In Silico Study of the Mechanisms Underlying the Action of the Snake Natriuretic-Like Peptide Lebetin 2 during Cardiac Ischemia

 ,
,  and
and
Abstract
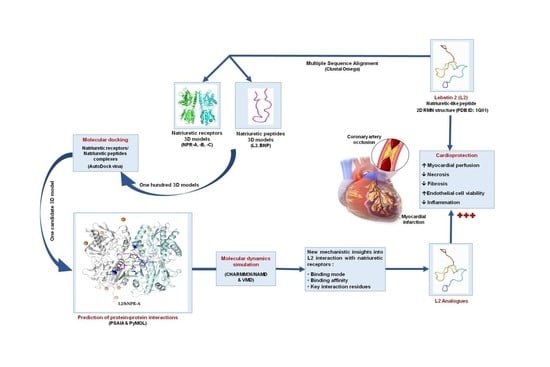
:
1. Introduction
2. Results
2.1. Amino Acid Sequence and Model Analysis
2.2. Molecular Modeling
2.3. Molecular Docking
2.4. Molecular Dynamics Simulations
2.4.1. RMSD Analysis
2.4.2. RMSF Analysis
2.4.3. Radius of Gyration Analysis
2.4.4. Hydrogen Bonds Analysis
2.4.5. Center of Mass Analysis
2.4.6. Binding Free Energy Analysis by the MM/PBSA Method
2.4.7. Receptor–Ligand Interaction Analysis
3. Discussion
4. Conclusions
5. Methods
5.1. Receptor–Ligand Preparation
5.1.1. Multiple Sequence Alignment
5.1.2. Receptor Structure Preparation
5.1.3. Ligand Structure Preparation
5.1.4. Molecular Modeling
5.1.5. Receptor–Ligand Molecular Docking
5.2. Molecular Dynamics Simulation
5.2.1. Analysis of RMSD
5.2.2. Analysis of RMSF
5.2.3. Analysis of Radius of Gyration
5.2.4. Dynamics of Hydrogen Bonds
5.2.5. Analysis of the Center of Mass
5.2.6. Analysis of Binding Free Energy
where
5.2.7. Receptor–Ligand Interaction Analysis
5.2.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Nakanishi, M.; Saito, Y.; Kishimoto, I.; Harada, M.; Kuwahara, K.; Takahashi, N.; Kawakami, R.; Nakagawa, Y.; Tanimoto, K.; Yasuno, S.; et al. Role of natriuretic peptide receptor guanylyl cyclase-A in myocardial infarction evaluated using genetically engineered mice. Hypertension 2005, 46, 441–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello-Boerrigter, L.C.; Boerrigter, G.; Cataliotti, A.; Harty, G.J.; Burnett, J.C., Jr. Renal and anti-aldosterone actions of vasopressin-2 receptor antagonism and B-type natriuretic peptide in experimental heart failure. Circ. Heart Fail. 2010, 3, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukoyama, M.; Nakao, K.; Hosoda, K.; Suga, S.; Saito, Y.; Ogawa, Y.; Shirakami, G.; Jougasaki, M.; Obata, K.; Yasue, H.; et al. Brain natriuretic peptide as a novel cardiac hormone in humans. Evidence for an exquisite dual natriuretic peptide system, atrial natriuretic peptide and brain natriuretic peptide. J. Clin. Investig. 1991, 87, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.G. Natriuretic peptides: Markers or modulators of cardiac hypertrophy? Trends Endocrinol. Metab. 2003, 14, 411–416. [Google Scholar] [CrossRef]
- Yandle, T.G. Biochemistry of natriuretic peptides. J. Intern. Med. 1994, 235, 561–576. [Google Scholar] [CrossRef]
- Burley, D.S.; Hamid, S.A.; Baxter, G.F. Cardioprotective actions of peptide hormones in myocardial ischemia. Heart Fail. Rev. 2007, 12, 279–291. [Google Scholar] [CrossRef]
- Maack, T. Receptors of atrial natriuretic factor. Annu. Rev. Physiol. 1992, 54, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.; Anand-Srivastava, M.B. Cytoplasmic domain of natriuretic peptide receptor C constitutes Gi activator sequences that inhibit adenylyl cyclase activity. J. Biol. Chem. 2001, 276, 22064–22070. [Google Scholar] [CrossRef] [Green Version]
- Koller, K.J.; Goeddel, D.V. Molecular biology of the natriuretic peptides and their receptors. Circulation 1992, 86, 1081–1088. [Google Scholar] [CrossRef]
- Barbouche, R.; Marrakchi, N.; Mansuelle, P.; Krifi, M.; Fenouillet, E.; Rochat, H.; el Ayeb, M. Novel anti-platelet aggregation polypeptides from Vipera lebetina venom: Isolation and characterization. FEBS Lett. 1996, 392, 6–10. [Google Scholar] [CrossRef]
- Vink, S.; Jin, A.H.; Poth, K.J.; Head, G.A.; Alewood, P.F. Natriuretic peptide drug leads from snake venom. Toxicon 2012, 59, 434–445. [Google Scholar] [CrossRef]
- Tourki, B.; Mateo, P.; Morand, J.; Elayeb, M.; Godin-Ribuot, D.; Marrakchi, N.; Belaidi, E.; Messadi, E. Lebetin 2, a Snake Venom-Derived Natriuretic Peptide, Attenuates Acute Myocardial Ischemic Injury through the Modulation of Mitochondrial Permeability Transition Pore at the Time of Reperfusion. PLoS ONE 2016, 11, e0162632–e0162653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tourki, B.; Dumesnil, A.; Belaidi, E.; Ghrir, S.; Godin-Ribuot, D.; Marrakchi, N.; Richard, V.; Mulder, P.; Messadi, E. Lebetin 2, a Snake Venom-Derived B-Type Natriuretic Peptide, Provides Immediate and Prolonged Protection against Myocardial Ischemia-Reperfusion Injury via Modulation of Post-Ischemic Inflammatory Response. Toxins 2019, 11, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefer, A.M.; Tsao, P.S.; Lefer, D.J.; Ma, X.L. Role of endothelial dysfunction in the pathogenesis of reperfusion injury after myocardial ischemia. Faseb J. 1991, 5, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Leitman, D.C.; Andresen, J.W.; Kuno, T.; Kamisaki, Y.; Chang, J.K.; Murad, F. Identification of multiple binding sites for atrial natriuretic factor by affinity cross-linking in cultured endothelial cells. J. Biol. Chem. 1986, 261, 11650–11655. [Google Scholar] [CrossRef]
- Chaudhary, K.R.; Batchu, S.N.; Das, D.; Suresh, M.R.; Falck, J.R.; Graves, J.P.; Zeldin, D.C.; Seubert, J.M. Role of B-type natriuretic peptide in epoxyeicosatrienoic acid-mediated improved post-ischaemic recovery of heart contractile function. Cardiovasc. Res. 2009, 83, 362–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Diaco, M.; Murohara, T.; Ferrara, N.; Isner, J.M.; Symes, J.F. Vascular endothelial growth factor attenuates myocardial ischemia-reperfusion injury. Ann. Thorac. Surg. 1997, 64, 993–998. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Bennett, B.D.; Bennett, G.L.; Vitangcol, R.V.; Jewett, J.R.; Burnier, J.; Henzel, W.; Lowe, D.G. Extracellular domain-IgG fusion proteins for three human natriuretic peptide receptors. Hormone pharmacology and application to solid phase screening of synthetic peptide antisera. J. Biol. Chem. 1991, 266, 23060–23067. [Google Scholar] [CrossRef]
- Suga, S.; Nakao, K.; Hosoda, K.; Mukoyama, M.; Ogawa, Y.; Shirakami, G.; Arai, H.; Saito, Y.; Kambayashi, Y.; Inouye, K.; et al. Receptor selectivity of natriuretic peptide family, atrial natriuretic peptide, brain natriuretic peptide, and C-type natriuretic peptide. Endocrinology 1992, 130, 229–239. [Google Scholar] [CrossRef]
- Johns, D.G.; Ao, Z.; Heidrich, B.J.; Hunsberger, G.E.; Graham, T.; Payne, L.; Elshourbagy, N.; Lu, Q.; Aiyar, N.; Douglas, S.A. Dendroaspis natriuretic peptide binds to the natriuretic peptide clearance receptor. Biochem. Biophys. Res. Commun. 2007, 358, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Shushni, M.A.; Azam, F.; Lindequist, U. Oxasetin from Lophiostoma sp. of the Baltic Sea: Identification, in silico binding mode prediction and antibacterial evaluation against fish pathogenic bacteria. Nat. Prod. Commun. 2013, 8, 1223–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eid, E.E.M.; Alanazi, S.A.; Koosha, S.; Alrasheedy, A.A.; Azam, F.; Taban, I.M.; Khalilullah, H.; Sadiq Al-Qubaisi, M.; Alshawsh, M.A. Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting alphavbeta3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules 2019, 24, 2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosbah, A.; Marrakchi, N.; Mansuelle, P.; Kouidhi, S.; Giralt, E.; Ayeb, M.E.; Herbette, G.; Cherif, A.; Gigmes, D.; Darbon, H.; et al. Lebetin Peptides, A New Class of Potent Platelet Aggregation Inhibitors: Chemical Synthesis, Biological Activity and NMR Spectroscopic Study. Int. J. Pept. Res. Ther. 2020, 26, 21–31. [Google Scholar] [CrossRef]
- Murthy, K.S.; Teng, B.; Jin, J.; Makhlouf, G.M. G protein-dependent activation of smooth muscle eNOS via natriuretic peptide clearance receptor. Am. J. Physiol. 1998, 275, C1409–C1416. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Matsukawa, N.; Grzesik, W.J.; Takahashi, N.; Pandey, K.N.; Pang, S.; Yamauchi, M.; Smithies, O. The natriuretic peptide clearance receptor locally modulates the physiological effects of the natriuretic peptide system. Proc. Natl. Acad. Sci. USA 1999, 96, 7403–7408. [Google Scholar] [CrossRef] [Green Version]
- Rose, R.A.; Giles, W.R. Natriuretic peptide C receptor signalling in the heart and vasculature. J. Physiol. 2008, 586, 353–366. [Google Scholar] [CrossRef]
- Chiba, A.; Watanabe-Takano, H.; Terai, K.; Fukui, H.; Miyazaki, T.; Uemura, M.; Hashimoto, H.; Hibi, M.; Fukuhara, S.; Mochizuki, N. Osteocrin, a peptide secreted from the heart and other tissues, contributes to cranial osteogenesis and chondrogenesis in zebrafish. Development 2017, 144, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Yandle, T.G.; Richards, A.M.; Nicholls, M.G.; Cuneo, R.; Espiner, E.A.; Livesey, J.H. Metabolic clearance rate and plasma half life of alpha-human atrial natriuretic peptide in man. Life Sci. 1986, 38, 1827–1833. [Google Scholar] [CrossRef]
- Mukoyama, M.; Nakao, K.; Saito, Y.; Ogawa, Y.; Hosoda, K.; Suga, S.; Shirakami, G.; Jougasaki, M.; Imura, H. Increased human brain natriuretic peptide in congestive heart failure. N. Engl. J. Med. 1990, 323, 757–758. [Google Scholar] [PubMed]
- He, X.L.; Dukkipati, A.; Garcia, K.C. Structural determinants of natriuretic peptide receptor specificity and degeneracy. J. Mol. Biol. 2006, 361, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Contard, F.; Glukhova, M.; Sabri, A.; Marotte, F.; Sartore, S.; Narcisse, G.; Schatz, C.; Guez, D.; Rappaport, L.; Samuel, J.L. Comparative effects of indapamide and hydrochlorothiazide on cardiac hypertrophy and vascular smooth-muscle phenotype in the stroke-prone, spontaneously hypertensive rat. J. Cardiovasc. Pharmacol. 1993, 22, S29–S34. [Google Scholar] [CrossRef] [PubMed]
- Dickey, D.M.; Flora, D.R.; Bryan, P.M.; Xu, X.; Chen, Y.; Potter, L.R. Differential regulation of membrane guanylyl cyclases in congestive heart failure: Natriuretic peptide receptor (NPR)-B, Not NPR-A, is the predominant natriuretic peptide receptor in the failing heart. Endocrinology 2007, 148, 3518–3522. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, Y.; Itoh, H.; Tamura, N.; Suga, S.; Yoshimasa, T.; Uehira, M.; Matsuda, S.; Shiono, S.; Nishimoto, H.; Nakao, K. Molecular cloning of the complementary DNA and gene that encode mouse brain natriuretic peptide and generation of transgenic mice that overexpress the brain natriuretic peptide gene. J. Clin. Investig. 1994, 93, 1911–1921. [Google Scholar] [CrossRef] [Green Version]
- Van Linthout, S.; Riad, A.; Dhayat, N.; Spillmann, F.; Du, J.; Dhayat, S.; Westermann, D.; Hilfiker-Kleiner, D.; Noutsias, M.; Laufs, U.; et al. Anti-inflammatory effects of atorvastatin improve left ventricular function in experimental diabetic cardiomyopathy. Diabetologia 2007, 50, 1977–1986. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.; Wang, X.; Lau, W.; Wang, Y.; Xing, Y.; Zhang, X.; Ma, X.; Gao, F. Ginsenoside Rd attenuates myocardial ischemia/reperfusion injury via Akt/GSK-3beta signaling and inhibition of the mitochondria-dependent apoptotic pathway. PLoS ONE 2013, 8, e70956–e70965. [Google Scholar] [CrossRef]
- Izumi, T.; Saito, Y.; Kishimoto, I.; Harada, M.; Kuwahara, K.; Hamanaka, I.; Takahashi, N.; Kawakami, R.; Li, Y.; Takemura, G.; et al. Blockade of the natriuretic peptide receptor guanylyl cyclase-A inhibits NF-kappaB activation and alleviates myocardial ischemia/reperfusion injury. J. Clin. Investig. 2001, 108, 203–213. [Google Scholar] [CrossRef]
- Yancy, C.W.; Krum, H.; Massie, B.M.; Silver, M.A.; Stevenson, L.W.; Cheng, M.; Kim, S.S.; Evans, R.; Investigators, F.I. Safety and efficacy of outpatient nesiritide in patients with advanced heart failure: Results of the Second Follow-Up Serial Infusions of Nesiritide (FUSION II) trial. Circ. Heart Fail. 2008, 1, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Rose, R.A. CD-NP, a chimeric natriuretic peptide for the treatment of heart failure. Curr. Opin. Investig. Drugs 2010, 11, 349–356. [Google Scholar]
- Huang, Y.; Ng, X.W.; Lim, S.G.; Chen, H.H.; Burnett, J.C., Jr.; Boey, Y.C.; Venkatraman, S.S. In vivo Evaluation of Cenderitide-Eluting Stent (CES) II. Ann. Biomed. Eng. 2016, 44, 432–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poux, S.; Arighi, C.N.; Magrane, M.; Bateman, A.; Wei, C.H.; Lu, Z.; Boutet, E.; Bye, A.J.H.; Famiglietti, M.L.; Roechert, B.; et al. On expert curation and scalability: UniProtKB/Swiss-Prot as a case study. Bioinformatics 2017, 33, 3454–3460. [Google Scholar] [CrossRef] [PubMed]
- Sussman, J.L.; Lin, D.; Jiang, J.; Manning, N.O.; Prilusky, J.; Ritter, O.; Abola, E.E. Protein Data Bank (PDB): Database of three-dimensional structural information of biological macromolecules. Acta Crystallogr. D Biol. Crystallogr. 1998, 54, 1078–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, H.; Qiu, Y.; Ogata, C.M.; Misono, K.S. Crystal structure of hormone-bound atrial natriuretic peptide receptor extracellular domain: Rotation mechanism for transmembrane signal transduction. J. Biol. Chem. 2004, 279, 28625–28631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Akker, F.; Zhang, X.; Miyagi, M.; Huo, X.; Misono, K.S.; Yee, V.C. Structure of the dimerized hormone-binding domain of a guanylyl-cyclase-coupled receptor. Nature 2000, 406, 101–104. [Google Scholar] [CrossRef]
- He, X.; Chow, D.; Martick, M.M.; Garcia, K.C. Allosteric activation of a spring-loaded natriuretic peptide receptor dimer by hormone. Science 2001, 293, 1657–1662. [Google Scholar] [CrossRef]
- Krauthammer, M.; Rzhetsky, A.; Morozov, P.; Friedman, C. Using BLAST for identifying gene and protein names in journal articles. Gene 2000, 259, 245–252. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Eswar, N.; Eramian, D.; Webb, B.; Shen, M.Y.; Sali, A. Protein structure modeling with MODELLER. Methods Mol. Biol. 2008, 426, 145–159. [Google Scholar]
- Nelson, M.T.; Humphrey, W.; Gursoy, A.; Dalke, A.; Kalé, L.V.; Skeel, R.D.; Schulten, K. Skeel and Klaus Schulten. NAMD: A Parallel, Object-Oriented Molecular Dynamics Program. Int. J. High Perform. Comput. Appl. 1996, 10, 251–268. [Google Scholar]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Chan, H.C.S.; Filipek, S.; Vogel, H. PyMOL and Inkscape Bridge the Data and the Data Visualization. Structure 2016, 24, 2041–2042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.; Li, Y.; Xia, Y.L.; Ai, S.M.; Liang, J.; Sang, P.; Ji, X.L.; Liu, S.Q. Insights into Protein-Ligand Interactions: Mechanisms, Models, and Methods. Int. J. Mol. Sci. 2016, 17, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitencourt-Ferreira, G.; de Azevedo, W.F. Development of a machine-learning model to predict Gibbs free energy of binding for protein-ligand complexes. Biophys. Chem. 2018, 240, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Brooks, C.L., III; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Wong, A.K.; Goscinski, A.M. A VMD plugin for NAMD simulations on Amazon EC2. Procedia Comput. Sci. 2012, 9, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Genheden, S.; Ryde, U.; Soderhjelm, P. Binding affinities by alchemical perturbation using QM/MM with a large QM system and polarizable MM model. J. Comput. Chem. 2015, 36, 2114–2124. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, Y.; Zhao, R.N.; Fan, S.; Han, J.G. Investigation on the mechanism for the binding and drug resistance of wild type and mutations of G86 residue in HIV-1 protease complexed with Darunavir by molecular dynamic simulation and free energy calculation. J. Mol. Model. 2014, 20, 2122–2132. [Google Scholar] [CrossRef]
- Mihel, J.; Sikic, M.; Tomic, S.; Jeren, B.; Vlahovicek, K. PSAIA—Protein structure and interaction analyzer. BMC Struct. Biol. 2008, 8, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand-protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| RMSD (Å) | L2 MODELLER Model |
|---|---|
| AlphaFold_L2 model | 16.576 |
| 1Q01_01 model | 14.110 |
| 1Q01_02 model | 18.277 |
| 1Q01_03 model | 14.878 |
| 1Q01_04 model | 17.960 |
| 1Q01_05 model | 14.591 |
| 1Q01_06 model | 16.407 |
| 1Q01_07 model | 14.373 |
| 1Q01_08 model | 17.637 |
| 1Q01_09 model | 17.412 |
| 1Q01_10 model | 16.558 |
| 1Q01_11 model | 13.952 |
| 1Q01_12 model | 16.115 |
| 1Q01_13 model | 13.961 |
| 1Q01_14 model | 14.061 |
| 1Q01_15 model | 18.362 |
| 1Q01_16 model | 13.592 |
| 1Q01_17 model | 16.661 |
| 1Q01_18 model | 19.069 |
| 1Q01_19 model | 16.237 |
| 1Q01_20 model | 14.835 |
| Human NPRs | Rat NPRs | Mouse NPRs | Chicken NPRs | |||||
|---|---|---|---|---|---|---|---|---|
| NPR-A | L2 | BNP | L2 | BNP | BNP | L2 | L2 | BNP |
| ΔG° (kcal.mol) | −25.4 * | −18.0 | −24.0 * | −16.9 | −22.5 | −19.9 | −19.0 * | −15.4 |
| SEM | 1.0 | 0.4 | 0.8 | 0.2 | 0.2 | 0.8 | 0.2 | 0.2 |
| NPR-B | L2 | BNP | L2 | BNP | L2 | BNP | L2 | BNP |
| ΔG° (kcal.mol) | −20.6 *,† | −19.0 † | −24.0 | −17.0 | −23.9 | −22.4 | −23.1 * | −18.4 |
| SEM | 0.3 | 0.8 | 0.4 | 0.3 | 0.4 | 0.2 | 0.1 | 0.1 |
| NPR-C | L2 | BNP | L2 | BNP | L2 | BNP | L2 | BNP |
| ΔG° (kcal.mol) | −23.4 * | −18.3 £ | −15.5 * | −14.4 | −23.0 * | −19.5 | −20.1 * | −17.7 |
| SEM | 0.7 | 0.4 | 0.2 | 0.1 | 0.2 | 0.1 | 0.2 | 0.2 |
| L2 | BNP | |||||
|---|---|---|---|---|---|---|
| Human | hNPR-A | hNPR-C | hNPR-B | hNPR-B | hNPR-A | hNPR-C |
| ΔG° (kcal.mol) | −24.6 | −20.9 † | −20.1 † | −16.2 | −15.8 | −14.1 |
| SEM | 1.0 | 0.7 | 0.6 | 0.4 | 0.5 | 0.1 |
| Rat | rNPR-A | rNPR-B | rNPR-C | rNPR-B | rNPR-A | rNPR-C |
| ΔG° (kcal.mol) | −23.3 | −17.04 † | −17.0 † | −16.0 | −14.9 £ | −13.6 £ |
| SEM | 1.0 | 0.4 | 0.3 | 0.2 | 0.5 | 0.0 |
| Mouse | mNPR-B | mNPR-C | mNPR-A | mNPR-A | mNPR-B | mNPR-C |
| ΔG° (kcal.mol) | −18.9 | −18.5 | −17.9 | −18.3 | −15.8 | −13.5 † |
| SEM | 0.4 | 0.5 | 0.4 | 1.0 | 0.2 | 0.1 |
| Chicken | chNPR-B | chNPR-A | chNPR-C | chNPR-B | chNPR-A | chNPR-C |
| ΔG° (kcal.mol) | −21.5 | −20.1 | −19.0 £ | −15.0 | −14.7 | −14.4 |
| SEM | 0.4 | 0.4 | 0.4 | 0.2 | 0.5 | 0.4 |
| hNPR-A | hNPR-B | hNPR-C | |
|---|---|---|---|
| L2 | GLY1-GLU119 (A) LYS4-GLU162 (B) LYS7-ASP192 (B) LYS7-ASP191(B) LYS8-GLU187 (B) GLY13-GLU187 (B) PHE15-HSD185 (B) LYS18-ASP177 (A) ARG21-GLU162 (A) ARG21-GLU162 (B) GLY23-GLU162 (B) ASP34-LYS132 (B) ASP34-ARG174 (B) ASP34-ARG178 (B) | GLY1-PHE419 (A) ASP2-ARG143 (A) ASP2-ARG200 (A) LYS7-GLU189 (C) PRO11-GLN177 (A) GLY13-GLU184 (C) LYS18-GLU163 (A) ASP20-ARG183 (C) ARG21-GLU55 (C) GLY23-ASP155 (C) SER24-GLU55 (C) LYS32-GLU55 (A) ASN36-HSD92 (C) LYS37-GLU337 (C) LYS37-ASP338 (C) GLY38-ARG1 (C) | GLY1-GLU180 (A) GLY1-GLU181 (A) LYS4-GLU180 (A) LYS7-ASP197 (B) LYS7-ASP200 (B) LYS18-GLU176 (A) LYS18-GLU180 (A) ASP20-LYS162 (B) ASP20-LYS194 (B) ARG21-GLU172 (A) ARG21-GLU176 (A) LYS37-ASP105 (B) |
| BNP | LYS3-ARG178 (A) ARG13-ASP177 (A) LYS14-GLU169 (A) ARG17-GLU162 (B) SER21-GLU162 (A) SER22-TYR156 (A) SER22-GLU169 (B) GLY25-GLU187 (A) LYS27-GLU187 (A) ARG31-GLU169 (B) | SER1-THR116 (A) SER1-GLN347 (A) ARG13-GLU129 (A) ARG13-GLU393 (A) MET15-GLU167 (A) ARG17-ASP155 (C) SER19-GLU55 (C) SER22-TYR56 (A) SER21-ARG88 (C) ARG30-GLU184 (C) ARG31-ASP155 (C) | ARG13-GLU180 (A) LYS14-HSD120 (A) LYS14-GLU176 (A) LYS14-GLU180 (A) ASP16-ARG99 (A) ASP16-SER123 (A) ARG17-GLU176 (A) SER22-GLY117 (B) LYS27-GLU172 (B) LYS27-GLU176 (B) LEU29-TYR168 (B) HSD32-ARG165 (B) |
| hNPR-A | hNPR-B | hNPR-C | |||||
|---|---|---|---|---|---|---|---|
| Type of Interaction | Monomer 1 | Monomer 2 | Monomer 1 | Monomer 2 | Monomer 1 | Monomer 2 | |
| L2 | Electrostatic | 5 | 5 | 8 | 8 | 7 | 6 |
| Hydrophobic aromatic | 1 | 2 | 1 | 1 | 1 | 2 | |
| Hydrophobic non aromatic | 4 | 5 | 5 | 1 | 3 | 4 | |
| BNP | Electrostatic | 9 | 9 | 18 | 8 | 10 | 3 |
| Hydrophobic aromatic | - | - | - | - | 1 | - | |
| Hydrophobic non aromatic | 12 | 5 | 3 | 6 | 1 | 4 | |
| hNPR-A | hNPR-B | hNPR-C | hNPR-A | hNPR-B | hNPR-C | ||||
|---|---|---|---|---|---|---|---|---|---|
| L2 | Monomer 1 electrostatic interactions | ASP2-ARG95(A) LYS18-ASP177(A) ASP20-ARG95(A) ARG21-TYR120(A) ARG21-GLU169(A) | ASP2-LYS110(A) LYS18-TYR159 (A) LYS18-GLU163(A) LYS18-GLU167(A) ASP20-LYS110(A) ARG21-TYR81(A) ARG21-SER85(A) SER26-TYR56(A) | LYS18-GLU176(A) LYS18-GLU180(A) ARG21-TYR125(A) ARG21-TYR168(A) ARG21-GLU172(A) SER26-LYS162(A) SER26-ARG165(A) | BNP | Monomer 1 electrostatic interactions | SER8-ARG178(A) ARG13-ASP177(A) LYS14-ASP177(A) ARG17-TYR120(A) ARG17-GLU169(A) LYS27-TYR154(A) LYS27-GLU187(A) ARG30-TYR154(A) ARG30-GLU169(A) | ARG13-SER172(A) LYS14-GLU129(A) LYS14-GLU167(A) LYS14-ASP361(A) LYS14-ASP365(A) SER21-SER85(A) SER21-ARG156(A) SER22-ARG156(A) LYS27-TYR148(A) LYS27-ASP150(A) LYS27-TYR159(A) LYS27-TYR181(A) ARG30-TYR148(A) ARG30-GLU163(A) ARG30-GLU167(A) ARG30-TYR148(A) ARG31-TYR148(A) ARG31-TYR159(A) | ARG13-ASP122(A) ARG13-GLU180(A) ASP16-ARG99(A) SER21-ARG165(A) SER22-ARG165(A) LYS27-TYR168(A) ARG30-TYR168(A) ARG30-GLU172(A) ARG30-GLU176(A) ARG31-TYR168 (A) |
| Monomer 1 hydrophobic interactions | PHE15-PHE165(A) PRO11-MET173(A) CYS14-MET173(A) ILE19-VAL116(A) LEU28VAL59(A) | PHE15-PHE166(A) PRO5-LEU174(A) PRO6-LEU174(A) PRO11-VAL176(A) LEU28-PRO82(A) VAL33-CYS53(A) | PHE15-PHE169(A) LEU28-CYS62(A) LEU28-CYS90(A) LEU28-ALA93(A) LEU28-LEU163(A) | Monomer 1 hydrophobic interactions | PRO2-ALA138(A) PRO2-ALA139(A) PRO2-LEU179(A) CYS10-MET173(A) ILE18-VAL87(A) ILE18-ALA90(A) ILE18-ALA91(A) ILE18-ALA111(A) ILE24-ALA155(A) ILE24-PRO158(A) ILE24-ALA189(A) CYS26-ALA189(A) | PRO2-LEU95(A) MET15-ALA168(A) LEU29-VAL180(A) | PHE11-TRP347(A) CYS10-ALA362(A) | ||
| Monomer 2 electrostatic interactions | LYS4-TYR88(B) LYS4-TYR156(B) LYS4-ASP160(B) LYS4-GLU161(B) LYS4-GLU162(B) | LYS4-TYR181(C) LYS7-GLU184(C) LYS8-TYR181(C) LYS8-GLU184(C) ASP20-ARG152(C) ARG21-GLU155(C) ARG21-TYR81(C) ARG21-ASP155(C) | LYS7-ASP195(B) LYS7-TYR188(B) LYS8-ASP191(B) LYS8-ASP197(B) LYS8-ASP200(B) ARG21-168TYR(B) | Monomer 2 electrostatic interactions | ARG17-ASP62(B) ARG17-TUR88(B) LYS27-TYR154(B) LYS27-GLU169(B) LYS27-GLU187(B) ARG31-TYR88(B) ARG31-TYR156(B) ARG31-GLU162(B) ARG31-GLU169(B) | ASP16-ARG152(C) ARG17-TYR81(C) ARG17-ASP150(C) ARG17-THR153(C) ARG17-ASP155(C) ARG31-TYR81(C) ARG31-ASP155(C) ARG31-TYR159(C) | ARG17-TYR92(B) ARG17-GLU164(B) SER21-ARG99(B) | ||
| Monomer 2 hydrophobic interactions | PHE15-TYR154(B) PHE15-PHE165(B) PRO5-PRO158(B) PRO6-PRO158(B) PRO6-ALA189(B) CYS30-VAL116(B) CYS30-MET173(B) | PHE15-TYR181(C) ILE22-LEU149(C) | PHE15-TYR181(B) PHE15-PHE190(B) PRO10-187ILE(B) PRO10-207ALA(B) ILE22-163LEU(B) | Monomer 2 hydrophobic interactions | MET15-VAL59(B) MET15-PRO158(B) LEU24–MET173(B) VAL28-LEU186(B) LEU29-ALA155(B) | ILE18-CYS53(C) ILE18-CYS79(C) ILE18-VAL80(C) ILE18-PRO82(C) ILE18-ALA151(C) LEU29-LEU149(C) | VAL28-ILE187(B) LEU29-LEU156(B) LEU29-LEU171(B) LEU29-ILE187(B) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allaoui, H.; Rached, N.; Marrakchi, N.; Cherif, A.; Mosbah, A.; Messadi, E. In Silico Study of the Mechanisms Underlying the Action of the Snake Natriuretic-Like Peptide Lebetin 2 during Cardiac Ischemia. Toxins 2022, 14, 787. https://doi.org/10.3390/toxins14110787
Allaoui H, Rached N, Marrakchi N, Cherif A, Mosbah A, Messadi E. In Silico Study of the Mechanisms Underlying the Action of the Snake Natriuretic-Like Peptide Lebetin 2 during Cardiac Ischemia. Toxins. 2022; 14(11):787. https://doi.org/10.3390/toxins14110787
Chicago/Turabian StyleAllaoui, Hinda, Nedra Rached, Naziha Marrakchi, Ameur Cherif, Amor Mosbah, and Erij Messadi. 2022. "In Silico Study of the Mechanisms Underlying the Action of the Snake Natriuretic-Like Peptide Lebetin 2 during Cardiac Ischemia" Toxins 14, no. 11: 787. https://doi.org/10.3390/toxins14110787