Effect of Lactic Acid Bacteria on the Fermentation Quality and Mycotoxins Concentrations of Corn Silage Infested with Mycotoxigenic Fungi


Abstract
:1. Introduction
2. Results
2.1. Chemical and Microbial Composition of Fresh Corn
2.2. Fermentation Characteristics of Whole-Crop Corn Silages
2.3. The Concentrations of AFBs, ZEN, DON, and FBs of Whole-Crop Corn Silages
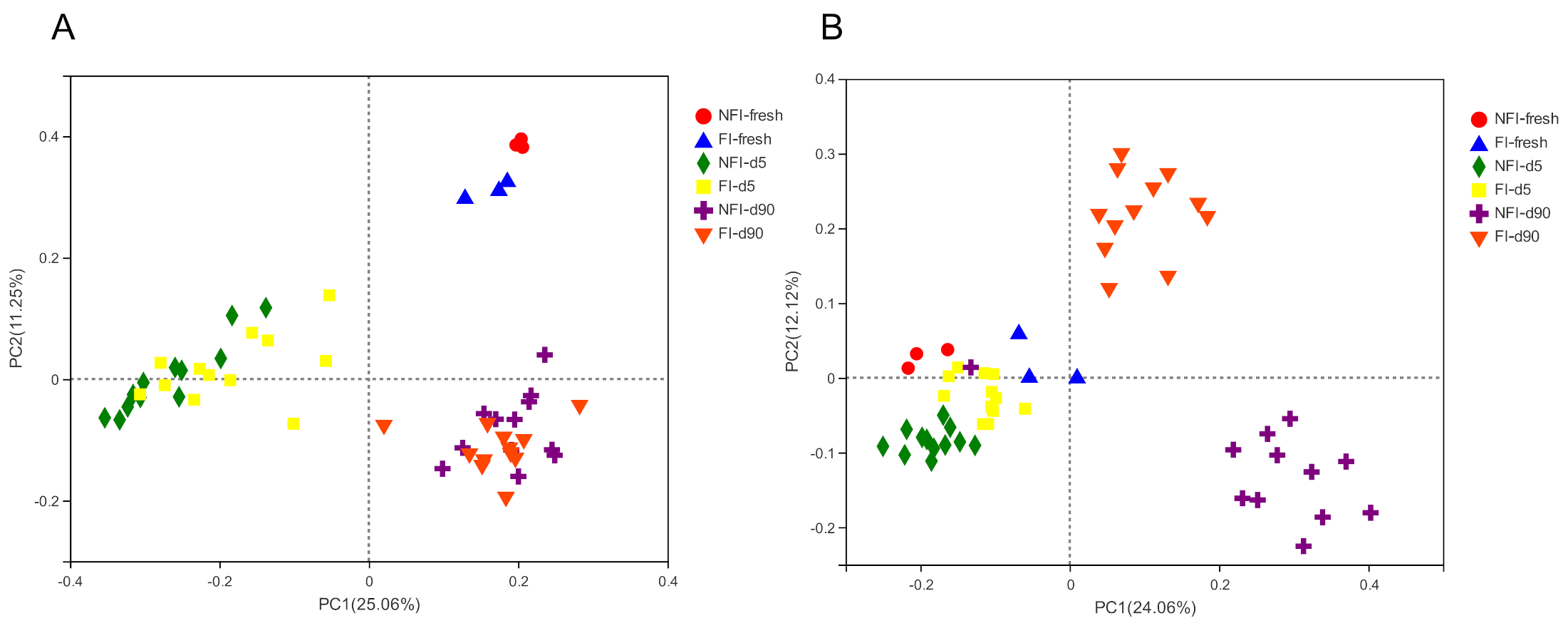
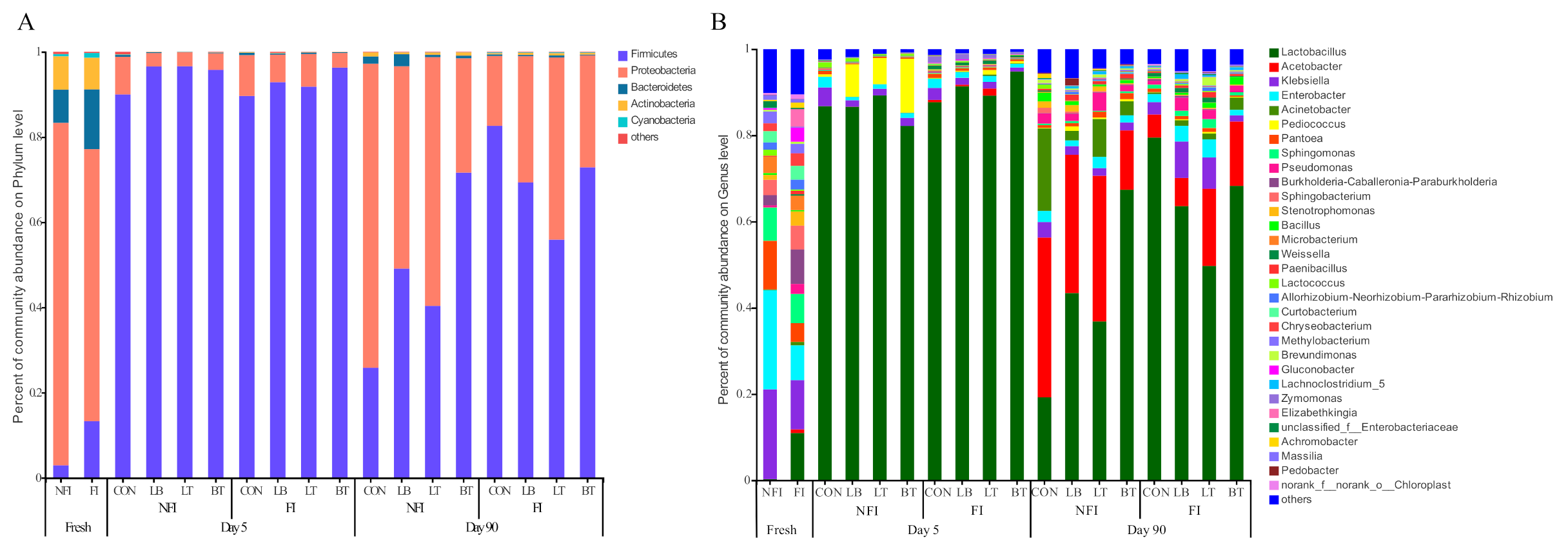
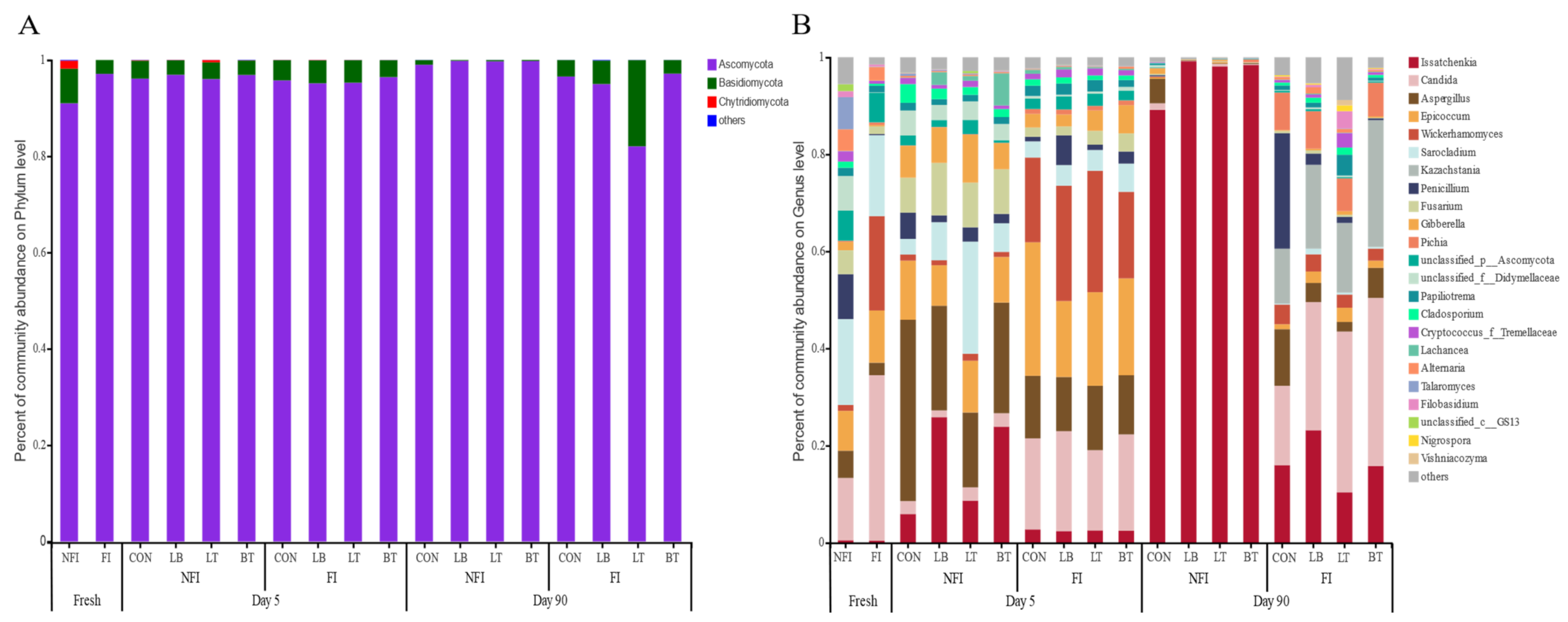
2.4. Bacterial and Fungal Composition of Whole-Crop Corn Silages
3. Discussion
3.1. Effect of Inoculants on Fermentation Quality of Corn with or without Fungal Infestation
3.2. Effect of Inoculants on the AFBS, ZEN, DON, and FBs Concentrations of Corn Silage with or without Fungal Infestation
3.3. Effect of Inoculants on Bacterial and Fungal Diversity and Composition of Corn with or without Fungal Infestation
4. Conclusions
5. Materials and Methods
5.1. Toxigenic Fungi Preparation and Artificial Infestation of Corn in the Field
5.2. Silage Preparation and Treatments
5.3. Sample Preparation and Analysis
5.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Queiroz, O.C.M.; Kim, S.C.; Adesogan, A.T. Effect of Treatment with a Mixture of Bacteria and Fibrolytic Enzymes on the Quality and Safety of Corn Silage Infested with Different Levels of Rust. J. Dairy Sci. 2012, 95, 5285–5291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, T.; De Boevre, M.; De Saeger, S.; Preußke, N.; Sönnichsen, F.D.; Kramer, E.; Klink, H.; Verreet, J.A.; Birr, T. Effect of Ensiling Duration on the Fate of Deoxynivalenol, Zearalenone and Their Derivatives in Maize Silage. Mycotoxin Res. 2020, 36, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.X.; Amaro, F.X.; Romero, J.J.; Pereira, O.G.; Jeong, K.C.; Adesogan, A.T. The Capacity of Silage Inoculant Bacteria to Bind Aflatoxin B1 in Vitro and in Artificially Contaminated Corn Silage. J. Dairy Sci. 2017, 100, 7198–7210. [Google Scholar] [CrossRef] [PubMed]
- Ogunade, I.M.; Jiang, Y.; Cervantes, A.P.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial Diversity and Composition of Alfalfa Silage as Analyzed by Illumina Miseq Sequencing: Effects of Escherichia Coli O157:H7 and Silage Additives. J. Dairy Sci. 2018, 101, 2048–2059. [Google Scholar] [CrossRef]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Kim, D.H.; Jiang, Y.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Control of Escherichia Coli O157:H7 in Contaminated Alfalfa Silage: Effects of Silage Additives. J. Dairy Sci. 2016, 99, 4427–4436. [Google Scholar] [CrossRef]
- Cai, Y.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus Spp. From an Inoculant and of Weissella and Leuconostoc Spp. From Forage Crops on Silage Fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [Green Version]
- Manners, M.; Maclean, D.J.; Scott, K.J. Pathways of Glucose Assimilation in Puccinia Graminis. J. Gen. Virol. 1982, 128, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- Kleinschmit, D.H.; Kung, L., Jr. The Effects of Lactobacillus Buchneri 40788 and Pediococcus Pentosaceus R1094 on the Fermentation of Corn Silage. J. Dairy Sci. 2006, 89, 3999–4004. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Li, T.; Klosterman, S.J.; Tian, C.; Wang, Y. The Bzip Transcription Factor Vdatf1 Regulates Virulence by Mediating Nitrogen Metabolism in Verticillium Dahliae. New Phytol. 2020, 226, 1461–1479. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wen, A.; Desta, S.T.; Dong, Z.; Shao, T. Effects of Four Short-Chain Fatty Acids or Salts on the Dynamics of Nitrogen Transformations and Intrinsic Protease Activity of Alfalfa Silage. J. Sci. Food Agric. 2017, 97, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Liu, S.; Zhao, X.J.; Wang, N.; Jiang, X.; Xin, H.S.; Zhang, Y.G. Lactobacillus Rhamnosus Gg Modulates Gastrointestinal Absorption, Excretion Patterns, and Toxicity in Holstein Calves Fed a Single Dose of Aflatoxin B1. J. Dairy Sci. 2019, 102, 1330–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garon, D.; Richard, E.; Sage, L.; Bouchart, V.; Pottier, D.; Lebailly, P. Mycoflora and Multimycotoxin Detection in Corn Silage: Experimental Study. J. Agric. Food Chem. 2006, 54, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, G.U.; Walker, H.W.; Stahr, H.M. Influence of Temperature, Ph, Water Activity and Antifungal Agents on Growth of Aspergillus Flaws and A. Parasiticus. J. Food Sci. 1983, 48, 778–782. [Google Scholar] [CrossRef]
- Oluwafemi, F.; Kumar, M.; Bandyopadhyay, R.; Ogunbanwo, T.; Ayanwande, K.B. Bio-Detoxification of Aflatoxin B1 in Artificially Contaminated Maize Grains Using Lactic Acid Bacteria. Toxin Rev. 2010, 29, 115–122. [Google Scholar] [CrossRef]
- Ahlberg, S.H.; Joutsjoki, V.; Korhonen, H.J. Potential of Lactic Acid Bacteria in Aflatoxin Risk Mitigation. Int. J. Food Microbiol. 2015, 207, 87–102. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of Fermentative Bacteria for Their Ability to Bind and Biotransform Deoxynivalenol, Zearalenone and Fumonisins in an in Vitro Simulated Corn Silage Model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Vandicke, J.; De Visschere, K.; Ameye, M.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Multi-Mycotoxin Contamination of Maize Silages in Flanders, Belgium: Monitoring Mycotoxin Levels from Seed to Feed. Toxins 2021, 13, 202. [Google Scholar] [CrossRef]
- Vega, M.F.; Dieguez, S.N.; Riccio, B.; Aranguren, S.; Giordano, A.; Denzoin, L.; Soraci, A.L.; Tapia, M.O.; Ross, R.; Apas, A. Zearalenone Adsorption Capacity of Lactic Acid Bacteria Isolated from Pigs. Braz. J. Microbiol. 2017, 48, 715–723. [Google Scholar] [CrossRef]
- Zheng, M.L.; Niu, D.Z.; Jiang, D.; Zuo, S.S.; Xu, C.C. Dynamics of Microbial Community During Ensiling Direct-Cut Alfalfa with and without Lab Inoculant and Sugar. J. Appl. Microbiol. 2017, 122, 1456–1470. [Google Scholar] [CrossRef]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial Diversity and Metabolic Networks in Acid Mine Drainage Habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar]
- Romero, J.J.; Zhao, Y.; Balseca-Paredes, M.A.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Laboratory Silo Type and Inoculation Effects on Nutritional Composition, Fermentation, and Bacterial and Fungal Communities of Oat Silage. J. Dairy Sci. 2017, 100, 1812–1828. [Google Scholar] [CrossRef] [Green Version]
- Fouts, D.E.; Tyler, H.L.; DeBoy, R.T.; Daugherty, S.; Ren, Q.; Badger, J.H.; Durkin, A.S.; Huot, H.; Shrivastava, S.; Kothari, S.; et al. Complete Genome Sequence of the N2-Fixing Broad Host Range Endophyte Klebsiella Pneumoniae 342 and Virulence Predictions Verified in Mice. PLoS Genet. 2008, 4, e1000141. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Dong, Z.; Li, J.; Shao, T. Microbial Community Dynamics and Their Contributions to Organic Acid Production During the Early Stage of the Ensiling of Napier Grass (Pennisetum Purpureum). Grass Forage Sci. 2020, 75, 37–44. [Google Scholar] [CrossRef]
- Guan, H.; Shuai, Y.; Yan, Y.; Ran, Q.; Wang, X.; Li, D.; Cai, Y.; Zhang, X. Microbial Community and Fermentationdynamics of Corn Silage Prepared Withheat-Resistant Lactic Acid Bacteria in a Hotenvironment. Microorganisms 2020, 8, 719. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Jiang, Y.; Kim, D.H.; Cervantes, A.P.; Arriola, K.G.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Fate of Escherichia Coli O157:H7 and Bacterial Diversity in Corn Silage Contaminated with the Pathogen and Treated with Chemical or Microbial Additives. J. Dairy Sci. 2017, 100, 1780–1794. [Google Scholar] [CrossRef]
- Romero, J.J.; Joo, Y.; Park, J.; Tiezzi, F.; Gutierrez-Rodriguez, E.; Castillo, M.S. Bacterial and Fungal Communities, Fermentation, and Aerobic Stability of Conventional Hybrids and Brown Midrib Hybrids Ensiled at Low Moisture with or without a Homo- and Heterofermentative Inoculant. J. Dairy Sci. 2018, 101, 3057–3076. [Google Scholar] [CrossRef] [Green Version]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome Dynamics During Ensiling of Corn with and without Lactobacillus Plantarum Inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Ndagano, D.; Lamoureux, T.; Dortu, C.; Vandermoten, S.; Thonart, P. Antifungal Activity of 2 Lactic Acid Bacteria of the Weissella Genus Isolated from Food. J. Food Sci. 2011, 76, M305–M311. [Google Scholar] [CrossRef]
- Baek, E.; Kim, H.; Choi, H.; Yoon, S.; Kim, J. Antifungal Activity of Leuconostoc Citreum and Weissella Confusa in Rice Cakes. J. Microbiol. 2012, 50, 842–848. [Google Scholar] [CrossRef]
- Wu, B.; Qin, H.; Yang, Y.; Duan, G.; Yang, S.; Xin, F.; Zhao, C.; Shao, H.; Wang, Y.; Zhu, Q. Engineered Zymomonas Mobilis Tolerant to Acetic Acid and Low Ph Via Multiplex Atmospheric and Room Temperature Plasma Mutagenesis. Biotechnol. Biofuels 2019, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hao, W.; Ning, T.; Zheng, M.; Xu, C. Characterization of Culturable Yeast Species Associating with Whole Crop Corn and Total Mixed Ration Silage. Asian-Australas. J. Anim. Sci. 2018, 31, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Ávila, C.L.S.; Carvalho, B.F. Silage Fermentation-Updates Focusing on the Performance of Micro-Organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a Microbial Community During Ensiling and Upon Aerobic Exposure in Lactic Acid Bacteria Inoculation-Treated and Untreated Barley Silages. Bioresour. Technol. 2018, 273, 212–219. [Google Scholar] [CrossRef]
- May, L.A.; Smiley, B.; Schmidt, M.G. Comparative Denaturing Gradient Gel Electrophoresis Analysis of Fungal Communities Associated with Whole Plant Corn Silage. Can. J. Microbiol. 2001, 47, 829–841. [Google Scholar] [CrossRef]
- Ädel Druvefors, U.; Schnürer, J. Mold-Inhibitory Activity of Different Yeast Species During Airtight Storage of Wheat Grain. FEMS Yeast Res. 2005, 5, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, B.F.; Ávila, C.L.S.; Krempser, P.M.; Batista, L.R.; Pereira, M.N.; Schwan, R.F. Occurrence of Mycotoxins and Yeasts and Moulds Identification in Corn Silages in Tropical Climate. J. Appl. Microbiol. 2016, 120, 1181–1192. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Xing, Y.; Pian, R.; Chen, X.; Zhang, Q. Improving the Quality of Rice Straw Silage with Moringa Oleifera Leaves and Propionic Acid: Fermentation, Nutrition, Aerobic Stability and Microbial Communities. Bioresour. Technol. 2020, 299, 122579. [Google Scholar] [CrossRef]
- Cabo, M.L.; Braber, A.F.; Koenraad, P.M.F.J. Apparent Antifungal Activity of Several Lactic Acid Bacteria against Penicillium Discolor Is Due to Acetic Acid in the Medium. J. Food Prot. 2002, 65, 1309–1316. [Google Scholar] [CrossRef]
- Tefera, T.; Vidal, S. Effect of Inoculation Method and Plant Growth Medium on Endophytic Colonization of Sorghum by the Entomopathogenic Fungus Beauveria Bassiana. BioControl 2009, 54, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Arthur Thomas, T. An Automated Procedure for the Determination of Soluble Carbohydrates in Herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and in Vitro Media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items | DM g/kg FW | WSC | CP | NDF | ADF | Starch | LAB | Yeasts and Molds | AB | Enterobacteria |
|---|---|---|---|---|---|---|---|---|---|---|
| FI corn | 339 | 171 | 45.3 | 486 | 243 | 105 | 7.50 | 6.80 a | 8.37 | 6.17 |
| NFI corn | 333 | 177 | 50.7 | 567 | 203 | 112 | 7.20 | 4.37 b | 7.93 | 6.57 |
| Item | pH | Lactic Acid | Acetic Acid | Latic/ Acetic Acid | Ethanol | WSC | Ammonia N g/kg TN | LAB | AB | Yeasts | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| FI | CON | 3.66 a | 88.2 bc | 29.0 ab | 3.04 c | 23.3 | 38.1 | 115 a | 3.87 | 0.00 a | 1.30 |
| LB | 3.64 ab | 95.1 abc | 30.8 a | 3.11 c | 23.5 | 33.0 | 110 ab | 3.25 | 2.43 a | 0.00 | |
| LT | 3.62 b | 88.6 bc | 26.7 b | 3.32 c | 22.5 | 39.1 | 104 ab | 2.87 | 1.67 a | 0.77 | |
| BT | 3.61 b | 98.3 abc | 27.1 b | 3.62 c | 23.4 | 36.4 | 100 b | 3.20 | 0.00 a | 0.33 | |
| NFI | CON | 3.57 c | 87.3 c | 16.0 c | 5.45 b | 24.4 | 41.8 | 71.1 c | 2.98 | 1.60 a | 0.33 |
| LB | 3.53 d | 106 a | 16.1 c | 6.58 a | 20.5 | 32.2 | 71.9 c | 2.93 | 1.10 a | 0.78 | |
| LT | 3.50 e | 105 a | 15.5 c | 6.82 a | 18.2 | 31.2 | 74.3 c | 2.97 | 0.00 a | 0.77 | |
| BT | 3.44 f | 104 ab | 16.0 c | 6.52 a | 18.1 | 30.4 | 66.9 c | 2.53 | 1.67 a | 0.00 | |
| Fungal infestation means | |||||||||||
| FI | 3.63 a | 92.5 b | 28.4 a | 3.27 b | 23.2 a | 36.6 | 107 a | 3.30 | 1.03 | 0.60 | |
| NFI | 3.51 b | 101 a | 15.9 b | 6.34 a | 20.3 b | 33.9 | 71.0 b | 2.85 | 1.09 | 0.47 | |
| Inoculant means | |||||||||||
| CON | 3.61 a | 87.8 b | 22.5 ab | 4.25 b | 23.8 | 40.0 a | 93.3 a | 3.43 | 0.80 | 0.82 | |
| LB | 3.54 c | 100 a | 23.5 a | 4.84 a | 22.0 | 32.6 b | 90.9 ab | 3.09 | 1.77 | 0.39 | |
| LT | 3.58 b | 96.9 ab | 21.1 b | 5.07 a | 20.4 | 35.1 ab | 89.1 ab | 2.92 | 0.83 | 0.77 | |
| BT | 3.56 c | 101 a | 21.5 ab | 5.07 a | 20.8 | 33.4 ab | 83.4 b | 2.87 | 0.83 | 0.17 | |
| SEM | 0.015 | 1.83 | 1.35 | 0.334 | 0.66 | 1.08 | 3.98 | 0.400 | 0.274 | 0.145 | |
| Effects and interactions | |||||||||||
| Fungal infestation | <0.01 | <0.01 | <0.01 | <0.01 | 0.02 | 0.14 | <0.01 | 0.64 | 0.89 | 0.65 | |
| Inoculants | <0.01 | <0.01 | 0.03 | <0.01 | 0.12 | 0.04 | 0.01 | 0.97 | 0.44 | 0.32 | |
| Fungal infestation × inoculants | <0.01 | 0.09 | 0.09 | <0.01 | 0.20 | 0.12 | 0.07 | 0.98 | 0.04 | 0.20 | |
| Item | AFB1 | AFB2 | ZEN | DON | FB1 | FB2 | FB3 | |
|---|---|---|---|---|---|---|---|---|
| FI | Fresh | 35.1 a | 2.77 | 2015 a | 34.8 a | 999 a | 265 | 20.3 |
| CON | 21.3 b | 0.00 | 1598 b | 17.4 b | 376 bc | 259 | 60.0 | |
| LB | 11.2 c | 4.33 | 1518 b | 6.12 c | 173 bc | 91.7 | 47.7 | |
| LT | 13.4 bc | 2.90 | 1563 b | 6.70 c | 126 c | 152 | 68.3 | |
| BT | 7.13 cd | <0.50 | 1094 c | 4.51 c | 207 bc | 148 | 79.3 | |
| NFI | Fresh | <0.50 | <0.50 | 406 d | 21.5 b | 467 b | 129 | 48.7 |
| CON | <0.50 | <0.50 | 449 d | 3.05 c | 266 bc | 60.3 | 18.0 | |
| LB | <0.50 | <0.50 | 405 d | 5.48 c | 366 bc | 67.3 | 20.7 | |
| LT | <0.50 | <0.50 | 488 d | 3.17 c | 246 bc | 79.3 | 27.0 | |
| BT | <0.50 | <0.50 | 335 d | 4.73 c | 210 bc | 72.0 | 17.7 | |
| Infestation means (90 days) | ||||||||
| FI | 13.3 | 1.81 | 1443 a | 8.68 a | 221 b | 163 a | 63.8 a | |
| NFI | <0.50 | <0.50 | 419 b | 4.11 b | 272 a | 69.8 b | 20.8 b | |
| Inoculants means (90 days) | ||||||||
| CON | 10.7 a | 0.00 | 1023 a | 10.2 a | 321 a | 160 | 39.0 | |
| LB | 5.62 b | 2.17 | 962 a | 5.80 b | 270 ab | 79.5 | 34.2 | |
| LT | 6.72 ab | 1.45 | 1026 a | 4.93 b | 186 b | 116 | 47.7 | |
| BT | 3.57 b | 0.00 | 714 b | 4.62 b | 208 b | 110 | 48.5 | |
| SEM | 2.130 | 0.421 | 113.9 | 1.871 | 47.9 | 17.27 | 5.33 | |
| Effects and interactions | ||||||||
| Fungal infestation | <0.01 | 0.06 | <0.01 | <0.01 | 0.03 | <0.01 | <0.01 | |
| Inoculants | <0.01 | 0.27 | <0.01 | <0.01 | <0.01 | 0.27 | 0.71 | |
| Fungal infestation × inoculants | <0.01 | 0.27 | 0.03 | <0.01 | <0.01 | 0.18 | 0.70 | |
| Treatment | Bacterial Community | Fungi Community | ||
|---|---|---|---|---|
| Shannon | Chao 1 | Shannon | Chao 1 | |
| FI Fresh | 3.80 | 427 | 2.33 b | 148 b |
| NFI Fresh | 3.22 | 490 | 3.00 a | 272 a |
| SEM | 0.165 | 19.2 | 0.157 | 29.0 |
| p-value | 0.07 | 0.10 | <0.01 | <0.01 |
| Fungal infestation | ||||
| FI | 2.29 a | 258 | 2.55 a | 168 |
| NFI | 1.58 b | 280 | 1.50 b | 166 |
| Inoculants | ||||
| CON | 1.98 | 307 a | 2.08 | 177 |
| LB | 2.03 | 270 ab | 2.02 | 160 |
| LT | 1.90 | 229 b | 2.1 | 160 |
| BT | 1.82 | 271 ab | 1.89 | 172 |
| Days | ||||
| 5 | 1.52 b | 194 a | 2.68 a | 228 a |
| 90 | 2.35 a | 345 b | 1.36 b | 107 b |
| SEM | 0.095 | 14.7 | 0.154 | 10.5 |
| p-value | ||||
| Fungal infestation | <0.01 | 0.15 | <0.01 | 0.83 |
| Inoculants | 0.35 | <0.01 | 0.32 | 0.50 |
| Days | <0.01 | <0.01 | <0.01 | <0.01 |
| Fungal infestation × inoculants | 0.07 | 0.05 | 0.09 | 0.57 |
| Fungal infestation × days | <0.01 | <0.01 | <0.01 | <0.01 |
| Inoculants × days | 0.2063 | 0.30 | 0.18 | 0.29 |
| Fungal infestation × inoculants × days | 0.6783 | 0.11 | 0.12 | 0.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, W.; Chen, S.; Shao, T.; Tao, X.; Yuan, X. Effect of Lactic Acid Bacteria on the Fermentation Quality and Mycotoxins Concentrations of Corn Silage Infested with Mycotoxigenic Fungi. Toxins 2021, 13, 699. https://doi.org/10.3390/toxins13100699
Li J, Wang W, Chen S, Shao T, Tao X, Yuan X. Effect of Lactic Acid Bacteria on the Fermentation Quality and Mycotoxins Concentrations of Corn Silage Infested with Mycotoxigenic Fungi. Toxins. 2021; 13(10):699. https://doi.org/10.3390/toxins13100699
Chicago/Turabian StyleLi, Jinyang, Wenbo Wang, Sifan Chen, Tao Shao, Xuxiong Tao, and Xianjun Yuan. 2021. "Effect of Lactic Acid Bacteria on the Fermentation Quality and Mycotoxins Concentrations of Corn Silage Infested with Mycotoxigenic Fungi" Toxins 13, no. 10: 699. https://doi.org/10.3390/toxins13100699