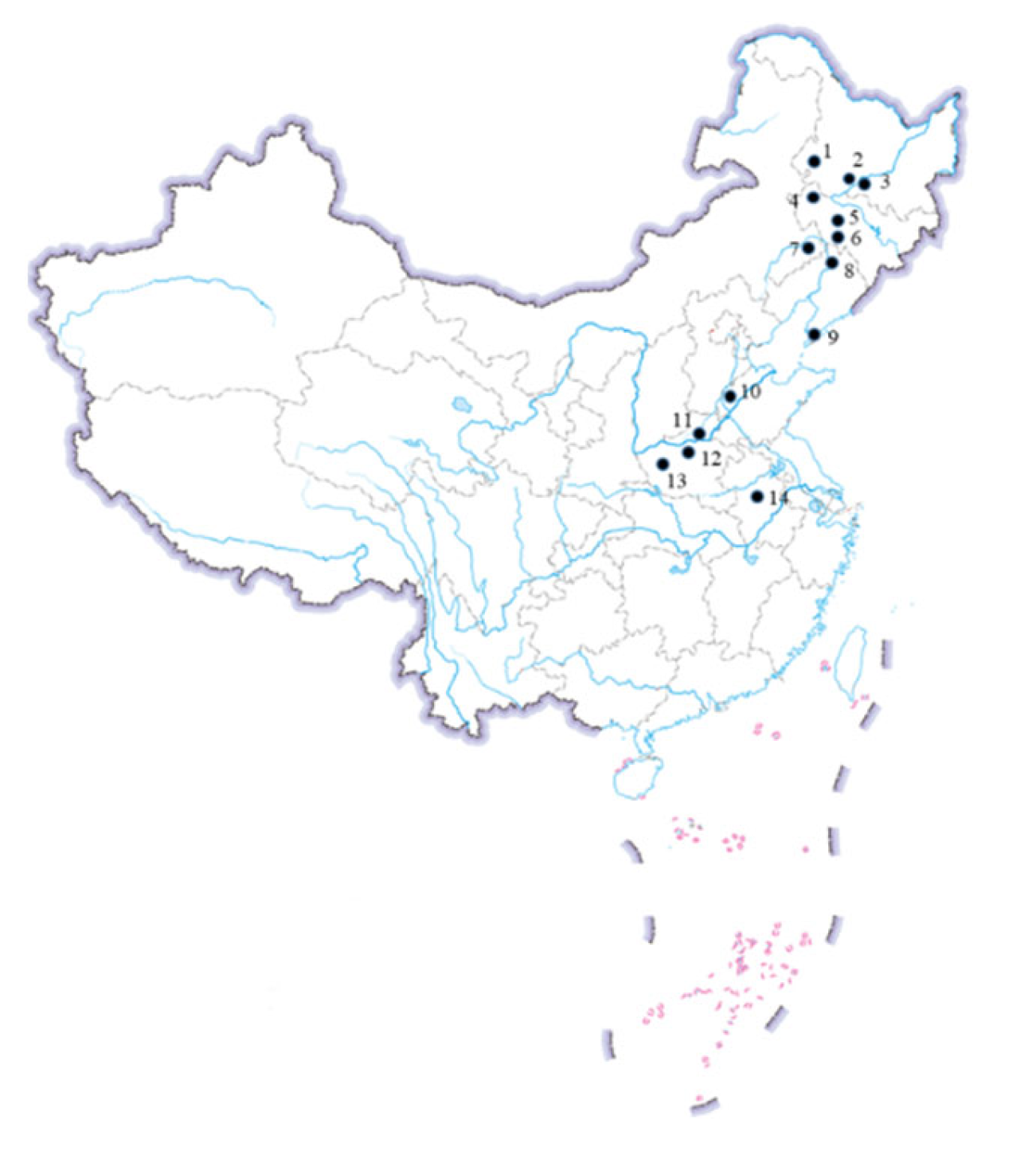
| Province | Field Population | Map No. | Region |
|---|---|---|---|
| Heilongjiang | Qiqihar | 1 | NSCR |
| Heilongjiang | Zhaodong | 2 | NSCR |
| Heilongjiang | Harbin | 3 | NSCR |
| Jilin | Songyuan | 4 | NSCR |
| Jilin | Nongan | 5 | NSCR |
| Jilin | Gongzhuling | 6 | NSCR |
| Inner Mongolia | Tongliao | 7 | NSCR |
| Liaoning | Tieling | 8 | NSCR |
| Liaoning | Dalian | 9 | NSCR |
| Shandong | Dezhou | 10 | HHHSCR |
| Henan | Xinxiang | 11 | HHHSCR |
| Henan | Zhengzhou | 12 | HHHSCR |
| Henan | Luoyang | 13 | HHHSCR |
| Anhui | Hefei | 14 | HHHSCR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhao, W.; Han, S.; Wang, L.; Chang, X.; Liu, K.; Quan, Y.; Wang, Z.; He, K. Seven Years of Monitoring Susceptibility to Cry1Ab and Cry1F in Asian Corn Borer. Toxins 2023, 15, 137. https://doi.org/10.3390/toxins15020137
Wang Y, Zhao W, Han S, Wang L, Chang X, Liu K, Quan Y, Wang Z, He K. Seven Years of Monitoring Susceptibility to Cry1Ab and Cry1F in Asian Corn Borer. Toxins. 2023; 15(2):137. https://doi.org/10.3390/toxins15020137
Chicago/Turabian StyleWang, Yueqin, Wenlu Zhao, Shuang Han, Lianxia Wang, Xue Chang, Kaiqiang Liu, Yudong Quan, Zhenying Wang, and Kanglai He. 2023. "Seven Years of Monitoring Susceptibility to Cry1Ab and Cry1F in Asian Corn Borer" Toxins 15, no. 2: 137. https://doi.org/10.3390/toxins15020137