
Patterns of Human Milk Oligosaccharides in Mature Milk Are Associated with Certain Gut Microbiota in Infants
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Basic Information Collection
2.3. Sample Collection and Preservation
2.4. HMO Analysis
2.5. 16SRNA Sequencing
2.6. Statistical Analysis
2.7. Ethical Considerations
3. Results
3.1. Characteristics of the Studied Population
3.2. HMO Concentration
3.3. Infant Faecal Microbiota
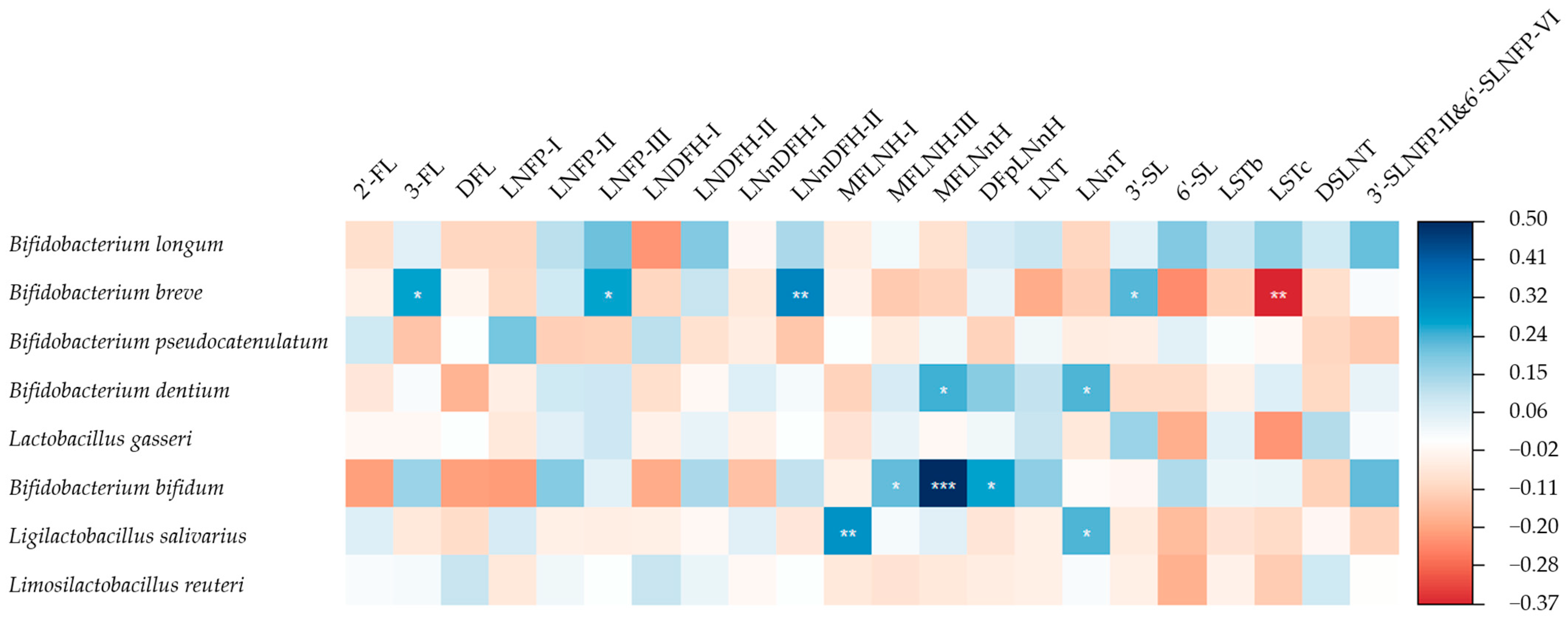
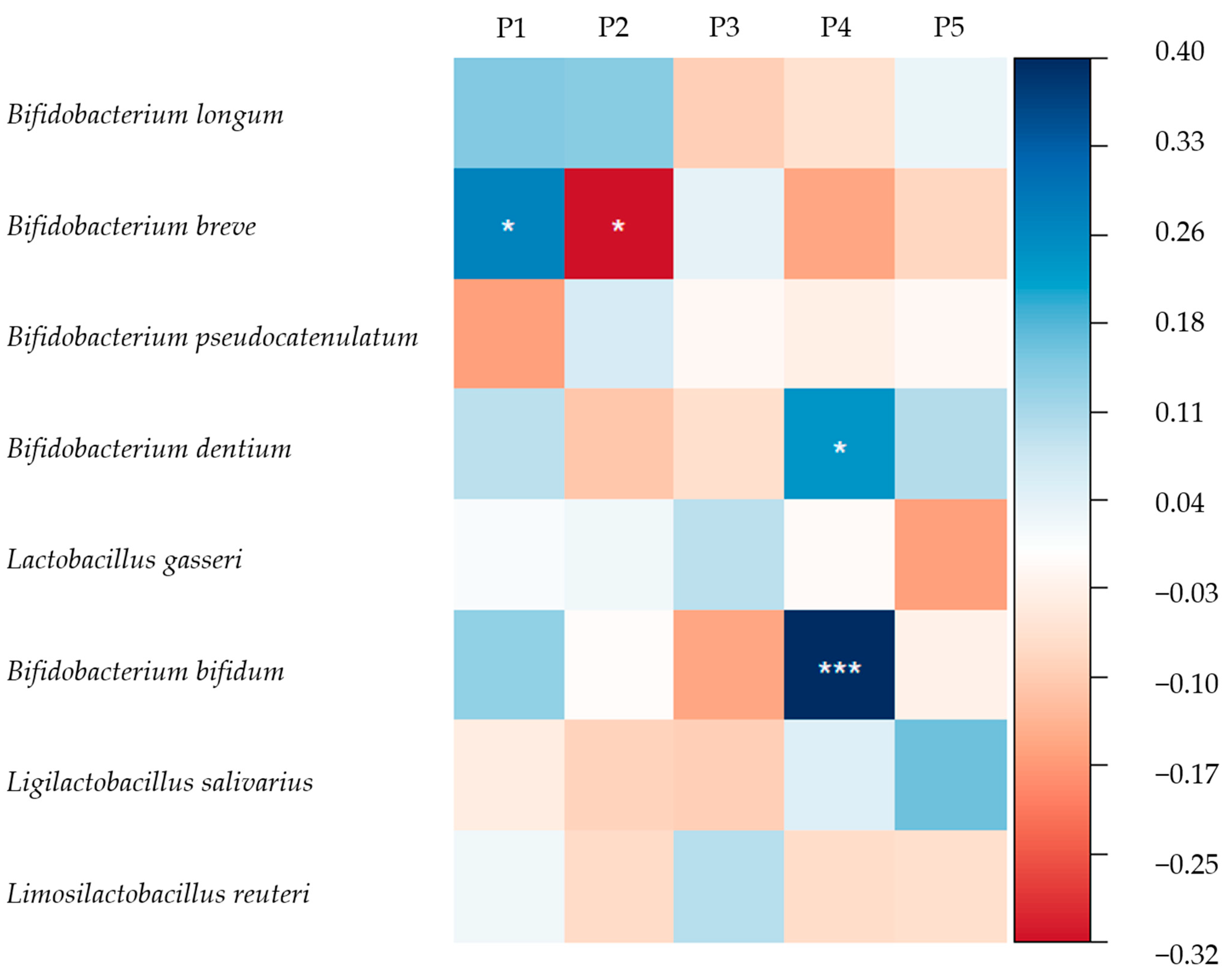
3.4. HMOs and the Gut Microbiome of Breastfed Infants
4. Discussion
4.1. HMOs Concentrations
4.2. Infant Gut Microbiota
4.3. Association of HMO Patterns in Mature Milk with Infant Gut Microbiota
4.3.1. Association of Se/nSe Pattern with Infant Gut Microbiota
4.3.2. Association of Other HMO Patterns with Infant Gut Microbiota
4.4. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HMOs | ≤30 (n = 42) | >30 (n = 54) | p |
|---|---|---|---|
| 2′-FL a | 954.5 (0.0, 1730.6) | 1685.8 (1278.1, 2097.3) | 0.011 a |
| 3-FL a | 911.2 (358.4, 1475.9) | 514.3 (339.9, 740.8) | 0.063 a |
| DFL a | 95.4 (0.0, 214.2) | 211.6 (125.0, 260.2) | 0.002 a |
| LNFP-I a | 75.2 (0.0, 547.0) | 401.7 (168.8, 731.9) | 0.009 a |
| LNFP-II a | 290.8 (103.0, 883.9) | 168.4 (107.1, 345.0) | 0.234 a |
| LNFP-III a | 329.2 (258.5, 410.5) | 289.8 (229.1, 339.9) | 0.049 a |
| LNDFH-I a | 47.3 (0.0, 526.9) | 428.6 (298.0, 608.4) | 0.011 a |
| LNDFH-II a | 40.6 (0.0, 138.6) | 9.2 (0.0, 54.7) | 0.136 a |
| LNnDFH-I a | 14.1 (0.0, 29.3) | 27.1 (17.0, 36.9) | 0.006 a |
| LNnDFH-II a | 41.4 (21.1, 82.7) | 23.6 (16.8, 36.4) | 0.032 a |
| MFLNH-I a | 1.5 (0.0, 59.8) | 35.5 (10.9, 84.5) | 0.024 a |
| MFLNH-III a | 241.5 (153.1, 388.9) | 193.2 (120.1, 258.1) | 0.083 a |
| MFLNnH a | 97.7 (59.0, 183.0) | 96.9 (55.6, 152.9) | 0.679 a |
| DFpLNnH a | 171.2 (113.6, 282.6) | 124.3 (92.7, 183.0) | 0.045 a |
| LNT a | 556.1 (300.1, 899.3) | 456.8 (294.3, 687.8) | 0.305 a |
| LNnT a | 53.6 (32.1, 90.2) | 96.6 (54.6, 136.3) | 0.016 a |
| 3′-SL a | 91.4 (81.8, 106.1) | 91.3 (83.5, 108.9) | 0.621 a |
| 6′-SL a | 250.6 (185.5, 315.6) | 259.1 (156.0, 410.0) | 0.296 a |
| LSTb a | 53.7 (29.6, 77.2) | 50.7 (30.7, 73.5) | 0.805 a |
| LSTc b | 131.1 ± 64.6 | 132.0 ± 57.2 | 0.945 b |
| DSLNT a | 208.5 (150.3, 306.4) | 184.1 (133.0, 286.7) | 0.545 a |
| 3′-SLNFP-II&6′-SLNFP-VI a | 60.1 (29.2, 83.7) | 35.3 (25.3, 60.9) | 0.080 a |
| Sum b | 6025.8 ± 1417.0 | 6174.0 ± 1241.2 | 0.593 b |
| HMOs | Chengdu (n = 15) | Guangzhou (n = 10) | Hohhot (n = 14) | Suzhou (n = 18) | Beijing (n = 39) | p |
|---|---|---|---|---|---|---|
| 2′-FL a | 0.0 (0.0, 1687.1) | 1378.7 (1005.0, 2336.0) | 1445.2 (298.4, 1588.5) | 1529.6 (800.5, 1758.4) | 1882.3 (940.4, 2170.3) | 0.224 a |
| 3-FL a | 612.9 (376.1, 1416.1) | 694.5 (390.0, 885.3) | 867.4 (587.8, 1521.9) | 550.2 (338.8, 1072.2) | 454.7 (302.9, 894.3) | 0.066 a |
| DFL a | 0.0 (0.0, 178.7) | 202.2 (51.2, 242.5) | 217.6 (47.0, 296.9) | 201.3 (104.8, 254.8) | 143.5 (64.2, 244.0) | 0.224 a |
| LNFP-I a | 0.0 (0.0, 441.8) | 196.9 (62.5, 781.8) | 153.7 (7.2, 280.6) | 320.3 (87.7, 688.5) | 558.2 (179.1, 803.9) | 0.025 a |
| LNFP-II a | 418.9 (112.4, 936.9) | 144.1 (105.9, 490.0) | 268.9 (163.3, 476.3) | 165.7 (98.8, 256.6) | 230.2 (98.9, 405.0) | 0.516 a |
| LNFP-III a | 371.0 (330.0, 407.0) | 299.4 (279.3, 330.8) | 319.6 (281.0, 390.1) | 241.6 (193.5, 313.5) | 281.2 (231.2, 374.5) | 0.010 a |
| LNDFH-I a | 39.1 (21.5, 482.3) | 335.4 (69.5, 623.0) | 458.7 (116.1, 599.5) | 312.0 (55.0, 433.4) | 411.8 (0.0, 636.7) | 0.795 a |
| LNDFH-II a | 10.5 (0.0, 172.9) | 0.0 (0.0, 43.1) | 23.2 (3.5, 103.0) | 26.9 (7.8, 50.7) | 0.0 (0.0, 114.2) | 0.715 a |
| LNnDFH-I a | 0.0 (0.0, 26.3) | 17.5 (8.8, 21.1) | 19.2 (3.9, 25.8) | 18.9 (4.6, 27.5) | 32.9 (14.9, 42.4) | 0.065 a |
| LNnDFH-II a | 38.7 (20.2, 78.2) | 25.7 (17.0, 35.0) | 41.4 (25.3, 82.4) | 24.8 (11.0, 43.7) | 33.5 (17.0, 45.4) | 0.175 a |
| MFLNH-I a | 0.0 (0.0, 30.1) | 13.0 (2.7, 36.7) | 21.1 (0.0, 53.2) | 13.9 (0.0, 86.5) | 56.5 (5.4, 119.2) | 0.109 a |
| MFLNH-III a | 195.3 (161.4, 400.4) | 224.9 (147.1, 254.1) | 237.3 (172.5, 307.6) | 133.1 (96.4, 174.3) | 222.3 (174.1, 282.2) | 0.079 a |
| MFLNnH a | 97.6 (74.2, 129.2) | 94.2 (82.8, 119.9) | 154.7 (97.1, 217.8) | 70.8 (53.5, 109.5) | 119.3 (47.2, 179.6) | 0.321 a |
| DFpLNnH a | 152.3 (119.0, 284.5) | 121.0 (84.8, 135.4) | 183.4 (134.1, 220.9) | 98.9 (84.4, 159.7) | 139.9 (98.2, 219.5) | 0.142 a |
| LNT a | 605.9 (430.6, 839.5) | 307.6 (271.2, 393.0) | 399.4 (266.2, 553.5) | 420.8 (296.2, 613.8) | 650.0 (324.6, 933.8) | 0.059 a |
| LNnT a | 52.7 (22.5, 113.5) | 54.5 (30.5, 82.4) | 85.0 (30.3, 102.9) | 82.0 (50.9, 129.3) | 86.0 (48.0, 136.1) | 0.479 a |
| 3′-SL a | 102.2 (85.8, 119.2) | 90.9 (83.3, 104.8) | 84.9 (73.1, 95.3) | 87.4 (82.7, 100.4) | 94.9 (82.3, 112.1) | 0.535 a |
| 6′-SL a | 225.5 (153.8, 279.5) | 240.3 (113.7, 254.4) | 206.9 (133.1, 290.4) | 199.4 (144.1, 288.8) | 361.6 (259.1, 447.1) | <0.001 a |
| LSTb a | 66.6 (42.5, 82.9) | 35.1 (27.9, 48.9) | 47.2 (17.3, 54.5) | 40.0 (24.8, 54.6) | 64.8 (40.3, 83.2) | 0.009 a |
| LSTc b | 147.6 ± 59.9 | 110.5 ± 56.0 | 122.7 ± 70.7 | 105.0 ± 60.0 | 146.3 ± 53.7 | 0.074 b |
| DSLNT a | 235.3 (201.0, 431.9) | 197.0 (140.8, 288.9) | 186.2 (123.9, 281.7) | 126.4 (79.9, 178.4) | 203.2 (166.5, 325.6) | 0.002 a |
| 3′-SLNFP-II&6′-SLNFP-VI a | 59.4 (32.4, 83.1) | 31.2 (22.6, 59.4) | 45.8 (27.1, 60.8) | 32.2 (23.7, 39.8) | 54.4 (33.1, 82.0) | 0.065 a |
| Sum b | 5833.8 ± 1050.7 | 6003.8 ± 1692.3 | 5893.3 ± 1161.2 | 5271.4 ± 1122.4 | 6706.2 ± 1212.8 | 0.002 b |
| HMOs | Caesarean (n = 29) | Vaginal (n = 67) | p |
|---|---|---|---|
| 2′-FL a | 1743.1 (1221.7, 2188.1) | 1485.4 (0.0, 2037.2) | 0.212 a |
| 3-FL a | 533.7 (324.9, 922.9) | 627.5 (363.8, 1133.1) | 0.478 a |
| DFL a | 210.8 (102.8, 253.6) | 143.5 (0.0, 246.7) | 0.348 a |
| LNFP-I a | 364.7 (135.2, 726.3) | 249.8 (0.0, 661.1) | 0.496 a |
| LNFP-II a | 175.3 (104.4, 445.9) | 229.5 (108.3, 728.8) | 0.607 a |
| LNFP-III a | 280.8 (243.5, 350.8) | 311.6 (235.0, 381.9) | 0.555 a |
| LNDFH-I a | 473.4 (303.8, 593.5) | 314.6 (7.6, 536.5) | 0.064 a |
| LNDFH-II a | 6.7 (0.0, 55.6) | 29.4 (0.0, 123.7) | 0.165 a |
| LNnDFH-I a | 27.2 (17.6, 34.9) | 19.1 (0.0, 35.2) | 0.310 a |
| LNnDFH-II a | 30.4 (18.9, 52.6) | 33.5 (16.8, 53.6) | 0.823 a |
| MFLNH-I a | 37.4 (0.0, 105.3) | 16.4 (0.0, 64.4) | 0.084 a |
| MFLNH-III a | 191.2 (168.1, 304.9) | 196.4 (127.9, 301.6) | 0.681 a |
| MFLNnH a | 145.4 (95.2, 182.7) | 89.5 (50.4, 148.1) | 0.013 a |
| DFpLNnH a | 133.2 (99.2, 205.9) | 139.0 (95.3, 225.0) | 0.883 a |
| LNT a | 498.1 (307.3, 644.8) | 456.3 (293.9, 859.1) | 0.563 a |
| LNnT a | 89.5 (59.8, 136.6) | 66.4 (36.8, 120.5) | 0.206 a |
| 3′-SL a | 94.9 (87.5, 119.7) | 88.2 (80.6, 106.5) | 0.070 a |
| 6′-SL a | 312.7 (196.7, 436.9) | 248.4 (155.5, 347.9) | 0.065 a |
| LSTb a | 51.1 (29.5, 67.0) | 51.8 (30.5, 77.1) | 0.753 a |
| LSTc b | 157.1 ± 65.6 | 120.6 ± 54.7 | 0.012 b |
| DSLNT a | 189.5 (157.0, 273.1) | 197.5 (129.3, 312.7) | 0.820 a |
| 3′-SLNFP-II&6′-SLNFP-VI a | 40.9 (33.1, 64.3) | 39.2 (25.1, 74.6) | 0.555 a |
| Sum b | 6258.1 ± 1150.8 | 6044.7 ± 1384.4 | 0.437 b |
| HMOs | Primipara (n = 71) | Multipara (n = 25) | p |
|---|---|---|---|
| 2′-FL a | 1568.0 (0.0, 2127.4) | 1336.0 (1139.2, 1700.1) | 0.495 |
| 3-FL a | 588.8 (360.4, 1168.3) | 518.8 (317.9, 754.4) | 0.341 |
| DFL a | 177.1 (0.0, 257.0) | 167.6 (102.8, 240.5) | 0.960 |
| LNFP-I a | 271.8 (0.0, 662.9) | 404.5 (154.6, 705.8) | 0.441 |
| LNFP-II a | 229.5 (98.8, 728.8) | 167.9 (134.3, 418.9) | 0.809 |
| LNFP-III a | 303.8 (238.2, 374.5) | 300.4 (232.4, 378.9) | 0.764 |
| LNDFH-I a | 338.0 (11.9, 550.5) | 394.6 (288.5, 556.7) | 0.297 |
| LNDFH-II a | 22.0 (0.0, 123.7) | 11.4 (0.0, 55.6) | 0.724 |
| LNnDFH-I a | 20.7 (0.0, 35.1) | 20.2 (6.8, 34.9) | 0.983 |
| LNnDFH-II a | 33.7 (18.4, 56.7) | 28.8 (15.7, 36.7) | 0.236 |
| MFLNH-I a | 25.6 (0.0, 72.9) | 24.5 (2.2, 84.8) | 0.426 |
| MFLNH-III a | 196.4 (132.9, 298.0) | 191.2 (157.9, 313.7) | 0.726 |
| MFLNnH a | 96.5 (54.9, 173.3) | 111.1 (59.9, 155.9) | 0.943 |
| DFpLNnH a | 139.9 (99.5, 225.6) | 129.6 (86.9, 142.9) | 0.299 |
| LNT a | 445.3 (280.1, 700.7) | 675.5 (413.3, 804.4) | 0.130 |
| LNnT a | 69.7 (39.2, 113.6) | 107.1 (47.5, 142.6) | 0.151 |
| 3′-SL a | 94.3 (82.7, 112.1) | 85.9 (79.6, 101.2) | 0.092 |
| 6′-SL a | 266.6 (175.6, 377.6) | 248.8 (155.5, 369.4) | 0.475 |
| LSTb a | 51.1 (28.2, 71.0) | 54.9 (39.1, 83.2) | 0.172 |
| LSTc b | 131.9 ± 58.1 | 130.6 ± 67.4 | 0.932 |
| DSLNT a | 197.5 (145.9, 306.0) | 184.1 (132.3, 266.6) | 0.673 |
| 3′-SLNFP-II&6′-SLNFP-VI a | 40.9 (28.5, 77.0) | 36.4 (23.9, 61.4) | 0.455 |
| Sum b | 6163.2 ± 1267.9 | 5955.8 ± 1460.3 | 0.532 |
References
- WHO. Breastfeeding-Recommendations; WHO: Geneva, Switzerland, 2022.
- Christian, P.; Smith, E.R.; Lee, S.E.; Vargas, A.J.; Bremer, A.A.; Raiten, D.J. The need to study human milk as a biological system. Am. J. Clin. Nutr. 2021, 113, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Lonnerdal, B. Human Milk MicroRNAs/Exosomes: Composition and Biological Effects. In Human Milk: Composition, Clinical Benefits and Future Opportunities; Nestlé Nutrition Institute Workshop Series; Karger Publishers: Basel, Switzerland, 2019; Volume 90, pp. 83–92. [Google Scholar] [CrossRef]
- Kim, S.Y.; Yi, D.Y. Components of human breast milk: From macronutrient to microbiome and microRNA. Clin. Exp. Pediatr. 2020, 63, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Vizzari, G.; Morniroli, D.; Ceroni, F.; Verduci, E.; Consales, A.; Colombo, L.; Cerasani, J.; Mosca, F.; Gianni, M.L. Human Milk, More Than Simple Nourishment. Children 2021, 8, 863. [Google Scholar] [CrossRef] [PubMed]
- Caba-Flores, M.D.; Ramos-Ligonio, A.; Camacho-Morales, A.; Martinez-Valenzuela, C.; Viveros-Contreras, R.; Caba, M. Breast Milk and the Importance of Chrononutrition. Front. Nutr. 2022, 9, 867507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Deng, Q.; Wang, J.; Wang, H.; Li, Q.; Zhu, B.; Ji, C.; Xu, X.; Johnston, L. The impact of breast milk feeding on early brain development in preterm infants in China: An observational study. PLoS ONE 2022, 17, e272125. [Google Scholar] [CrossRef]
- Binia, A.; Lavalle, L.; Chen, C.; Austin, S.; Agosti, M.; Al-Jashi, I.; Pereira, A.B.; Costeira, M.J.; Silva, M.G.; Marchini, G.; et al. Human milk oligosaccharides, infant growth, and adiposity over the first 4 months of lactation. Pediatr. Res. 2021, 90, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.D.; Chaturvedi, P.; Newburg, A.R.; Oftedal, O.T.; Tilden, C.D.; Newburg, D.S. Comparison of oligosaccharides in milk specimens from humans and twelve other species. Adv. Exp. Med. Biol. 2001, 501, 325–332. [Google Scholar] [CrossRef]
- Cheng, L.; Akkerman, R.; Kong, C.; Walvoort, M.; de Vos, P. More than sugar in the milk: Human milk oligosaccharides as essential bioactive molecules in breast milk and current insight in beneficial effects. Crit. Rev. Food Sci. Nutr. 2021, 61, 1184–1200. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef]
- Wicinski, M.; Sawicka, E.; Gebalski, J.; Kubiak, K.; Malinowski, B. Human Milk Oligosaccharides: Health Benefits, Potential Applications in Infant Formulas, and Pharmacology. Nutrients 2020, 12, 266. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; Garcia-Mantrana, I.; Bertua-Rios, B.; Martinez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Williams, J.E.; McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Brooker, S.L.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; Prentice, A.M.; et al. Key genetic variants associated with variation of milk oligosaccharides from diverse human populations. Genomics 2021, 113, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Tao, J.; Zhou, J.; Fan, Q.; Liu, M.; Hu, Y.; Xu, Y.; Zhang, L.; Yuan, J.; Li, W.; et al. Fucosylated Human Milk Oligosaccharides and N-Glycans in the Milk of Chinese Mothers Regulate the Gut Microbiome of Their Breast-Fed Infants during Different Lactation Stages. mSystems 2018, 3, e00206-18. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, Z.; Zhao, A.; Zhang, J.; Wu, W.; Ren, Z.; Wang, P.; Zhang, Y. Neutral Human Milk Oligosaccharides Are Associated with Multiple Fixed and Modifiable Maternal and Infant Characteristics. Nutrients 2020, 12, 826. [Google Scholar] [CrossRef]
- Wu, J.; Wu, S.; Huo, J.; Ruan, H.; Xu, X.; Hao, Z.; Wei, Y. Systematic Characterization and Longitudinal Study Reveal Distinguishing Features of Human Milk Oligosaccharides in China. Curr. Dev. Nutr. 2020, 4, a113. [Google Scholar] [CrossRef] [PubMed]
- Castanys-Mu Oz, E.; Martin, M.J.; Prieto, P.A. 2′-fucosyllactose: An abundant, genetically determined soluble glycan present in human milk. Nutr. Rev. 2013, 71, 773–789. [Google Scholar] [CrossRef]
- Zhang, S.; Li, T.; Xie, J.; Zhang, D.; Pi, C.; Zhou, L.; Yang, W. Gold standard for nutrition: A review of human milk oligosaccharide and its effects on infant gut microbiota. Microb. Cell Factories 2021, 20, 108. [Google Scholar] [CrossRef] [PubMed]
- Soyyilmaz, B.; Miks, M.H.; Rohrig, C.H.; Matwiejuk, M.; Meszaros-Matwiejuk, A.; Vigsnaes, L.K. The Mean of Milk: A Review of Human Milk Oligosaccharide Concentrations throughout Lactation. Nutrients 2021, 13, 2737. [Google Scholar] [CrossRef]
- Huang, C.; Lu, Y.; Kong, L.; Guo, Z.; Zhao, K.; Xiang, Z.; Ma, X.; Gao, H.; Liu, Y.; Gao, Z.; et al. Human milk oligosaccharides in milk of mothers with term and preterm delivery at different lactation stage. Carbohydr. Polym. 2023, 321, 121263. [Google Scholar] [CrossRef]
- McGuire, M.K.; Meehan, C.L.; McGuire, M.A.; Williams, J.E.; Foster, J.; Sellen, D.W.; Kamau-Mbuthia, E.W.; Kamundia, E.W.; Mbugua, S.; Moore, S.E.; et al. What’s normal? Oligosaccharide concentrations and profiles in milk produced by healthy women vary geographically. Am. J. Clin. Nutr. 2017, 105, 1086–1100. [Google Scholar] [CrossRef]
- Dinleyici, M.; Barbieur, J.; Dinleyici, E.C.; Vandenplas, Y. Functional effects of human milk oligosaccharides (HMOs). Gut Microbes 2023, 15, 2186115. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Shang, J.; Yang, S.; Meng, X. Correlations between oligosaccharides in breast milk and the composition of the gut microbiome in breastfed infants. J. Dairy Sci. 2022, 105, 4818–4828. [Google Scholar] [CrossRef] [PubMed]
- Siziba, L.P.; Mank, M.; Stahl, B.; Gonsalves, J.; Blijenberg, B.; Rothenbacher, D.; Genuneit, J. Human Milk Oligosaccharide Profiles over 12 Months of Lactation: The Ulm SPATZ Health Study. Nutrients 2021, 13, 1973. [Google Scholar] [CrossRef] [PubMed]
- Turpin, W.; Bedrani, L.; Espin-Garcia, O.; Xu, W.; Silverberg, M.S.; Smith, M.I.; Guttman, D.S.; Griffiths, A.; Moayyedi, P.; Panaccione, R.; et al. FUT2 genotype and secretory status are not associated with fecal microbial composition and inferred function in healthy subjects. Gut Microbes 2018, 9, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Ramani, S.; Stewart, C.J.; Laucirica, D.R.; Ajami, N.J.; Robertson, B.; Autran, C.A.; Shinge, D.; Rani, S.; Anandan, S.; Hu, L.; et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nat. Commun. 2018, 9, 5010. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Bi, Y.; Zheng, Y.; Cao, C.; Yu, L.; Yang, Z.; Chai, W.; Yan, J.; Lai, J.; Liang, X. Development of high-throughput UPLC-MS/MS using multiple reaction monitoring for quantitation of complex human milk oligosaccharides and application to large population survey of secretor status and Lewis blood group. Food Chem. 2022, 397, 133750. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, M.; Yang, X.; Sun, J.; Weng, L.; Qiu, Z. Rice Water-Fried Atractylodis Rhizoma Relieves Spleen Deficiency Diarrhea by Regulating the Intestinal Microbiome. Oxidative Med. Cell. Longev. 2023, 2023, 1983616. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Tonon, K.M.; de Morais, B.M.; Abrão, A.C.F.V.; Miranda, A.; Morais, T.B. Maternal and Infant Factors Associated with Human Milk Oligosaccharides Concentrations According to Secretor and Lewis Phenotypes. Nutrients 2019, 11, 1358. [Google Scholar] [CrossRef]
- Ferreira, A.L.; Alves, R.; Figueiredo, A.; Alves-Santos, N.; Freitas-Costa, N.; Batalha, M.; Yonemitsu, C.; Manivong, N.; Furst, A.; Bode, L.; et al. Human Milk Oligosaccharide Profile Variation Throughout Postpartum in Healthy Women in a Brazilian Cohort. Nutrients 2020, 12, 790. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, M.S.; Wang, R.S.; Hu, J.Q.; Liu, S.; Wang, Y.Y.; Xing, X.L.; Zhang, B.W.; Liu, J.M.; Wang, S. Current Advances in Structure-Function Relationships and Dose-Dependent Effects of Human Milk Oligosaccharides. J. Agric. Food Chem. 2022, 70, 6328–6353. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Davis, J.C.; Goonatilleke, E.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Absolute Quantitation of Human Milk Oligosaccharides Reveals Phenotypic Variations during Lactation. J. Nutr. 2017, 147, 117–124. [Google Scholar] [CrossRef]
- Viverge, D.; Grimmonprez, L.; Cassanas, G.; Bardet, L.; Bonnet, H.; Solere, M. Variations of lactose and oligosaccharides in milk from women of blood types secretor A or H, secretor Lewis, and secretor H/nonsecretor Lewis during the course of lactation. Ann. Nutr. Metab. 1985, 29, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.; Gabrielli, O.; Pierani, P.; Catassi, C.; Carlucci, A.; Giorgi, P.L. Changes in carbohydrate composition in human milk over 4 months of lactation. Pediatrics 1993, 91, 637–641. [Google Scholar] [CrossRef] [PubMed]
- Elwakiel, M.; Hageman, J.A.; Wang, W.; Szeto, I.M.; van Goudoever, J.B.; Hettinga, K.A.; Schols, H.A. Human Milk Oligosaccharides in Colostrum and Mature Milk of Chinese Mothers: Lewis Positive Secretor Subgroups. J. Agric. Food Chem. 2018, 66, 7036–7043. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Mao, Y.; Liu, S.; Wang, J.; Li, X.; Zhao, Y.; Hill, D.R.; Wang, S. Vitamins, Vegetables and Metal Elements Are Positively Associated with Breast Milk Oligosaccharide Composition among Mothers in Tianjin, China. Nutrients 2022, 14, 4131. [Google Scholar] [CrossRef]
- Zhang, W.; Vervoort, J.; Pan, J.; Gao, P.; Zhu, H.; Wang, X.; Zhang, Y.; Chen, B.; Liu, Y.; Li, Y.; et al. Comparison of twelve human milk oligosaccharides in mature milk from different areas in China in the Chinese Human Milk Project (CHMP) study. Food Chem. 2022, 395, 133554. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, T.; Chen, X.; Pang, X.; Zhang, S.; Obaroakpo, J.U.; Shilong, J.; Lu, J.; Lv, J. Absolute quantification of twelve oligosaccharides in human milk using a targeted mass spectrometry-based approach. Carbohydr. Polym. 2019, 219, 328–333. [Google Scholar] [CrossRef]
- Liu, S.; Mao, Y.; Wang, J.; Tian, F.; Hill, D.R.; Xiong, X.; Li, X.; Zhao, Y.; Wang, S. Lactational and geographical variation in the concentration of six oligosaccharides in Chinese breast milk: A multicenter study over 13 months postpartum. Front. Nutr. 2023, 10, 1267287. [Google Scholar] [CrossRef]
- Jiang, S.; Pan, J.; Li, Y.; Ju, M.; Zhang, W.; Lu, J.; Lv, J.; Li, K. Comprehensive Human Milk Patterns Are Related to Infant Growth and Allergy in the CHMP Study. Mol. Nutr. Food Res. 2021, 65, e2100011. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Zhao, F. Microbial transmission, colonisation and succession: From pregnancy to infancy. Gut 2023, 72, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Kiely, L.J.; Busca, K.; Lane, J.A.; van Sinderen, D.; Hickey, R.M. Molecular strategies for the utilisation of human milk oligosaccharides by infant gut-associated bacteria. FEMS Microbiol. Rev. 2023, 47, fuad056. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Diana, A.; Rahmannia, S.; Gibson, R.S.; Houghton, L.A.; Slupsky, C.M. Impact of milk secretor status on the fecal metabolome and microbiota of breastfed infants. Gut Microbes 2023, 15, 2257273. [Google Scholar] [CrossRef] [PubMed]
- Katoh, T.; Ojima, M.N.; Sakanaka, M.; Ashida, H.; Gotoh, A.; Katayama, T. Enzymatic Adaptation of Bifidobacterium bifidum to Host Glycans, Viewed from Glycoside Hydrolyases and Carbohydrate-Binding Modules. Microorganisms 2020, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, M.; Li, C.; Miao, M.; Zhang, T. Effects of Human Milk Oligosaccharides in Infant Health Based on Gut Microbiota Alteration. J. Agric. Food Chem. 2023, 71, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Thorman, A.W.; Adkins, G.; Conrey, S.C.; Burrell, A.R.; Yu, Y.; White, B.; Burke, R.; Haslam, D.; Payne, D.C.; Staat, M.A.; et al. Gut Microbiome Composition and Metabolic Capacity Differ by FUT2 Secretor Status in Exclusively Breastfed Infants. Nutrients 2023, 15, 471. [Google Scholar] [CrossRef] [PubMed]
- Lewis, Z.T.; Totten, S.M.; Smilowitz, J.T.; Popovic, M.; Parker, E.; Lemay, D.G.; Van Tassell, M.L.; Miller, M.J.; Jin, Y.S.; German, J.B.; et al. Maternal fucosyltransferase 2 status affects the gut bifidobacterial communities of breastfed infants. Microbiome 2015, 3, 13. [Google Scholar] [CrossRef]
- Bunesova, V.; Lacroix, C.; Schwab, C. Fucosyllactose and L-fucose utilization of infant Bifidobacterium longum and Bifidobacterium kashiwanohense. BMC Microbiol. 2016, 16, 248. [Google Scholar] [CrossRef]
- Zheng, J.; Xu, H.; Fang, J.; Zhang, X. Enzymatic and chemoenzymatic synthesis of human milk oligosaccharides and derivatives. Carbohydr. Polym. 2022, 291, 119564. [Google Scholar] [CrossRef]
- Aakko, J.; Kumar, H.; Rautava, S.; Wise, A.; Autran, C.; Bode, L.; Isolauri, E.; Salminen, S. Human milk oligosaccharide categories define the microbiota composition in human colostrum. Benef. Microbes 2017, 8, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Moyano, S.; Totten, S.M.; Garrido, D.A.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Variation in consumption of human milk oligosaccharides by infant gut-associated strains of Bifidobacterium breve. Appl. Environ. Microbiol. 2013, 79, 6040–6049. [Google Scholar] [CrossRef] [PubMed]
- Lordan, C.; Roche, A.K.; Delsing, D.; Nauta, A.; Groeneveld, A.; MacSharry, J.; Cotter, P.D.; van Sinderen, D. Linking human milk oligosaccharide metabolism and early life gut microbiota: Bifidobacteria and beyond. Microbiol. Mol. Biol. Rev. 2024, 88, e9423. [Google Scholar] [CrossRef] [PubMed]
- Ojima, M.N.; Jiang, L.; Arzamasov, A.A.; Yoshida, K.; Odamaki, T.; Xiao, J.; Nakajima, A.; Kitaoka, M.; Hirose, J.; Urashima, T.; et al. Priority effects shape the structure of infant-type Bifidobacterium communities on human milk oligosaccharides. ISME J. 2022, 16, 2265–2279. [Google Scholar] [CrossRef]
- Borewicz, K.; Gu, F.; Saccenti, E.; Arts, I.; Penders, J.; Thijs, C.; van Leeuwen, S.S.; Lindner, C.; Nauta, A.; van Leusen, E.; et al. Correlating Infant Fecal Microbiota Composition and Human Milk Oligosaccharide Consumption by Microbiota of 1-Month-Old Breastfed Infants. Mol. Nutr. Food Res. 2019, 63, e1801214. [Google Scholar] [CrossRef] [PubMed]
- Moya-Gonzalvez, E.M.; Rubio-Del-Campo, A.; Rodriguez-Diaz, J.; Yebra, M.J. Infant-gut associated Bifidobacterium dentium strains utilize the galactose moiety and release lacto-N-triose from the human milk oligosaccharides lacto-N-tetraose and lacto-N-neotetraose. Sci. Rep. 2021, 11, 23328. [Google Scholar] [CrossRef] [PubMed]
- Sakanaka, M.; Gotoh, A.; Yoshida, K.; Odamaki, T.; Koguchi, H.; Xiao, J.Z.; Kitaoka, M.; Katayama, T. Varied Pathways of Infant Gut-Associated Bifidobacterium to Assimilate Human Milk Oligosaccharides: Prevalence of the Gene Set and Its Correlation with Bifidobacteria-Rich Microbiota Formation. Nutrients 2019, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T. Host-derived glycans serve as selected nutrients for the gut microbe: Human milk oligosaccharides and bifidobacteria. Biosci. Biotechnol. Biochem. 2016, 80, 621–632. [Google Scholar] [CrossRef]
- Kassai, S.; de Vos, P. Gastrointestinal barrier function, immunity, and neurocognition: The role of human milk oligosaccharide (hMO) supplementation in infant formula. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13271. [Google Scholar] [CrossRef]





| HMOs | Se (n = 70) | nSe (n = 26) | p |
|---|---|---|---|
| 2′-FL | 1760.5 (1405.0, 2188.3) | - | - |
| 3-FL a | 454.0 (317.3, 638.5) | 1522.7 (1181.0, 1871.8) | <0.001 |
| DFL | 216.4 (144.5, 271.7) | - | - |
| LNFP-I | 447.3 (252.1, 804.5) | - | - |
| LNFP-II a | 154.8 (94.0, 250.3) | 992.7 (834.9, 1191.8) | <0.001 |
| LNFP-III a | 281.0 (226.2, 335.2) | 403.5 (320.1, 464.8) | <0.001 |
| LNDFH-I a | 479.9 (318.5, 639.9) | 3.9 (0.0, 21.1) | <0.001 |
| LNDFH-II a | 0.0 (0.0, 35.2) | 179.7 (119.1, 328.2) | <0.001 |
| LNnDFH-I | 28.2 (19.1, 37.1) | - | - |
| LNnDFH-II a | 22.4 (15.7, 34.0) | 90.8 (59.1, 116.7) | <0.001 |
| MFLNH-I | 48.0 (17.9, 98.4) | - | - |
| MFLNH-III a | 178.0 (120.6, 243.0) | 423.7 (225.4, 626.9) | <0.001 |
| MFLNnH a | 97.8 (57.1, 154.4) | 96.4 (50.1, 215.7) | 0.677 |
| DFpLNnH a | 116.4 (84.1, 140.3) | 361.2 (278.9, 465.4) | <0.001 |
| LNT a | 374.9 (279.7, 578.8) | 896.2 (652.5, 1081.7) | <0.001 |
| LNnT a | 104.9 (65.0, 136.3) | 36.4 (18.2, 51.7) | <0.001 |
| 3′-SL a | 88.7 (81.1, 106.1) | 101.8 (86.5, 112.9) | 0.124 |
| 6′-SL a | 249.6 (158.4, 348.9) | 284.2 (226.7, 391.9) | 0.213 |
| LSTb a | 41.4 (26.3, 57.9) | 77.1 (62.0, 102.6) | <0.001 |
| LSTc b | 133.9 ± 63.6 | 125.4 ± 50.9 | 0.503 |
| DSLNT a | 172.8 (121.0, 257.6) | 281.4 (217.7, 416.4) | <0.001 |
| 3′-SLNFP-II&6′-SLNFP-VI a | 33.8 (24.3, 54.3) | 91.3 (75.9, 119.4) | <0.001 |
| Sum b | 6063.2 ± 1299.9 | 6232.8 ± 1376.3 | 0.589 |
| Species | Crude Model | Model 1 |
|---|---|---|
| Bifidobacterium longum | −0.088 (−0.292, 0.116) | −0.161 (−0.385, 0.063) |
| Bifidobacterium breve | −0.129 (−0.332, 0.074) | −0.111 (−0.334, 0.113) |
| Bifidobacterium pseudocatenulatum | 0.132 (−0.071, 0.335) | 0.165 (−0.058, 0.388) |
| Bifidobacterium dentium | −0.056 (−0.261, 0.148) | −0.128 (−0.353, 0.097) |
| Lactobacillus gasseri | 0.039 (−0.166, 0.244) | 0.013 (−0.207, 0.233) |
| Bifidobacterium bifidum | −0.160 (−0.362, 0.042) | −0.245 (−0.465, −0.025) |
| Ligilactobacillus salivarius | 0.065 (−0.140, 0.269) | 0.032 (−0.196, 0.259) |
| Limosilactobacillus reuteri | 0.087 (−0.117, 0.291) | 0.063 (−0.154, 0.281) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, S.; Zhao, A.; Jiang, H.; Yan, J.; Zhong, W.; Xun, Y.; Zhang, Y. Patterns of Human Milk Oligosaccharides in Mature Milk Are Associated with Certain Gut Microbiota in Infants. Nutrients 2024, 16, 1287. https://doi.org/10.3390/nu16091287
Mao S, Zhao A, Jiang H, Yan J, Zhong W, Xun Y, Zhang Y. Patterns of Human Milk Oligosaccharides in Mature Milk Are Associated with Certain Gut Microbiota in Infants. Nutrients. 2024; 16(9):1287. https://doi.org/10.3390/nu16091287
Chicago/Turabian StyleMao, Shuai, Ai Zhao, Hua Jiang, Jingyu Yan, Wuxian Zhong, Yiping Xun, and Yumei Zhang. 2024. "Patterns of Human Milk Oligosaccharides in Mature Milk Are Associated with Certain Gut Microbiota in Infants" Nutrients 16, no. 9: 1287. https://doi.org/10.3390/nu16091287