Novel Endophytic Pseudescherichia sp. GSE25 Strain Significantly Controls Fusarium graminearum and Reduces Deoxynivalenol in Wheat

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of Endophytic Bacteria to Control F. graminearum Pathogenicity and DON Production
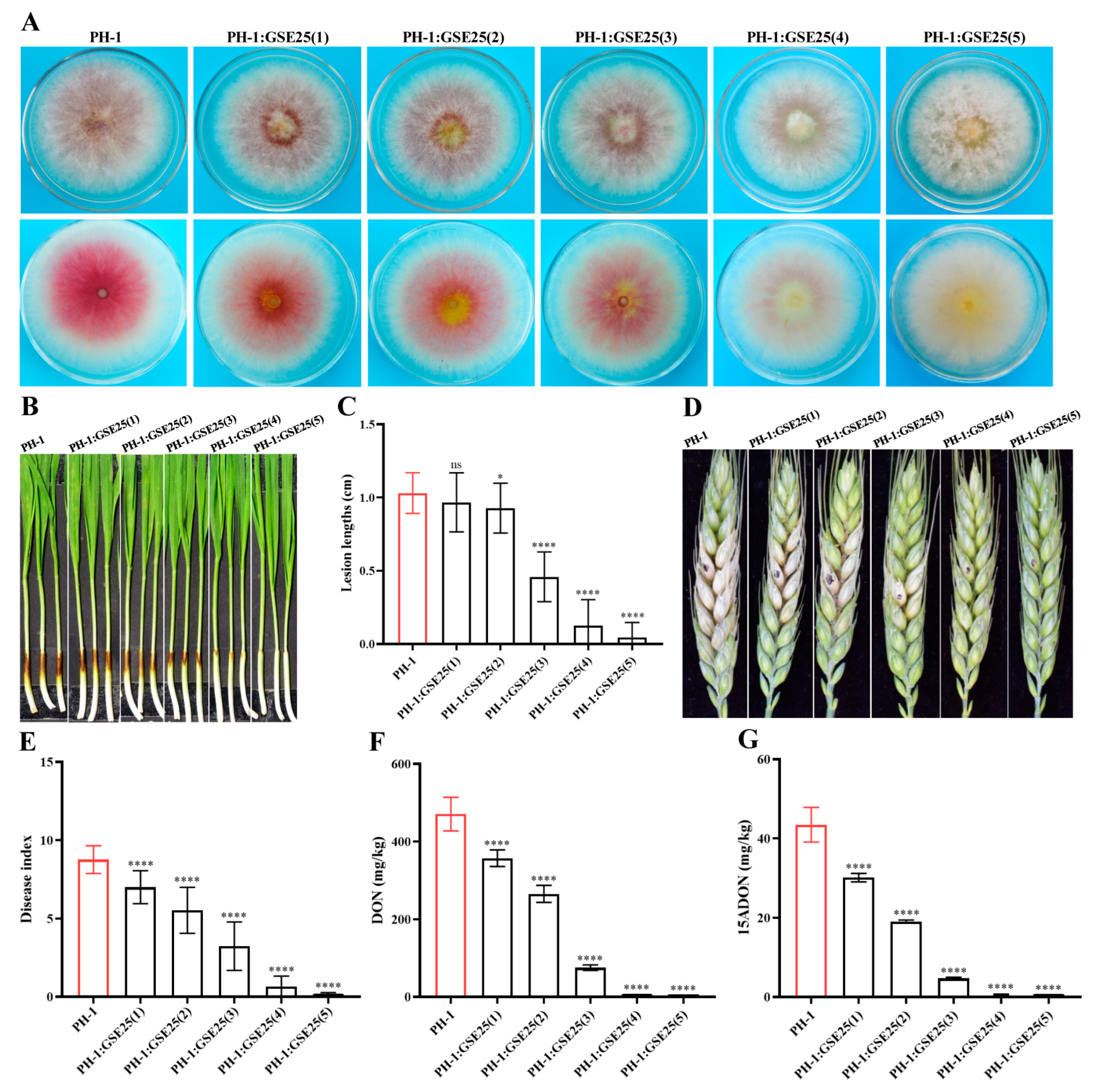
2.2. Biosafety Properties, Plant Pathogenicity Test, and Growth-Promoting Ability of Strain GSE25
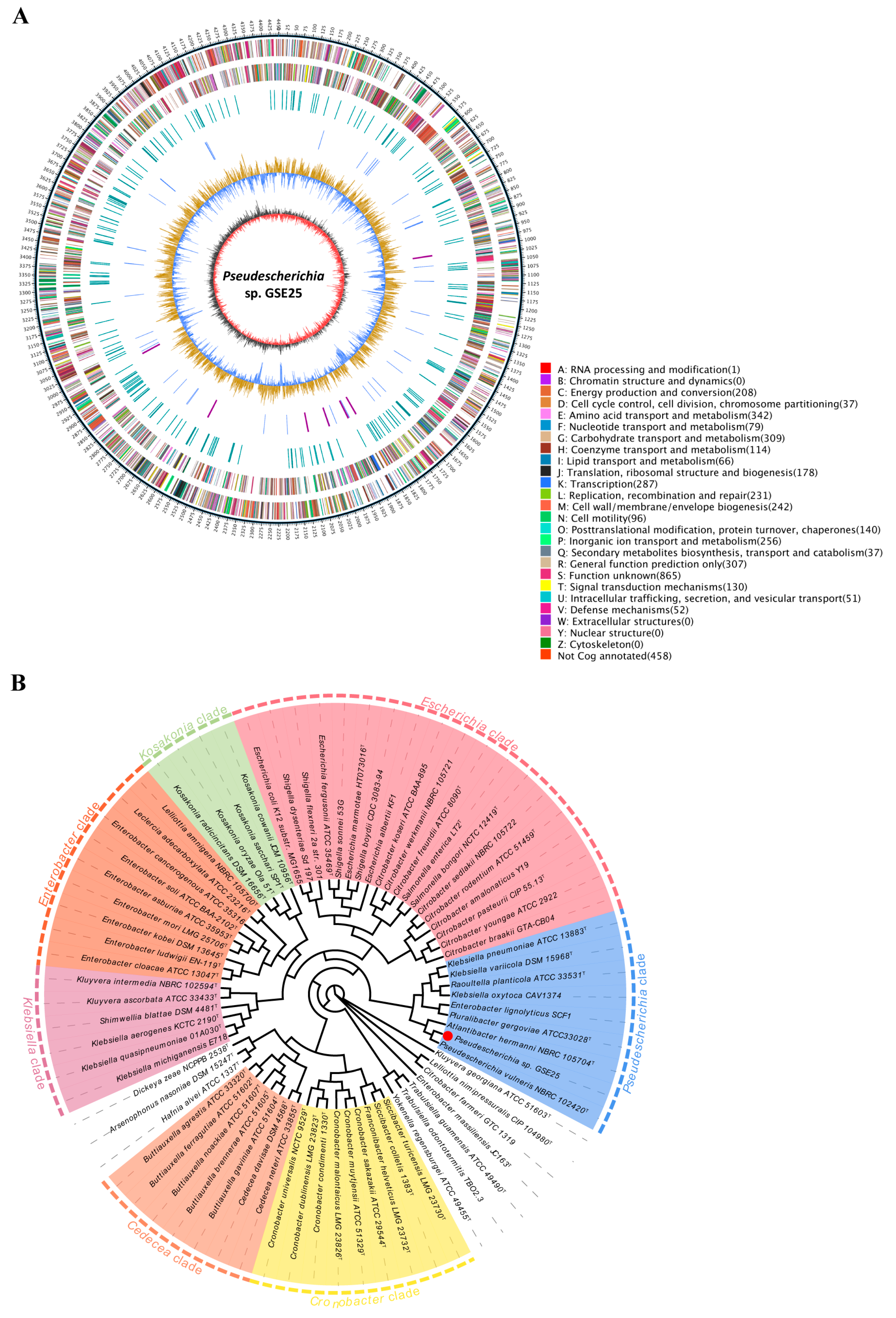
2.3. Whole Genome Sequence and Phylogenetic Relationship of Strain GSE25
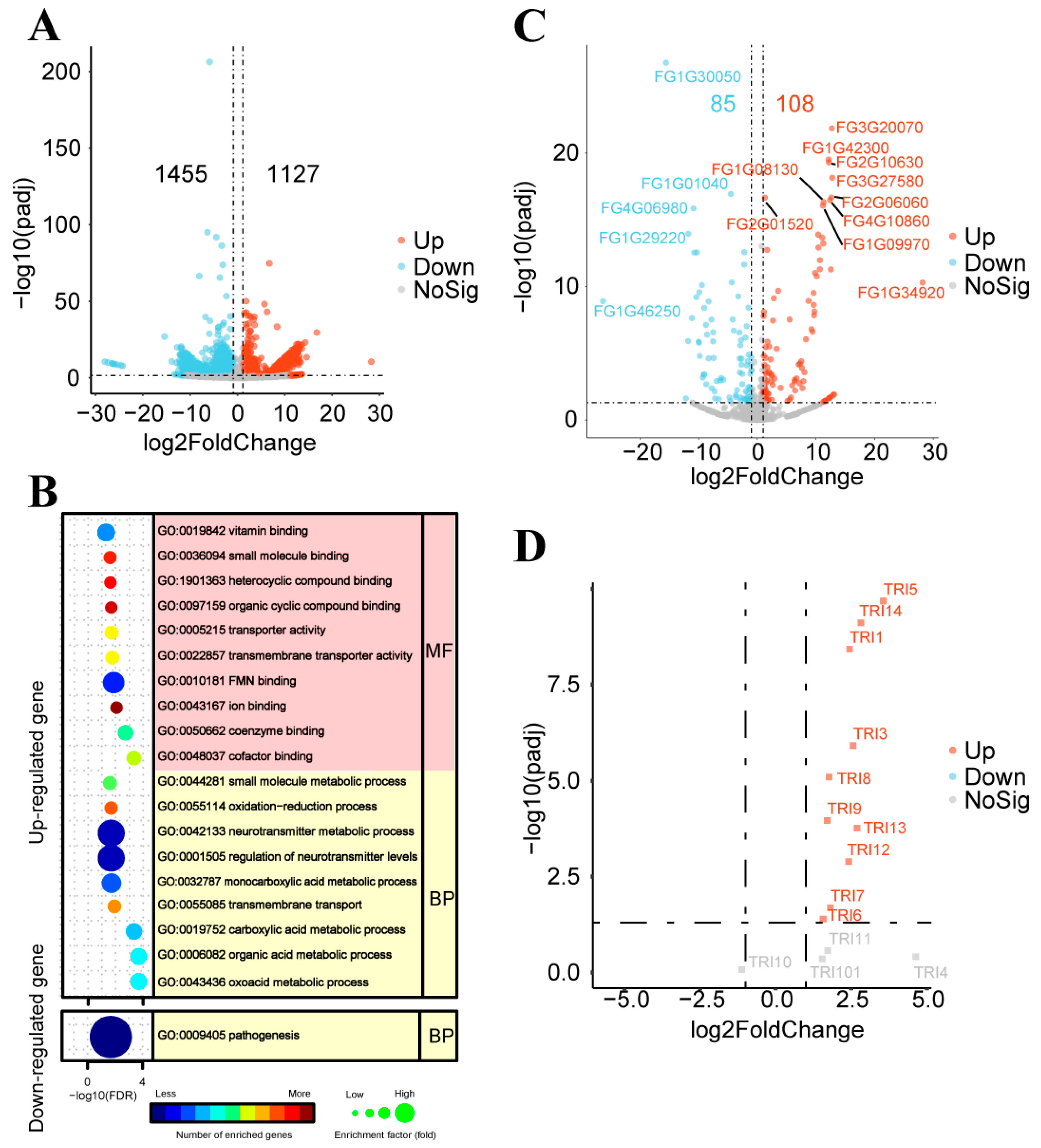
2.4. DNA-seq and RNA-seq Analyses of Inhibitory Effects of Strain GSE25 on F. graminearum PH-1
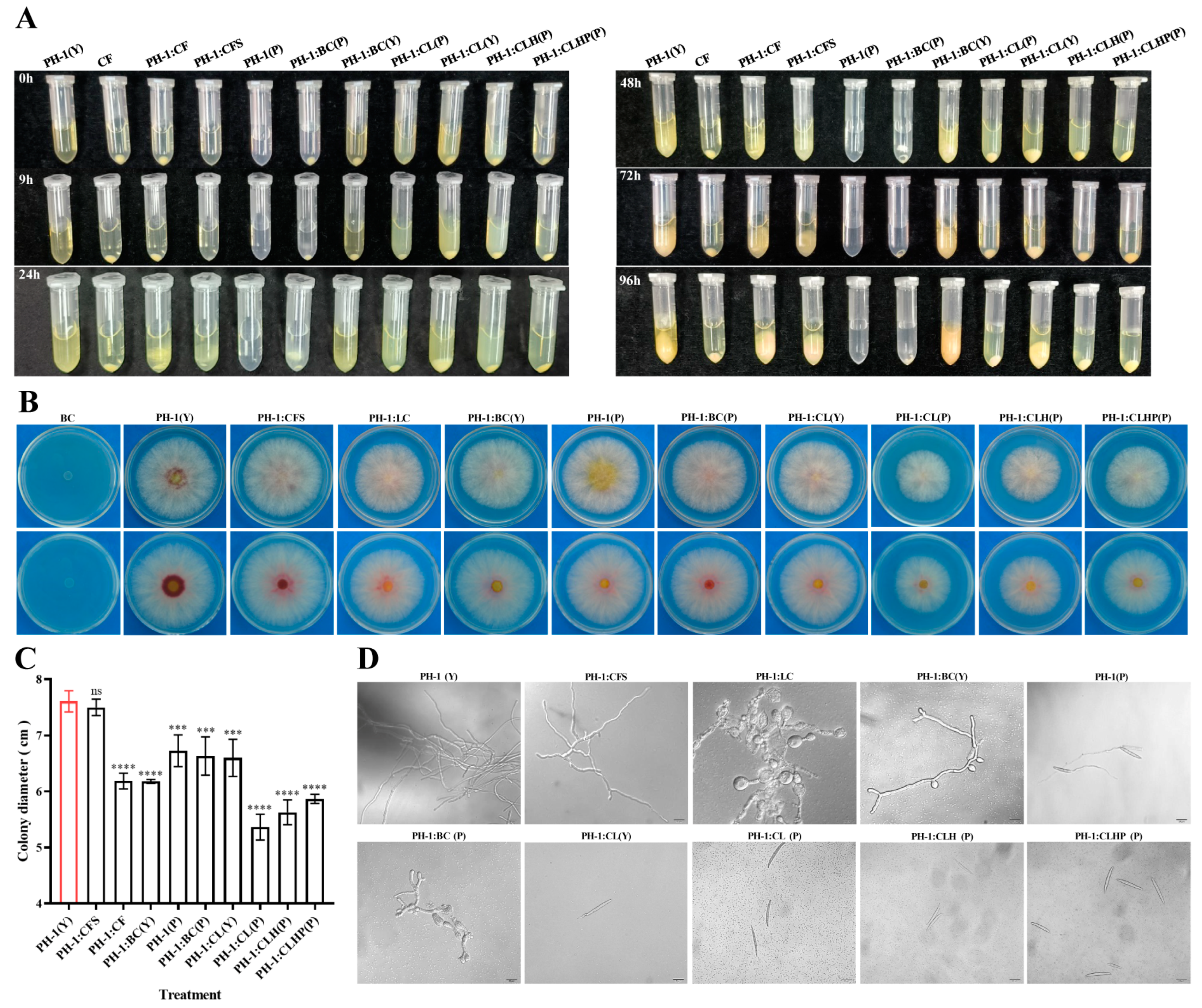
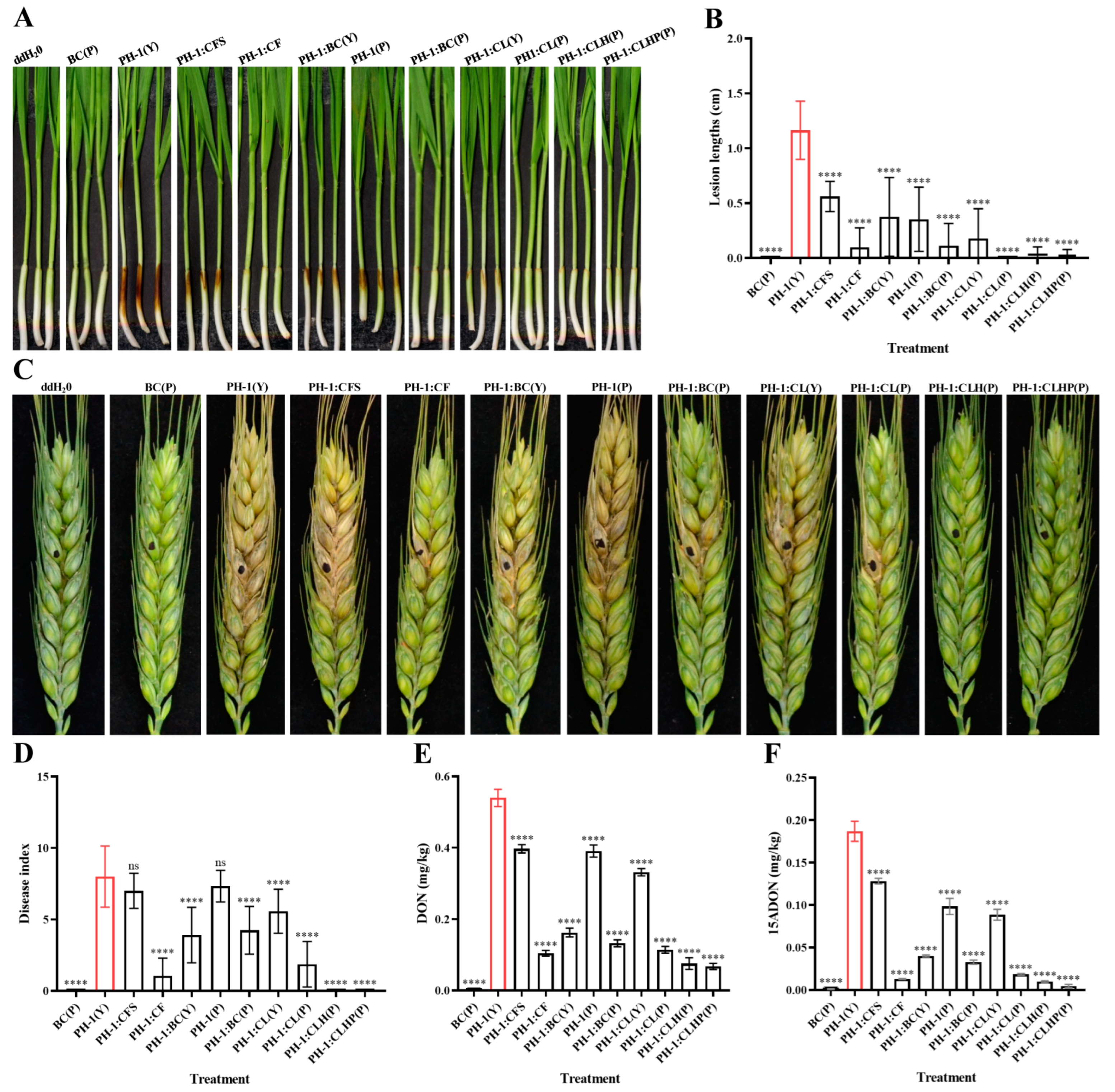
2.5. In Vitro Effect of Strain GSE25 on F. graminearum
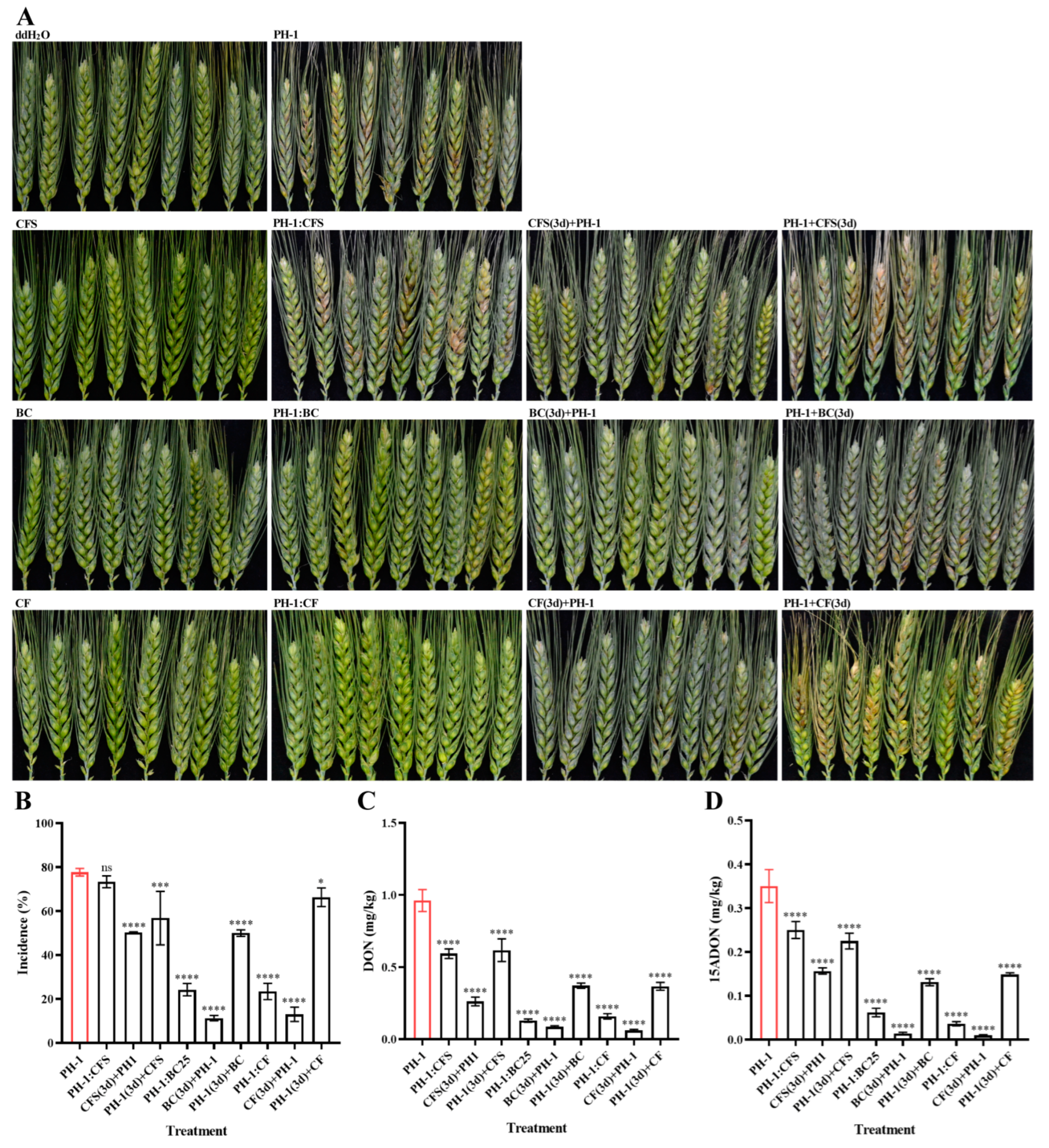
2.6. Biocontrol Effect of Strain GSE25 on FHB in Field Trials
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant, Fungal, and Bacterial Materials
5.2. Isolation and Identification of Endophytic Bacteria from Wheat Seeds
5.3. Screening of the Isolated Strains for Reducing F. graminearum PH-1 Pathogenicity and DON Production
5.4. Biosafety Assessed of Strain GSE25
5.5. Plant Pathogenicity Test of Strain GSE25
5.6. Whole Genome Sequence of Strain GSE25
5.7. Phylogenetic Tree Analysis of Strain GSE25
5.8. DNA-seq and RNA-seq Analyses of Inhibitory Effects of Strain GSE25 on F. graminearum PH-1
5.9. In Vitro Antifungal Activity of Strain GSE25 against F. graminearum
5.10. Biocontrol Effect of Strain GSE25 on FHB under Field Conditions
5.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal–Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, D.; Cai, P.; Lin, H.; Ying, H.; Hu, Q.; Wu, A. Elimination of Fusarium mycotoxin deoxynivalenol (DON) via microbial and enzymatic strategies: Current status and future perspectives. Trend Food Sci. Technol. 2022, 124, 96–107. [Google Scholar] [CrossRef]
- Bissonnette, K.M.; Kolb, F.L.; Ames, K.A.; Bradley, C.A. Effect of Fusarium head blight management practices on mycotoxin contamination of wheat straw. Plant Dis. 2018, 102, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.N.; Léon, J.; Naz, A.A.; Ballvora, A. Genetics and genomics of root system variation in adaptation to drought stress in cereal crops. J. Exp. Bot. 2021, 72, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, F.M.; Ameye, M.; De Saeger, S.; Audenaert, K.; Haesaert, G. Biological control of mycotoxigenic fungi and their toxins: An update for the pre-harvest approach. In Mycotoxins—Impact and Management Strategies; IntechOpen: London, UK, 2019; pp. 59–89. [Google Scholar]
- Egamberdieva, D.; Wirth, S.J.; Shurigin, V.V.; Hashem, A.; Abd Allah, E.F. Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Front. Microbiol. 2017, 8, 1887. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Zhang, P.; Xie, G.; Wang, L.; Xing, Y. Bacillus velezensis BY6 promotes growth of poplar and improves resistance contributing to the biocontrol of Armillaria solidipes. Microorganisms 2022, 10, 2472. [Google Scholar] [CrossRef]
- Chen, Q.; Qiu, Y.; Yuan, Y.; Wang, K.; Wang, H. Biocontrol activity and action mechanism of Bacillus velezensis strain SDTB038 against Fusarium crown and root rot of tomato. Front. Microbiol. 2022, 13, 994716. [Google Scholar] [CrossRef]
- Ibrahim, E.; Nasser, R.; Hafeez, R.; Ogunyemi, S.O.; Abdallah, Y.; Khattak, A.A.; Shou, L.; Zhang, Y.; Ahmed, T.; Atef Hatamleh, A.; et al. Biocontrol efficacy of endophyte Pseudomonas poae to alleviate fusarium seedling bight by refining the morpho-physiological attributes of wheat. Plants 2023, 12, 2277. [Google Scholar] [CrossRef]
- Yan, H.; Qiu, Y.; Yang, S.; Wang, Y.; Wang, K.; Jiang, L.; Wang, H. Antagonistic activity of Bacillus velezensis SDTB038 against phytophthora infestans in potato. Plant Dis. 2021, 105, 1738–1747. [Google Scholar] [CrossRef]
- Dobrzyński, J.; Jakubowska, Z.; Kulkova, I.; Kowalczyk, P.; Kramkowski, K. Biocontrol of fungal phytopathogens by Bacillus pumilus. Front. Microbiol. 2023, 14, 1194606. [Google Scholar] [CrossRef]
- Rishad, K.S.; Rebello, S.; Shabanamol, P.S.; Jisha, M.S. Biocontrol potential of halotolerant bacterial chitinase from high yielding novel Bacillus pumilus MCB-7 autochthonous to mangrove ecosystem. Pestic. Biochem. Physiol. 2017, 137, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Taheri, E.; Tarighi, S.; Taheri, P. An endophytic bacterium with biocontrol activity against important wheat pathogens. Biol. Control 2023, 183, 105243. [Google Scholar] [CrossRef]
- Deng, S.; Liu, Y.; Deng, Z.; Huang, Y. Isolation of actinobacterial endophytes from wheat sprouts as biocontrol agents to control seed pathogenic fungi. Arch. Microbiol. 2010, 203, 6163–6171. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.D.; Grossi, C.; Zawoznik, M.; Groppa, M.D. Wheat seeds harbour bacterial endophytes with potential as plant growth promoters and biocontrol agents of Fusarium graminearum. Microbiol. Res. 2016, 186, 37–43. [Google Scholar] [CrossRef]
- Diabankana, R.G.C.; Afordoanyi, D.M.; Safin, R.I.; Nizamov, R.M.; Karimova, L.Z.; Validov, S.Z. Antifungal properties, abiotic stress resistance, and biocontrol ability of Bacillus mojavensis PS17. Curr. Microbiol. 2021, 78, 3124–3132. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Huang, C.H.; Adi, V.S.K.; Huang, C.W.; Cheng, Y.I.; Chen, J.H.; Liu, Y.C. A statistical approach to identify prevalent virulence factors responsible for post-weaning diarrhoeic piglets. Vet. Med. 2022, 67, 430–439. [Google Scholar] [CrossRef]
- Vergara, A.; Normanno, G.; Di Ciccio, P.; Pedonese, F.; Nuvoloni, R.; Parisi, A.; Santagada, G.; Colagiorgi, A.; Zanardi, E.; Ghidini, S.; et al. Biofilm formation and its relationship with the molecular characteristics of food-related methicillin-resistant Staphylococcus aureus (MRSA). J. Food Sci. 2017, 82, 2364–2370. [Google Scholar] [CrossRef]
- Kizheva, Y.; Georgiev, G.; Donchev, D.; Nizamov, R.M.; Karimova, L.Z.; Validov, S.Z. Cross-over pathogenic bacteria detected in infected tomatoes (Solanum lycopersicum L.) and peppers (Capsicum annuum L.) in Bulgaria. Pathogens 2022, 11, 1507. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Alnajar, S.; Gupta, R.S. Phylogenomics and comparative genomic studies delineate six main clades within the family Enterobacteriaceae and support the reclassification of several polyphyletic members of the family. Infect. Genet. Evol. 2017, 54, 108–127. [Google Scholar] [CrossRef]
- Fan, Y.; Niu, X.; Zhang, D. Analysis of the process and factors influencing microbial phosphine production. Environ. Sci. Pollut. Res. 2023, 30, 76595–76605. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.J.; Park, S.Y.; Kim, C.G. Acidic neutralization by indigenous bacteria isolated from abandoned mine areas. Appl. Sci. 2022, 12, 3324. [Google Scholar] [CrossRef]
- Redkar, A.; Sabale, M.; Zuccaro, A.; Di Pietro, A. Determinants of endophytic and pathogenic lifestyle in root colonizing fungi. Curr. Opin. Plant Biol. 2022, 67, 102226. [Google Scholar] [CrossRef] [PubMed]
- Franken, P.; Takken, F.L.W.; Rep, M. Transcript accumulation in a trifold interaction gives insight into mechanisms of biocontrol. New Phytol. 2019, 224, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Faris, J.D. Fusarium graminearum KP4-like proteins possess root growth-inhibiting activity against wheat and potentially contribute to fungal virulence in seedling rot. Fungal Genet. Biol. 2019, 123, 1–13. [Google Scholar] [CrossRef]
- Baldwin, T.K.; Urban, M.; Brown, N.; Hammond-Kosack, K.E. A role for topoisomerase I in Fusarium graminearum and F. culmorum pathogenesis and sporulation. Mol. Plant-Microbe Interact. 2010, 23, 566–577. [Google Scholar] [CrossRef]
- Van Nguyen, T.; Kröger, C.; Bönnighausen, J.; Schäfer, W.; Bormann, J. The ATF/CREB transcription factor Atf1 is essential for full virulence, deoxynivalenol production, and stress tolerance in the cereal pathogen Fusarium graminearum. Mol. Plant-Microbe Interact. 2013, 26, 1378–1394. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, S.; Zhang, Q.; Tao, Y.; Wang, C.; Xu, J.R. FgSKN7 and FgATF1 have overlapping functions in ascosporogenesis, pathogenesis and stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 1245–1260. [Google Scholar] [CrossRef]
- Song, X.; Li, H.; Zhang, J.; Song, B.; Huang, T.; Du, X.M.; Gong, A.D.; Liu, Y.K.; Feng, Y.N.; Agboola, R.S.; et al. Trehalose 6-phosphate phosphatase is required for development, virulence and mycotoxin biosynthesis apart from trehalose biosynthesis in Fusarium graminearum. Fungal Genet. Biol. 2014, 63, 24–41. [Google Scholar] [CrossRef]
- Min, K.; Lee, J.; Kim, J.; Kim, S.G.; Kim, Y.H.; Vogel, S.; Trail, F.; Lee, Y.W. A novel gene, ROA, is required for normal morphogenesis and discharge of ascospores in Gibberella zeae. Eukaryot. Cell 2010, 9, 1495–1503. [Google Scholar] [CrossRef]
- Gaffoor, I.; Brown, D.W.; Plattner, R.; Proctor, R.H.; Qi, W.; Trail, F. Functional analysis of the polyketide synthase genes in the filamentous fungus Gibberella zeae (anamorph Fusarium graminearum). Eukaryot. Cell 2005, 4, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, R.J.N.; Schütt, C.; Lund, B.W.; Staerk, D.; Nielsen, J.; Olsson, S.; Giese, H. Two novel classes of enzymes are required for the biosynthesis of aurofusarin in Fusarium graminearum. J. Biol. Chem. 2011, 286, 10419–10428. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Mionetto, A.; Tiscornia, S.; Bettucci, L. Endophytic bacteria from wheat grain as biocontrol agents of Fusarium graminearum and deoxynivalenol production in wheat. Mycotoxin Res. 2015, 31, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Ponts, N.; Pinson-Gadais, L.; Barreau, C.; Richard-Forget, F.; Ouellet, T. Exogenous H2O2 and catalase treatments interfere with Tri genes expression in liquid cultures of Fusarium graminearum. FEBS Lett. 2007, 581, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, Q.; Wang, K.; Brian, K.; Liu, C.; Gu, Y. Study of the antifungal activity of Bacillus vallismortis ZZ185 in vitro and identification of its antifungal components. Bioresour. Technol. 2010, 101, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Function and biosynthesis of cell wall α-1,3-glucan in fungi. J. Fungi 2017, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Pupakdeepan, W.; Termsung, N.; Ruangwong, O.U.; Kunasakdakul, K. Plant growth promoting and colonization of endophytic Streptomyces albus CINv1 against strawberry anthracnose. Horticulturae 2023, 9, 766. [Google Scholar] [CrossRef]
- Ling, L.; Luo, H.; Yang, C.; Wang, Y.; Cheng, W.; Pang, M.; Jiang, K. Volatile organic compounds produced by Bacillus velezensis L1 as a potential biocontrol agent against postharvest diseases of wolfberry. Front. Microbiol. 2022, 13, 987844. [Google Scholar] [CrossRef]
- Setlow, P. Spore germination. Curr. Opin. Microbiol. 2003, 6, 550–556. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, Y.; Ma, D.; Xu, W.; Zhou, J.; Yan, H.; Yang, L.; Yin, J. Screening, identification, and optimization of fermentation conditions of an antagonistic endophyte to wheat had blight. Agronomy 2019, 9, 476. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; Hinton, D.M. Potential for control of seedling blight of wheat caused by Fusarium graminearum and related species using the bacterial endophyte Bacillus mojavensis. Biocontrol Sci. Technol. 2007, 17, 81–94. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, L.; Goodwin, P.H.; Xia, M.; Zhang, J.; Wang, Q.; Liang, J.; Sun, R.; Wu, C.; Yang, L. Isolation, identification, and complete genome assembly of an endophytic Bacillus velezensis YB-130, potential biocontrol agent against Fusarium graminearum. Front. Microbiol. 2020, 11, 598285. [Google Scholar] [CrossRef]
- Wu, C.; Chen, H.; Yuan, M.; Zhang, M.; Abubakar, Y.S.; Chen, X.; Zhong, H.; Zheng, W.; Zheng, H.; Zhou, J. FgAP1σ is critical for vegetative growth, conidiation, virulence, and DON biosynthesis in Fusarium graminearum. J. Fungi 2023, 9, 145. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, N.; Yang, L.; Deng, Y.; Wang, J.; Song, S.; Lin, S.; Wu, A.; Zhou, Z.; Hou, J. Multi-mycotoxin analysis of animal feed and animal-derived food using LC–MS/MS system with timed and highly selective reaction monitoring. Anal. Bioanal. Chem. 2015, 407, 7359–7368. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Fan, Z.; Li, Y.; Li, Z.; Zhao, H.; Feng, Y.; Xue, G.; Cui, J.; Yan, C.; Gan, L.; et al. Roles of the Crp/Fnr family regulator ArcR in the hemolysis and biofilm of Staphylococcus aureus. Microorganisms 2023, 11, 1656. [Google Scholar] [CrossRef] [PubMed]
- Viçosa, G.N.; Botelho, C.V.; Botta, C.; Bertolino, M.; de Carvalho, A.F.; Nero, L.A.; Cocolin, L. Impact of co-cultivation with Enterococcus faecalis over growth, enterotoxin production and gene expression of Staphylococcus aureus in broth and fresh cheeses. Int. J. Food Microbiol. 2019, 308, 108291. [Google Scholar] [CrossRef]
- Segata, N.; Börnigen, D.; Morgan, X.C.; Huttenhower, C. PhyloPhlAn is a new method for improved phylogenetic and taxonomic placement of microbes. Nat. Commun. 2013, 4, 2304. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Closest Relative (Obtained from BLAST Search) | Identity (%) | Matched GenBank No. |
|---|---|---|---|
| GSE01 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE06 | Pseudescherichia vulneris strain ATCC 33821 | 99 | NR041927 |
| GSE07 | Enterobacter cancerogenus strain LMG 2693 | 99 | NR044977 |
| GSE08 | Enterobacter hormaechei subsp. strain 10–17 | 100 | NR126208 |
| GSE09 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE10 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE12 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE14 | Enterobacter bugandensis strain 247BMC | 99 | NR148649 |
| GSE15 | Klebsiella grimontii strain SB73 | 100 | NR159317 |
| GSE16 | Enterobacter cancerogenus strain LMG 2693 | 99 | NR116756 |
| GSE19 | Enterobacter asburiae strain JCM6051 | 98 | NR024640 |
| GSE25 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE26 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE28 | Pseudescherichia vulneris strain ATCC 33821 | 99 | NR041927 |
| GSE32 | Yokenella regensburgei strain CIP 105435 | 99 | NR104934 |
| GSE37 | Yokenella regensburgei strain JCM 2403 | 100 | NR112986 |
| GSE38 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE43 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| GSE44 | Pseudescherichia vulneris strain ATCC 33821 | 99 | NR041927 |
| GSE47 | Pseudescherichia vulneris strain NBRC 102420 | 100 | NR114080 |
| Statistics | Number |
|---|---|
| Clean reads | 287,299 |
| Average read length (bp) | 12,475 |
| N50 length (bp) | 18,768 |
| Genome length (bp) | 4,761,311 |
| Coding gene | 4486 |
| Total length (bp) | 4,208,967 |
| GC (%) | 55.6 |
| rRNA | 22 |
| tRNA | 85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, M.; Abdallah, M.F.; Song, M.; Xu, Y.; Sun, D.; Lu, P.; Wang, J. Novel Endophytic Pseudescherichia sp. GSE25 Strain Significantly Controls Fusarium graminearum and Reduces Deoxynivalenol in Wheat. Toxins 2023, 15, 702. https://doi.org/10.3390/toxins15120702
Gao M, Abdallah MF, Song M, Xu Y, Sun D, Lu P, Wang J. Novel Endophytic Pseudescherichia sp. GSE25 Strain Significantly Controls Fusarium graminearum and Reduces Deoxynivalenol in Wheat. Toxins. 2023; 15(12):702. https://doi.org/10.3390/toxins15120702
Chicago/Turabian StyleGao, Meiling, Mohamed F. Abdallah, Minggui Song, Yiqian Xu, Daiyuan Sun, Ping Lu, and Jianhua Wang. 2023. "Novel Endophytic Pseudescherichia sp. GSE25 Strain Significantly Controls Fusarium graminearum and Reduces Deoxynivalenol in Wheat" Toxins 15, no. 12: 702. https://doi.org/10.3390/toxins15120702