Different Resistance to DON versus HT2 + T2 Producers in Nordic Oat Varieties

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
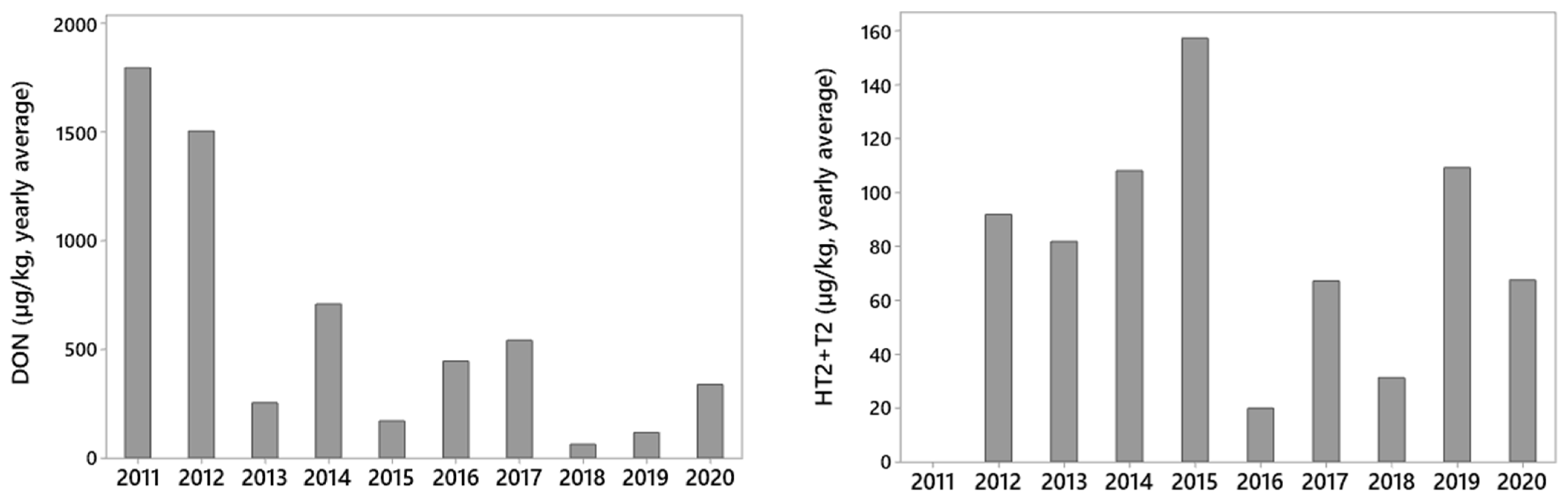
2.1. Variation in Mycotoxin Levels between Years
2.2. Ranking of Oat Varieties According to Mycotoxin Content in Harvested Grain
2.3. Ranking of Oat Varieties According to Fungal DNA Levels in Harvested Grain
2.4. Ranking of Oat Varieties According to Mycotoxin versus Fungal DNA Levels
2.5. Factors Associated with Mycotoxin Content
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Field Trials
5.2. Grain Samples
5.3. Mycotoxin Analysis of Harvested Grains
5.4. Fungal DNA Content of Harvested Grains
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khodaei, D.; Javanmardi, F.; Khaneghah, A.M. The global overview of the occurrence of mycotoxins in cereals: A three-year survey. Curr. Opin. Food Sci. 2021, 39, 36–42. [Google Scholar] [CrossRef]
- Langseth, W.; Elen, O.N. Differences between barley, oats and wheat in the occurrence of deoxynivalenol and other trichothecenes in Norwegian grain. J. Phytopathol. 1996, 144, 113–118. [Google Scholar] [CrossRef]
- Langseth, W.; Elen, O.N. The occurence of deoxynivalenol in Norwegian cereals- differences between years and districts, 1988-1996. Acta Agric. Scand. B Soil Plant Sci. 1997, 47, 176–184. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Aamot, H.U.; Torp, T.; Jestoi, M.; Lattanzio, V.M.T.; Klemsdal, S.S.; Waalwijk, C.; van der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers’ fields in Norway over a six-year period. World Mycotoxin J. 2016, 9, 365–378. [Google Scholar] [CrossRef]
- Bernhoft, A.; Eriksen, G.S.; Sundheim, L.; Berntssen, M.; Brantsæter, L.A.; Brodal, G.; Fæste, C.K.; Hofgaard, I.S.; Rafoss, T.; Sivertsen, T.; et al. Risk Assessment of Mycotoxins in Cereal Grain in Norway; VKM Report no 2013:21. Norwegian Scientific Committee for Food Safety: Oslo, Norway, 2013; pp. 1–287. Available online: https://vkm.no/english/riskassessments/allpublications/riskassessmentofmycotoxinsincerealgraininnorway.4.27ef9ca915e07938c3b2bad8.html (accessed on 29 March 2022).
- Bernhoft, A.; Clasen, P.E.; Kristoffersen, A.B.; Torp, M. Less Fusarium infestation and mycotoxin contamination in organic than in conventional cereals. Food Addit. Contam. Part A 2010, 27, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Kosiak, B.; Torp, M.; Skjerve, E.; Thrane, U. The prevalence and distribution of Fusarium species in Norwegian cereals: A survey. Acta Agric. Scand.-Sect. B-Soil Plant Sci. 2003, 53, 168–176. [Google Scholar] [CrossRef]
- Kim, M.O. Fusarium and Microdochium on Cereal Seeds—Survival during Storage and Impact on Germination. Master’s Thesis, Norwegian University of Life Sciences, Ås Municipality, Norway, 2014. [Google Scholar]
- Torp, M.; Nirenberg, H.I. Fusarium langsethiae sp. nov. on cereals in Europe. Int. J. Food Microbiol. 2004, 95, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Divon, H.H.; Razzaghian, J.; Aamot, H.U.; Klemsdal, S.S. Fusarium langsethiae (Torp and Nirenberg), investigation of alternative infection routes in oats. Eur. J. Plant Pathol. 2012, 132, 147–161. [Google Scholar] [CrossRef]
- Opoku, N.; Back, M.; Edwards, S.G. Development of Fusarium langsethiae in commercial cereal production. Eur. J. Plant Pathol. 2013, 136, 159–170. [Google Scholar] [CrossRef]
- Knutsen, A.K.; Torp, M.; Holst-Jensen, A. Phylogenetic analyses of the Fusarium poae, F. sporotrichioides and F. langsethiae species complex based on partial sequences of the translation elongation factor-1 alpha gene. Int. J. Food Microbiol. 2004, 95, 287–295. [Google Scholar] [CrossRef]
- Thrane, U.; Adler, A.; Clasen, P.E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Gagkaeva, T.Y.; Orina, A.S.; Gavrilova, O.P.; Gogina, N.N. Evidence of Microdochium Fungi Associated with Cereal Grains in Russia. Microorganisms 2020, 8, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwan, M.; Cockerell, V. Identification of the Microdochium species causing reduced seedling emergence in spring oat and barley seed. In Proceedings of the Dundee Conference: Crop Protection in Northern Britain, Dundee, UK, 23–24 February 2016; pp. 105–110. [Google Scholar]
- European Commission. COMMISSION REGULATION (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Off. J. Eur. Union 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:02006R1881-20170728&qid=1512388911628&from=EN (accessed on 29 March 2022).
- Tekle, S.; Lillemo, M.; Skinnes, H.; Reitan, L.; Buraas, T.; Bjørnstad, Å. Screening of Oat Accessions for Fusarium Head Blight Resistance Using Spawn-Inoculated Field Experiments. Crop Sci. 2018, 58, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit. Contam. Part A 2009, 26, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredlund, E.; Gidlund, A.; Pettersson, H.; Olsen, M.; Börjesson, T. Real-time PCR detection of Fusarium species in Swedish oats and correlation to T-2 and HT-2 toxin content. World Mycotoxin J. 2010, 3, 77–88. [Google Scholar] [CrossRef]
- Kaukoranta, T.; Hietaniemi, V.; Rämö, S.; Koivisto, T.; Parikka, P. Contrasting responses of T-2, HT-2 and DON mycotoxins and Fusarium species in oat to climate, weather, tillage and cereal intensity. Eur. J. Plant Pathol. 2019, 115, 93–110. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Madden, L.V.; Edwards, S. Modelling the effects of environmental conditions on HT2 andT2 toxin accumulation in field oat grains. Phytopathology 2014, 104, 57–66. [Google Scholar] [CrossRef]
- Hjelkrem, A.G.R.; Torp, T.; Brodal, G.; Aamot, H.U.; Strand, E.; Nordskog, B.; Dill-Macky, R.; Edwards, S.G.; Hofgaard, I.S. DON content in oat grains in Norway related to weather conditions at different growth stages. Eur. J. Plant Pathol. 2017, 148, 577–594. [Google Scholar] [CrossRef] [Green Version]
- Hjelkrem, A.G.R.; Aamot, H.U.; Brodal, G.; Strand, E.C.; Torp, T.; Edwards, S.G.; Dill-Macky, R.; Hofgaard, I.S. HT-2 and T-2 toxins in Norwegian oat grains related to weather conditions at different growth stages. Eur. J. Plant Pathol. 2018, 151, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Elen, O.; Hofgaard, I.S.; Brodal, G.; Aamot, H.U.; Jestoi, M.; Klemsdal, S.S. Effect of Fungicide Treatment in field trials on different Fusarium spp. and mycotoxins. In Proceedings of the 10th International Fusarium Workshop and Fusarium genomics Workshop, Alghero, Italy, 30 August–2 September 2008; p. 72. [Google Scholar]
- European Food Safety Authority (EFSA); Arcella, D.; Gergelova, P.; Innocenti, M.L.; Steinkellner, H. Human and animal dietary exposure to T-2 and HT-2 toxin. EFSA J. 2017, 15, 57. [Google Scholar] [CrossRef]
- European Commission. COMMISSION RECOMMENDATION of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products (2013/165/EC). Off. J. Eur. Union 2013, 91, 12–15. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32013H0165&qid=1425464527839&from=EN (accessed on 29 March 2022).
- van Eeuwijk, F.A.; Mesterhazy, A.; Kling, C.; Ruckenbauer, P.; Saur, L.; Bürstmayr, H.; Lemmens, M.; Keizer, L.C.P.; Maurin, N.; Snijders, C.H.A. Assessing non-specificity of resistance in wheat to head blight caused by inoculation with European strains of Fusarium culmorum, F. graminearum and F. nivale using a multiplicative model for interaction. Theor. Appl. Genet. 1995, 90, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A. Updating the Breeding Philosophy of Wheat to Fusarium Head Blight (FHB): Resistance Components, QTL Identification, and Phenotyping—A Review. Plants 2020, 9, 1702. [Google Scholar] [CrossRef]
- Audenaert, K.; Broeck, R.; Bekaert, B.; Witte, F.; Heremans, B.; Messens, K.; Häfte, M.; Haesaert, G. Fusarium head blight (FHB) in Flanders: Population diversity, inter-species associations and DON contamination in commercial winter wheat varieties. Eur. J. Plant Pathol. 2009, 125, 445–458. [Google Scholar] [CrossRef]
- Herrmann, M.H.; Hautsalo, J.; Georgieva, P.; Bund, A.; Winter, M.; Beuch, S. Relationship between genetic variability of flowering traits and Fusarium mycotoxin contamination in oats. Crop Sci. 2020, 60, 852–862. [Google Scholar] [CrossRef] [Green Version]
- Aureli, G.; Amoriello, T.; Belocchi, A.; D’Egidio, M.G.; Fornara, M.; Melloni, S.; Quaranta, F. Preliminary survey on the co-occurrence of DON and T2+HT2 toxins in durum wheat in Italy. Cereal Res. Commun. 2015, 43, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Tekauz, A.; McCallum, B.; Ames, N.; Fetch, J.M. Fusarium head blight of oat-current status in western Canada. Can. J. Plant Pathol. 2004, 26, 473–479. [Google Scholar] [CrossRef]
- Aamot, H.U.; Mousavi, H.; Razzaghian, J.; Brodal, G.; Sulyok, M.; Krska, R.; Edwards, S.G.; Hofgaard, I.S. Fusarium langsethiae and mycotoxin contamination in oat grain differed with growth stage at inoculation. Toxins 2022. submitted for publication. [Google Scholar]
- Russenes, A.L.; Abrahamsen, U.; Tangsveen, J.; Kufoalor, B. Sorter og sortsprøving 2020. In Jord og Plantekultur 2021. Forsøk i korn, olje-og Belgvekster, Engføavl og Potet 2020; Strand, E., Ed.; NIBIO bok; Norwegian Institute of Bioeconomy: Ås, Norway, 2021; Volume 7, pp. 28–62. [Google Scholar]
- Nazari, L.; Pattori, E.; Terzi, V.; Morcia, C.; Rossi, V. Influence of temperature on infection, growth, and mycotoxin production by Fusarium langsethiae and F. sporotrichioides in durum wheat. Food Microbiol. 2014, 39, 19–26. [Google Scholar] [CrossRef]
- Lillemo, M.; Sørensen, E.; Almvik, M.; Aamot, H.U.; Hofgaard, I.S. Development of a reliable field-testing methodology for Fusarium langsethiae resistance in oats confirms F. langsethiae-specific susceptibility in certain genotypes with otherwise good FHB resistance. In Proceedings of the 72nd Annual Plant Breeders Conference, Saatgut, Austria, 22 November 2021. [Google Scholar]
- Hofgaard, I.S.; Seehusen, T.; Aamot, H.U.; Riley, H.; Razzaghian, J.; Le, V.H.; Hjelkrem, A.G.R.; Dill-Macky, R.; Brodal, G. Inoculum potential of Fusarium spp. relates to tillage and straw management in Norwegian fields of spring oats. Front. Microbiol. 2016, 7, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodal, G.; Aamot, U.H.; Almvik, M.; Hofgaard, S.I. Removal of Small Kernels Reduces the Content of Fusarium mycotoxins in Oat Grain. Toxins 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Hautsalo, J.; Jauhiainen, L.; Hannukkala, A.; Manninen, O.; Veteläinen, M.; Pietilä, L.; Peltoniemi, K.; Jalli, M. Resistance to Fusarium head blight in oats based on analyses of multiple field and greenhouse studies. Eur. J. Plant Pathol. 2020, 158, 15–33. [Google Scholar] [CrossRef]
- Bergo, K. Enda et år med lite DON. Norsk Landbruk; Oslo, Norway. Available online: https://www.norsklandbruk.no/plantekultur/enda-et-ar-med-lite-don/ (accessed on 29 March 2022).
- Bernhoft, A.; Christensen, E.; Er, C.; Plassen, C.; Eriksen, G.; Tukun, F.-L. The Surveillance Programme for Feed and Feed Materials in Norway 2020—Mycotoxins and Fungi; The Norwegian Veterinary Institute: Oslo, Norway, 2021; pp. 1–26. Available online: https://www.vetinst.no/en/surveillance-programmes/feed-safety (accessed on 29 March 2022).
- Hjelkrem, A.G.; Aamot, H.U.; Lillemo, M.; Sørensen, E.S.; Brodal, G.; Russenes, A.L.; Edwards, S.G.; Hofgaard, I.S. Weather Patterns Associated with DON Levels in Norwegian Spring Oat Grain: A Functional Data Approach. Plants 2022, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Hole, H.; Eikemo, H.; Nordskog, B. Været i Vekstsesongen 2014; Bioforsk Tema; Bioforsk: Sterhdm, Norway, 2014; p. 29. [Google Scholar]
- Söderström, M.; Börjesson, T.; Roland, B.; Stadig, H. Modelling within-field variations in deoxynivalenol (DON) content in oats using proximal and remote sensing. Precis. Agric. 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Klötzel, M.; Lauber, U. New SPE Sorbent for Clean-Up of Fusarium Toxin-Contaminated Cereals & Cereal-Based Foods, Bond Elut Mycotoxin. 2017, pp. 1–5. Available online: https://www.agilent.com/cs/library/applications/SI-00295.pdf (accessed on 15 January 2018).
- Elbers, I.J.W.; Pereboom, D.P.K.H.; Mol, J.G.J.; de Nijs, W.C.M. Proficiency Test for Mycotoxins in Oat Meal; Wageningen Food Safety Research: Wageningen, The Netherlands, 2019. [Google Scholar]
- Aamot, H.U.; Lysøe, E.; Koga, S.; Nielsen, K.A.G.; Böcker, U.; Brodal, G.; Dill-Macky, R.; Uhlen, A.K.; Hofgaard, I.S. Microdochium majus and other fungal pathogens associated with reduced gluten quality in wheat grain. Int. J. Food Microbiol. 2020, 331, 108712. [Google Scholar] [CrossRef]
- Koga, S.; Aamot, H.U.; Uhlen, A.K.; Seehusen, T.; Veiseth-Kent, E.; Hofgaard, I.S.; Moldestad, A.; Böcker, U. Environmental factors associated with glutenin polymer assembly during grain maturation. J. Cereal Sci. 2020, 91, 102865. [Google Scholar] [CrossRef]
- Halstensen, A.S.; Nordby, K.C.; Eduard, W.; Klemsdal, S.S. Real-time PCR detection of toxigenic Fusarium in airborne and settled grain dust and associations with trichothecene mycotoxins. J. Environ. Monit. 2006, 8, 1235–1241. [Google Scholar] [CrossRef]
- Waalwijk, C.; van der Heide, R.; de Vries, I.; van der Lee, T.; Schoen, C.; Costrel-de Corainville, G.; Häuser-Hahn, I.; Kastelein, P.; Köhl, J.; Lonnet, P.; et al. Quantitative detection of Fusarium species in wheat using TaqMan. Eur. J. Plant Pathol. 2004, 110, 481–494. [Google Scholar] [CrossRef]
- Waalwijk, C.; Köhl, J.; de Vries, P.M.; van der Lee, T. Fusarium in Wintertarwe (2007 en 2008). Inventarisatie, Identificatie en Kwantificering van Nieuwe Soorten; 272; Wageningen Plant Research International: Wageningen, The Netherlands, 2009. [Google Scholar]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Jestoi, M.; Parikka, P.; Hietaniemi, V.; Gagkaeva, T.; Sarlin, T.; Haikara, A.; Laaksonen, S.; Rizzo, A. Real-time PCR detection and quantification of Fusarium poae, F. graminearum, F. sporotrichioides and F. langsethiae in cereal grains in Finland and Russia. Arch. Phytopathol. Plant Prot. 2008, 41, 243–260. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Justesen, A.F.; Jensen, J.D.; Jørgensen, L.N. Microdochium nivale and Microdochium majus in seed samples of Danish small grain cereals. Crop Prot. 2013, 43, 192–200. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oat Variety 1 | FaDNA (18) 2,3 | FcFgDNA (10) | FgDNA (10) | FlDNA (19) | FpDNA (16) | MmDNA (8) | MnDNA (7) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Avetron | 84 | B | 34 | AB | 33 | AB | 419 | BCD | 51 | AB | 118 | A | 21 | C |
| Belinda | 194 | A | 151 | A | 123 | A | 586 | AB | 76 | AB | 240 | A | 420 | A |
| Haga | 164 | AB | 75 | AB | 57 | AB | 464 | BC | 115 | A | 233 | A | 77 | BC |
| Odal | 123 | AB | 31 | B | 31 | AB | 889 | A | 78 | AB | 186 | A | 35 | BC |
| Ringsaker | 135 | AB | 28 | B | 23 | B | 453 | BCD | 101 | AB | 108 | A | 63 | BC |
| Vinger | 134 | AB | 36 | AB | 35 | AB | 332 | CD | 46 | AB | 105 | A | 80 | BC |
| Våler | 135 | AB | 36 | AB | 36 | AB | 278 | D | 42 | B | 203 | A | 115 | AB |
| P 4 | 0.06 | 0.01 | 0.03 | <0.001 | 0.01 | 0.02 | <0.001 | |||||||
| Target Species 1 | Ref. 2 | Primers, Probe | Primer/Probe Sequences |
|---|---|---|---|
| F. avenaceum, F. arthrosporoides | [49] | TMAVf | AGATCGGACAATGGTGCATTATAA |
| TMAVr | GGCCCTACTATTTACTCTTGCTTTTG | ||
| TMAVp | Cyanine5-CTCCTGAGAGGTCCCAGAGATGAACATAACTTC-BHQ3 | ||
| F. culmorum | [50] | culmorum MGB-F | TCACCCAAGACGGGAATGA |
| culmorum MGB-R | GAACGCTGCCCTCAAGCTT | ||
| culmorum MGB pr | 6FAM-CACTTGGATATATTTCC-MGBNFQ | ||
| F. graminearum | [50] | graminearum MGB-F | GGCGCTTCTCGTGAACACA |
| graminearum MGB-R | TGGCTAAACAGCACGAATGC | ||
| graminearum MGB pr | 6FAM-AGATATGTCTCTTCAAGTCT-MGBNFQ | ||
| F. langsethiae | [4,51] | Flan forw | GTTGGCGTCTCACTTATTATT C |
| Flan rev | TGACATTGTTCAGATAGTAGTCC | ||
| Flan probe | 6FAM-CACACC[+C]ATA[+C]CTA[+C]GTGTAA-TAMRA | ||
| F. poae | [52] | TMpoaef | GCTGAGGGTAAGCCGTCCTT |
| TMpoaer | TCTGTCCCCCCTACCAAGCT | ||
| TMpoae probe | TexasRed-ATTTCCCCAACTTC GACTCTCCGAGGA-BHQ2 | ||
| M. majus | [50] | nivale 2-F | CGCCAAGGACTCCTCCAGTAG |
| nivale 2-R | GCCGACGAATGGATATTAAGAACT | ||
| nivale 2 probe | 6FAM-TCCCGCCTTCACGGTGGAAAGC-TAMRA | ||
| M. nivale (SYBR) | [53] | Mniv1f | TTGGCTTGCACAAACAATACTTTTT |
| Mniv1r | AGCACAACAGGCGTGGATAAG | ||
| Cereals | [10] | Cox554f | GGTTGTTGCCACCAAGTCTCTT |
| Cox554r | TGCCGCTGCCAACTTC | ||
| Cox554p | VIC-CTCCTATTAAGCTCAGCCTT-MGBNFQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofgaard, I.S.; Brodal, G.; Almvik, M.; Lillemo, M.; Russenes, A.L.; Edwards, S.G.; Aamot, H.U. Different Resistance to DON versus HT2 + T2 Producers in Nordic Oat Varieties. Toxins 2022, 14, 313. https://doi.org/10.3390/toxins14050313
Hofgaard IS, Brodal G, Almvik M, Lillemo M, Russenes AL, Edwards SG, Aamot HU. Different Resistance to DON versus HT2 + T2 Producers in Nordic Oat Varieties. Toxins. 2022; 14(5):313. https://doi.org/10.3390/toxins14050313
Chicago/Turabian StyleHofgaard, Ingerd Skow, Guro Brodal, Marit Almvik, Morten Lillemo, Aina Lundon Russenes, Simon Graham Edwards, and Heidi Udnes Aamot. 2022. "Different Resistance to DON versus HT2 + T2 Producers in Nordic Oat Varieties" Toxins 14, no. 5: 313. https://doi.org/10.3390/toxins14050313