Cytotoxicity and Radiosensitizing Potentials of Pilosulin-3, a Recombinant Ant Venom, in Breast Cancer Cells

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Specificity of Pilosulin-3 Peptide
2.2. Cell Proliferation Using the Real-Time Cell Analyzer (RTCA)
2.3. Pilosulin-3 Effect on Cell Cycle Progression
2.4. Pilosulin-3 Effect on Apoptosis
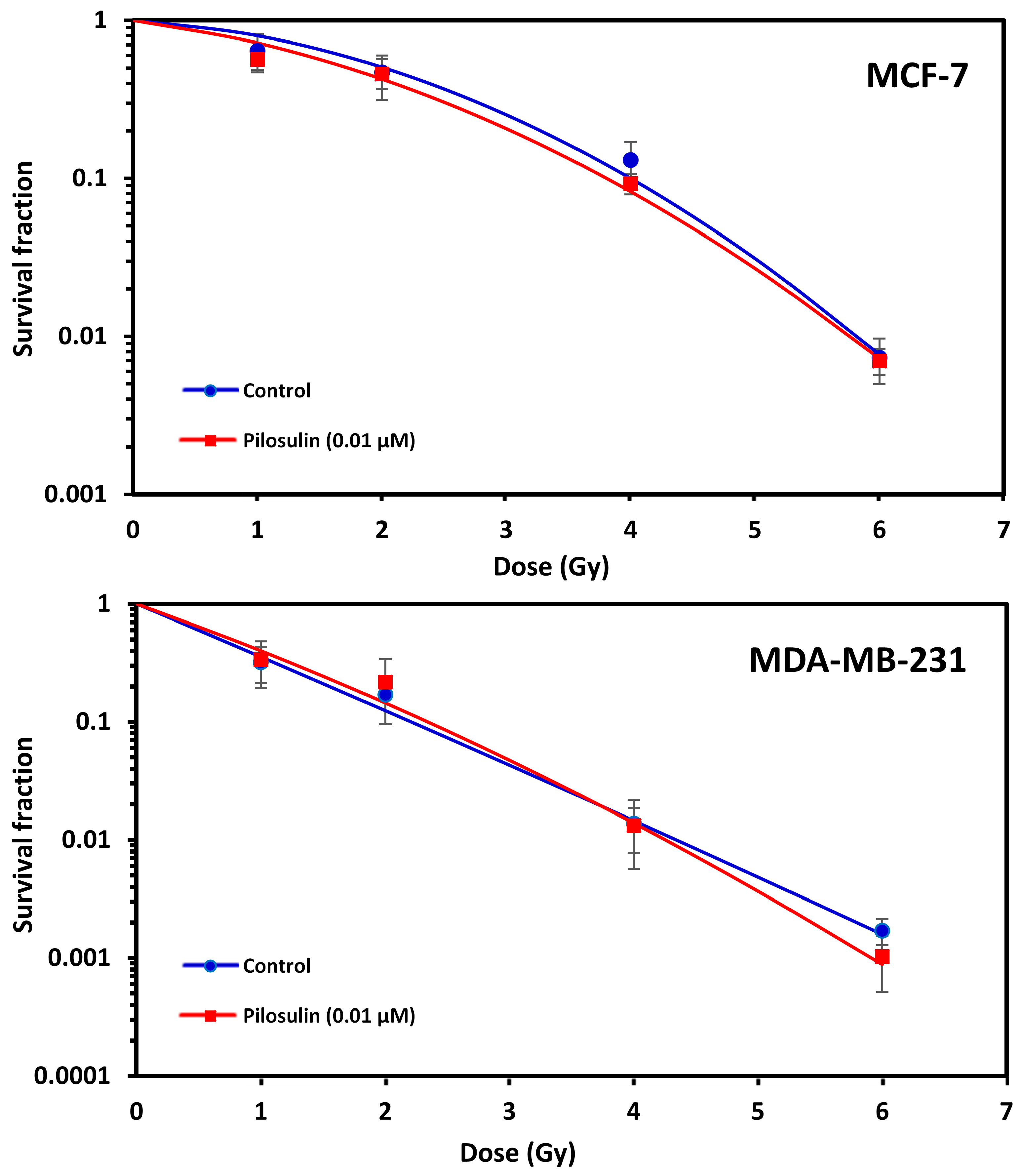
2.5. Clonogenic Cell Survival
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Pilosulin-3 Solution
5.2. Cell Culture
5.3. Real Time Cell Proliferation Assay
5.4. Cell Cycle Analysis
5.5. Cell Apoptosis Analysis
5.6. Clonogenic Survival Assay
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fouad, Y.A.; Aanei, C. Revisiting the hallmarks of cancer. Am. J. Cancer Res. 2017, 7, 1016–1036. [Google Scholar] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Hortobagyi, G.N. Breast Cancer: 45 Years of Research and Progress. J. Clin. Oncol. 2020, 38, 2454–2462. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K.K. Various types and management of breast cancer: An overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar] [PubMed]
- Sica, A. Role of tumour-associated macrophages in cancer-related inflammation. Exp. Oncol. 2010, 32, 153–158. [Google Scholar] [PubMed]
- Seabrooks, L.; Hu, L. Insects: An underrepresented resource for the discovery of biologically active natural products. Acta Pharm. Sinica. B 2017, 7, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Almeer, R.S.; Alarifi, S.; Alkahtani, S.; Ibrahim, S.R.; Ali, D.; Moneim, A. The potential hepatoprotective effect of royal jelly against cadmium chloride-induced hepatotoxicity in mice is mediated by suppression of oxidative stress and upregulation of Nrf2 expression. Biomed. Pharmacother. 2018, 106, 1490–1498. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B. Therapeutic arthropods and other, largely terrestrial, folk-medicinally important invertebrates: A comparative survey and review. J. Ethnobiol. Ethnomed. 2017, 13, 9. [Google Scholar] [CrossRef]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A.B. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, eaau4640. [Google Scholar] [CrossRef]
- Haight, K.L.; Tschinkel, W.R. Patterns of venom synthesis and use in the fire ant, Solenopsis invicta. Toxicon 2003, 42, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Street, M.D.; Donovan, G.R.; Baldo, B.A. Molecular cloning and characterization of the major allergen Myr p II from the venom of the jumper ant Myrmecia pilosula: Myr p I and Myr p II share a common protein leader sequence. Biochim. Et Biophys. Acta 1996, 1305, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Wanandy, T.; Gueven, N.; Davies, N.W.; Brown, S.G.; Wiese, M.D. Pilosulins: A review of the structure and mode of action of venom peptides from an Australian ant Myrmecia pilosula. Toxicon 2015, 98, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Guido-Patino, J.C.; Plisson, F. Profiling hymenopteran venom toxins: Protein families, structural landscape, biological activities, and pharmacological benefits. Toxicon X 2022, 14, 100119. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.D.; Brown, S.G.; Chataway, T.K.; Davies, N.W.; Milne, R.W.; Aulfrey, S.J.; Heddle, R.J. Myrmecia pilosula (Jack Jumper) ant venom: Identification of allergens and revised nomenclature. Allergy 2007, 62, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Wanandy, T.; Wilson, R.; Gell, D.; Rose, H.E.; Gueven, N.; Davies, N.W.; Brown, S.G.A.; Wiese, M.D. Towards complete identification of allergens in Jack Jumper (Myrmecia pilosula) ant venom and their clinical relevance: An immunoproteomic approach. Clin. Exp. Allergy 2018, 48, 1222–1234. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, H.; Akagi, M.; Imai, H.T.; Taylor, R.W.; Kubo, T. Molecular cloning and biological characterization of novel antimicrobial peptides, pilosulin 3 and pilosulin 4, from a species of the Australian ant genus Myrmecia. Arch. Biochem. Biophys. 2004, 428, 170–178. [Google Scholar] [CrossRef]
- Al-Shahwan, M.; Al-Khenaizan, S.; Al-Khalifa, M. Black (samsum) ant induced anaphylaxis in Saudi Arabia. Saudi Med. J. 2006, 27, 1761–1763. [Google Scholar]
- Alsharani, M.; Alanazi, M.; Alsalamah, M. Black ant stings caused by Pachycondyla sennaarensis: A significant health hazard. Ann. Saudi Med. 2009, 29, 207–211. [Google Scholar]
- Lockhart, G.J. Ants, and Other Great Medicines. Jacobson, A.L., Ed.; 2007. Available online: https://arthurleej.com/Ants.pdf (accessed on 1 January 2023).
- Wanandy, T.; Mulcahy, E.; Lau, W.Y.; Brown, S.G.A.; Wiese, M.D. Global View on Ant Venom Allergy: From Allergenic Components to Clinical Management. Clin. Rev. Allergy Immunol. 2022, 62, 123–144. [Google Scholar] [CrossRef]
- Ma, R.; Mahadevappa, R.; Kwok, H.F. Venom-based peptide therapy: Insights into anti-cancer mechanism. Oncotarget 2017, 8, 100908–100930. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (Review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Fekri, H.S.; Hashemi, F.; Hushmandi, K.; Mohammadinejad, R.; Ashrafizadeh, M.; Zarrabi, A.; Garg, M. Venom peptides in cancer therapy: An updated review on cellular and molecular aspects. Pharmacol. Res. 2021, 164, 105327. [Google Scholar] [CrossRef] [PubMed]
- Walle, T.; Martinez Monge, R.; Cerwenka, A.; Ajona, D.; Melero, I.; Lecanda, F. Radiation effects on antitumor immune responses: Current perspectives and challenges. Ther. Adv. Med. Oncol. 2018, 10, 1758834017742575. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B. Animal Venoms have Potential to Treat Cancer. Curr. Top. Med. Chem. 2018, 18, 2555–2566. [Google Scholar] [CrossRef] [PubMed]
- Alsbeih, G.; Al-Meer, R.S.; Al-Harbi, N.; Bin Judia, S.; Al-Buhairi, M.; Venturina, N.Q.; Moftah, B. Gender bias in individual radiosensitivity and the association with genetic polymorphic variations. Radiother. Oncol. 2016, 119, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.X.; King, M.A.; Donovan, G.R.; Alewood, D.; Alewood, P.; Sawyer, W.H.; Baldo, B.A. Cytotoxicity of pilosulin 1, a peptide from the venom of the jumper ant Myrmecia pilosula. Biochim. Et Biophys. Acta 1998, 1425, 74–80. [Google Scholar] [CrossRef] [PubMed]
- King, M.A.; Wu, Q.X.; Donovan, G.R.; Baldo, B.A. Flow cytometric analysis of cell killing by the jumper ant venom peptide pilosulin 1. Cytometry 1998, 32, 268–273. [Google Scholar] [CrossRef]
- Arbiser, J.L.; Kau, T.; Konar, M.; Narra, K.; Ramchandran, R.; Summers, S.A.; Vlahos, C.J.; Ye, K.; Perry, B.N.; Matter, W.; et al. Solenopsin, the alkaloidal component of the fire ant (Solenopsis invicta), is a naturally occurring inhibitor of phosphatidylinositol-3-kinase signaling and angiogenesis. Blood 2007, 109, 560–565. [Google Scholar] [CrossRef]
- Achmad, M.H.; Horax, S.; Ramadhany, S.; Rieuwpassa, I.E.; Sari, M.; Handayani, H.; Singgih, M.F.; Sugiharto, S. Anti-Cancer and Anti-Proliferation Activity of Ethyl Asetat Extract From Ant Nest (Myrmecodia pendans) in Burkitt’s Lymphoma Cancer Cells. Pesqui. Bras. Odontopediatria Clínica Integr. 2019, 19, e4325. [Google Scholar] [CrossRef]
- Borojeni, S.K.; Zolfagharian, H.; Babaie, M.; Javadi, I. Cytotoxic Effect of Bee (A. mellifera) Venom on Cancer Cell Lines. J. Pharmacopunct. 2020, 23, 212–219. [Google Scholar] [CrossRef]
- Donovan, G.R.; Baldo, B. Method of Cell Inhibition Using Polypeptides Derived from the Venom of the Austrialian Jumper ant Myrmecia pilosula. Patent# US 6,294,649 B1, 2001. [Google Scholar]
- Robles-Loaiza, A.A.; Pinos-Tamayo, E.A.; Mendes, B.; Ortega-Pila, J.A.; Proano-Bolanos, C.; Plisson, F.; Teixeira, C.; Gomes, P.; Almeida, J.R. Traditional and Computational Screening of Non-Toxic Peptides and Approaches to Improving Selectivity. Pharmaceuticals 2022, 15, 323. [Google Scholar] [CrossRef]
- Timmons, P.B.; Hewage, C.M. ENNAACT is a novel tool which employs neural networks for anticancer activity classification for therapeutic peptides. Biomed. Pharmacother. 2021, 133, 111051. [Google Scholar] [CrossRef]
- Badr, G.; Garraud, O.; Daghestani, M.; Al-Khalifa, M.S.; Richard, Y. Human breast carcinoma cells are induced to apoptosis by samsum ant venom through an IGF-1-dependant pathway, PI3K/AKT and ERK signaling. Cell Immunol. 2012, 273, 10–16. [Google Scholar] [CrossRef]
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal protein toxins: Origins and therapeutic applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef]
- Roshan Moniri, M.; Young, A.; Reinheimer, K.; Rayat, J.; Dai, L.J.; Warnock, G.L. Dynamic assessment of cell viability, proliferation and migration using real time cell analyzer system (RTCA). Cytotechnology 2015, 67, 379–386. [Google Scholar] [CrossRef]
- Hamidi, H.; Lilja, J.; Ivaska, J. Using xCELLigence RTCA Instrument to Measure Cell Adhesion. Bio-Protoc 2017, 7, e2646. [Google Scholar] [CrossRef]
- Kim, K.H.; Sederstrom, J.M. Assaying Cell Cycle Status Using Flow Cytometry. Curr. Protoc. Mol. Biol. 2015, 111, 28.6.1–28.6.11. [Google Scholar] [CrossRef] [PubMed]
- Cummings, B.S.; Schnellmann, R.G. Measurement of cell death in mammalian cells. Curr. Protoc. Pharmacol. 2004, 12, 12.8.1–12.8.22. [Google Scholar] [CrossRef] [PubMed]
- Salem, S.D.; Abou-Tarboush, F.M.; Saeed, N.M.; Al-Qadasi, W.D.; Farah, M.A.; Al-Buhairi, M.; Al-Harbi, N.; Alhazza, I.; Alsbeih, G. Involvement of p53 in gemcitabine mediated cytotoxicity and radiosensitivity in breast cancer cell lines. Gene 2012, 498, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Rafehi, H.; Orlowski, C.; Georgiadis, G.T.; Ververis, K.; El-Osta, A.; Karagiannis, T.C. Clonogenic assay: Adherent cells. J. Vis. Exp. 2011, 49, e2573. [Google Scholar] [CrossRef]
- Fertil, B.; Dertinger, H.; Courdi, A.; Malaise, E.P. Mean inactivation dose: A useful concept for intercomparison of human cell survival curves. Radiat. Res. 1984, 99, 73–84. [Google Scholar] [CrossRef]
- Minafra, L.; Porcino, N.; Bravata, V.; Gaglio, D.; Bonanomi, M.; Amore, E.; Cammarata, F.P.; Russo, G.; Militello, C.; Savoca, G.; et al. Radiosensitizing effect of curcumin-loaded lipid nanoparticles in breast cancer cells. Sci. Rep. 2019, 9, 11134. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alzeer, R.M.; Al-Hadyan, K.S.; Al-Harbi, N.M.; Bin Judia, S.S.; Almeer, R.S.; Alsbeih, G.A. Cytotoxicity and Radiosensitizing Potentials of Pilosulin-3, a Recombinant Ant Venom, in Breast Cancer Cells. Toxins 2023, 15, 701. https://doi.org/10.3390/toxins15120701
Alzeer RM, Al-Hadyan KS, Al-Harbi NM, Bin Judia SS, Almeer RS, Alsbeih GA. Cytotoxicity and Radiosensitizing Potentials of Pilosulin-3, a Recombinant Ant Venom, in Breast Cancer Cells. Toxins. 2023; 15(12):701. https://doi.org/10.3390/toxins15120701
Chicago/Turabian StyleAlzeer, Reema M., Khaled S. Al-Hadyan, Najla M. Al-Harbi, Sara S. Bin Judia, Rafa S. Almeer, and Ghazi A. Alsbeih. 2023. "Cytotoxicity and Radiosensitizing Potentials of Pilosulin-3, a Recombinant Ant Venom, in Breast Cancer Cells" Toxins 15, no. 12: 701. https://doi.org/10.3390/toxins15120701