
Cow’s Milk: A Benefit for Human Health? Omics Tools and Precision Nutrition for Lactose Intolerance Management
 ,
,  , , , and
, , , and 
Abstract
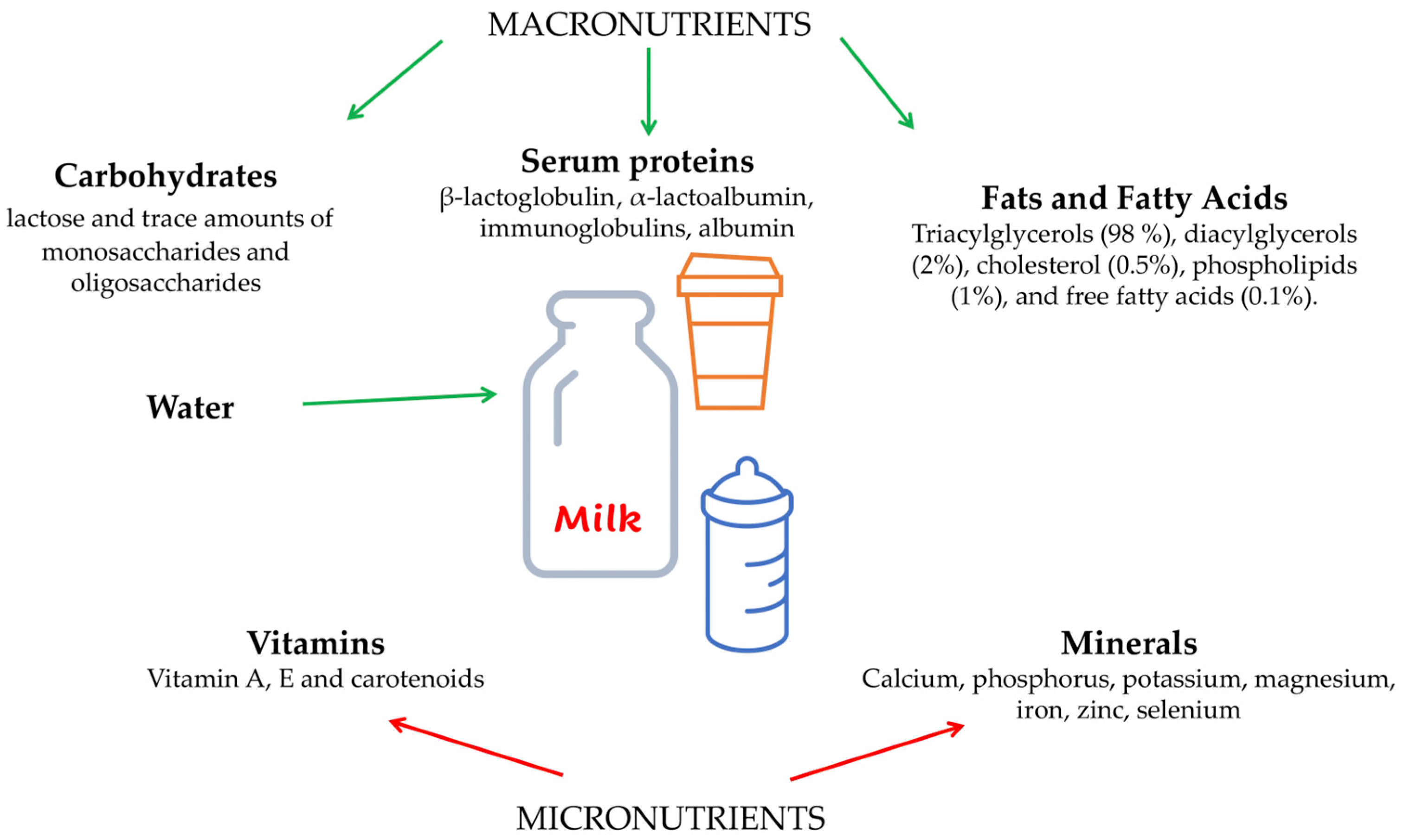
:1. What Is Cow’s Milk Made of?
2. Cow’s Milk Proteins
3. Cow’s Milk Fats and Micronutrients
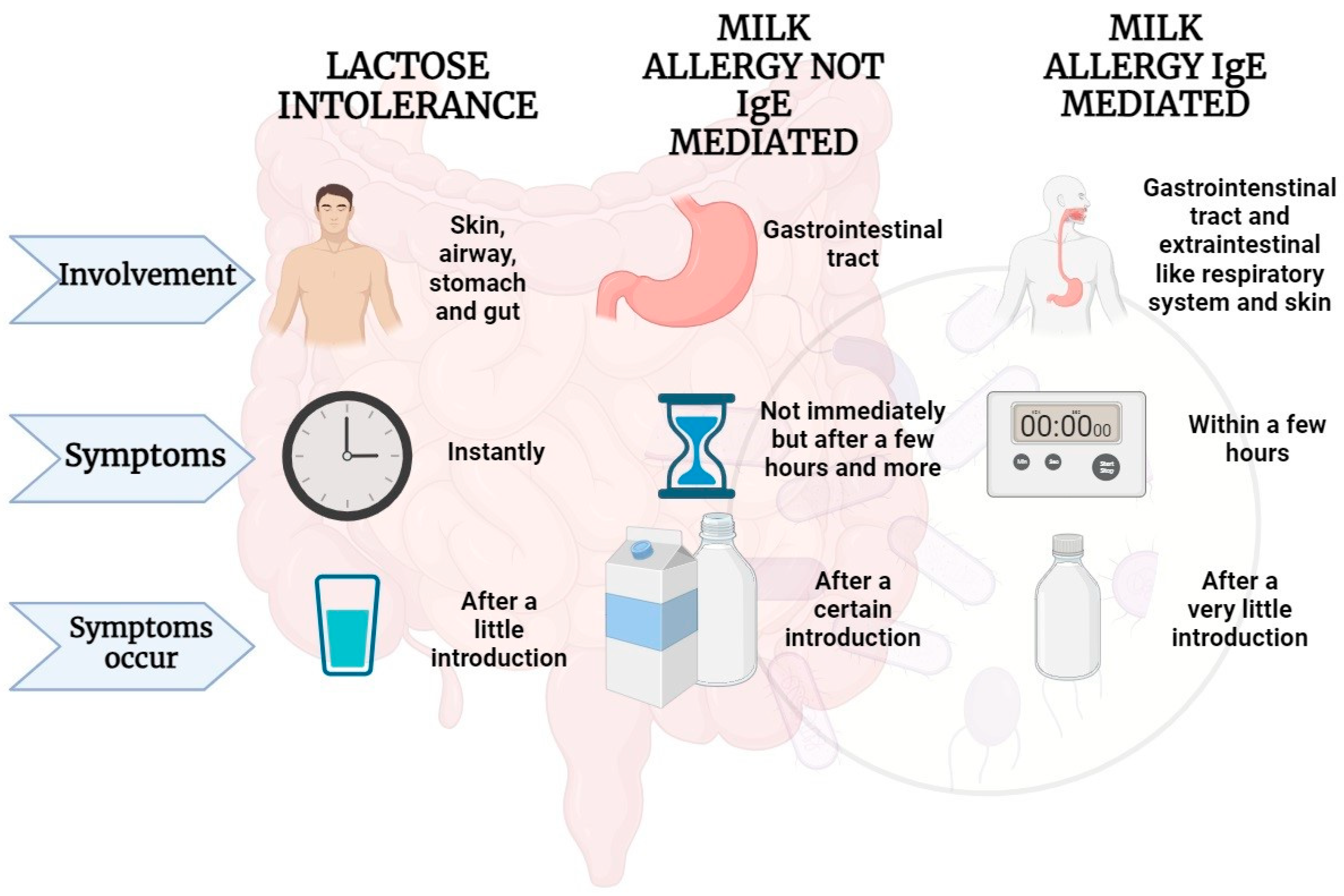
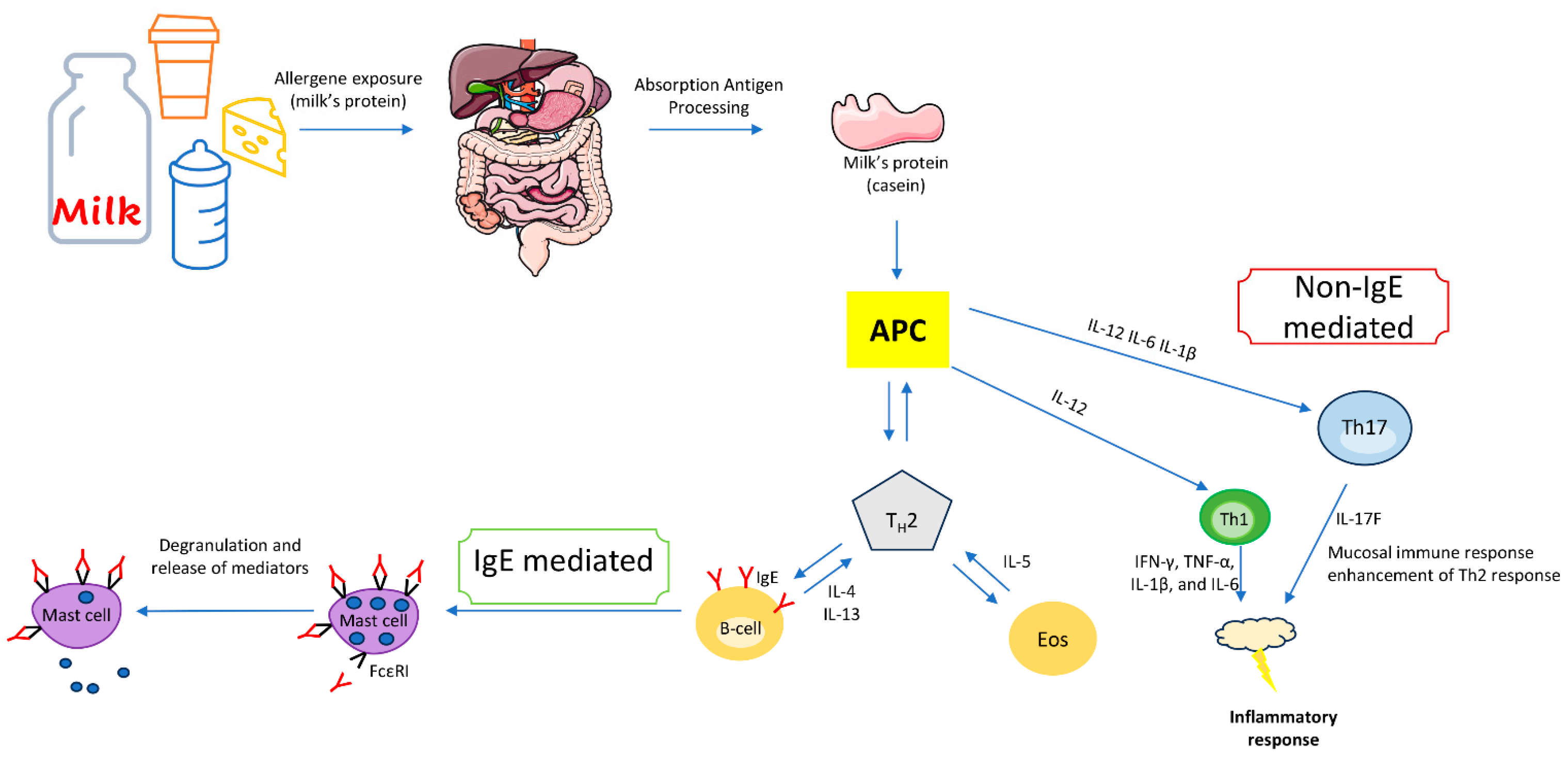
4. IgE-Mediated Immune Response to Cow’s Milk Proteins
5. Non-IgE-Mediated Immune Response to Milk Proteins
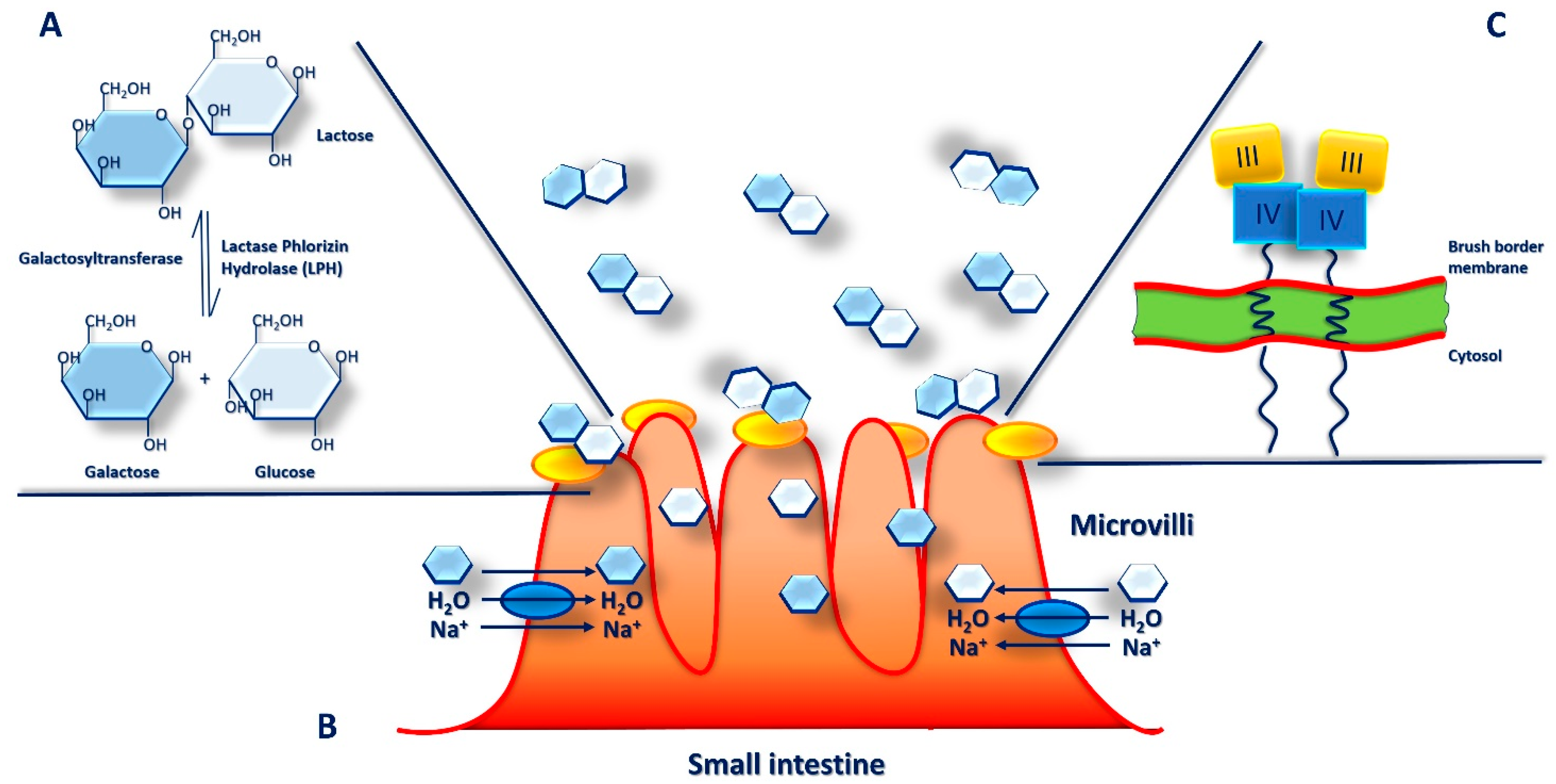
6. Microbiota and Lactose Intolerance
7. Lactase Deficiency and Lactose Intolerance
- Secondary Lactase Deficiency: is a transient condition caused by damage to the intestinal epithelium following to several diseases including infections, small bowel bacterial overgrowth, AIDS, malnutrition, Crohn’s disease, IBS, antibiotic usage, celiac disease, radiation/chemotherapy, rotavirus gastroenteritis, food allergy, and antibiotic usage. However, the reduction in lactase activity is transient and reversible and improves once the intestinal damage is resolved [90,110,111].
- Congenital Lactase Deficiency: is a pediatric autosomal recessive inherited disease that affects infants and leads to a reduced or absent lactase activity at birth. This is an extremely rare disorder, resulting in severe and potentially lethal symptoms with serious difficulties in growth and development after birth [90,110,111].
- Developmental Lactase Deficiency: is a condition observed in premature neonates born between 28 and 37 weeks of gestation. These neonates generally have incomplete development of the enzyme or insufficient intestinal lactase activity. This condition can improve with age and as intestine matures with feeding [90,110,111].
8. Omics Tools: Nutrigenetics and Epigenetics Approaches for Lactose Intolerance Management
9. Precision Nutrition through Metabolomics Approaches
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Heinig, M.J.; Nommsen, L.A.; Peerson, J.M.; Lonnerdal, B.; Dewey, K.G. Energy and protein intakes of breast-fed and formula-fed infants during the first year of life and their association with growth velocity: The DARLING Study. Am. J. Clin. Nutr. 1993, 58, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ayaz, M.; Ajmal, M.; Ellahi, M.Y.; Khalique, A. Antioxidant capacity and fatty acids characterization of heat-treated cow and buffalo milk. Lipids Health Dis. 2017, 16, 163. [Google Scholar] [CrossRef]
- Leduc, A.; Souchet, S.; Gele, M.; Le Provost, F.; Boutinaud, M.; Pascottini, O.; Carvalho, M.; Schyndel, S.; Ticiani, E.; Spricigo, J. Effect of feed restriction on dairy cow milk production. J. Anim. Sci. 2021, 99, 1. [Google Scholar] [CrossRef]
- Foroutan, A.; Guo, A.C.; Vazquez-Fresno, R.; Lipfert, M.; Zhang, L.; Zheng, J.; Badran, H.; Budinski, Z.; Mandal, R.; Ametaj, B.N. Chemical composition of commercial cow’s milk. J. Agric. Food Chem. 2019, 67, 4897–4914. [Google Scholar] [CrossRef]
- Leischner, C.; Egert, S.; Burkard, M.; Venturelli, S. Potential protective protein components of cow’s milk against certain tumor entities. Nutrients 2021, 13, 1974. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Duncan, S.E.; Knowlton, K.F.; Ray, W.K.; Dietrich, A.M. Milk protein composition and stability changes affected by iron in water sources. J. Dairy Sci. 2016, 99, 4206–4219. [Google Scholar] [CrossRef]
- Dyrda-Terniuk, T.; Pryshchepa, O.; Rafińska, K.; Kolankowski, M.; Gołębiowski, A.; Gloc, M.; Dobrucka, R.; Kurzydłowski, K.; Pomastowski, P. Immobilization of silver ions onto casein. Colloids Surf. A Physicochem. Eng. Asp. 2023, 667, 131390. [Google Scholar] [CrossRef]
- Thorning, T.K.; Raben, A.; Tholstrup, T.; Soedamah-Muthu, S.S.; Givens, I.; Astrup, A. Milk and dairy products: Good or bad for human health? An assessment of the totality of scientific evidence. Food Nutr. Res. 2016, 60, 32527. [Google Scholar] [CrossRef]
- Carter, B.; Cheng, N.; Kapoor, R.; Meletharayil, G.; Drake, M. Invited review: Microfiltration-derived casein and whey proteins from milk. J. Dairy Sci. 2021, 104, 2465–2479. [Google Scholar] [CrossRef]
- Carter, B.; DiMarzo, L.; Pranata, J.; Barbano, D.M.; Drake, M. Efficiency of removal of whey protein from sweet whey using polymeric microfiltration membranes. J. Dairy Sci. 2021, 104, 8630–8643. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rico, S.; Mondragón, A.D.C.; López-Santamarina, A.; Cardelle-Cobas, A.; Regal, P.; Lamas, A.; Ibarra, I.S.; Cepeda, A.; Miranda, J.M. A2 Milk: New Perspectives for Food Technology and Human Health. Foods 2022, 11, 2387. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, M.R.; Kapila, R.; Saliganti, V. Consumption of β-casomorphins-7/5 induce inflammatory immune response in mice gut through Th2 pathway. J. Funct. Foods 2014, 8, 150–160. [Google Scholar] [CrossRef]
- Jung, T.H.; Hwang, H.J.; Yun, S.S.; Lee, W.J.; Kim, J.W.; Ahn, J.Y.; Jeon, W.M.; Han, K.S. Hypoallergenic and physicochemical properties of the A2 beta-casein fraction of goat milk. Korean J. Food Sci. Anim. Resour. 2017, 37, 940–947. [Google Scholar] [PubMed]
- Kim, J.; Paik, H.-D.; Yoon, Y.-C.; Park, E. Whey protein inhibits iron overload-induced oxidative stress in rats. J. Nutr. Sci. Vitaminol. 2013, 59, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Chen, T.; Xie, M.-Y.; Luo, J.-Y.; He, J.-J.; Xi, Q.-Y.; Sun, J.-J.; Zhang, Y.-L. Exploration of long noncoding RNA in bovine milk exosomes and their stability during digestion in vitro. J. Dairy Sci. 2019, 102, 6726–6737. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Chen, X.; Zheng, X.; Zhu, H.; Qi, Q.; Liu, S.; Zhang, H.; Che, J. Latest trend of milk derived exosomes: Cargos, functions, and applications. Front. Nutr. 2021, 8, 747294. [Google Scholar] [CrossRef] [PubMed]
- Contarini, G.; Povolo, M. Phospholipids in milk fat: Composition, biological and technological significance, and analytical strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef]
- Ortega-Anaya, J.; Jiménez-Flores, R. Symposium review: The relevance of bovine milk phospholipids in human nutrition—Evidence of the effect on infant gut and brain development. J. Dairy Sci. 2019, 102, 2738–2748. [Google Scholar] [CrossRef]
- Stonehouse, W.; Klingner, B.; McJarrow, P.; Fong, B.; O’callaghan, N. Exploring in vivo dynamics of bovine milk derived gangliosides. Nutrients 2020, 12, 711. [Google Scholar] [CrossRef]
- Benbrook, C.M.; Butler, G.; Latif, M.A.; Leifert, C.; Davis, D.R. Organic production enhances milk nutritional quality by shifting fatty acid composition: A United States–wide, 18-month study. PLoS ONE 2013, 8, e82429. [Google Scholar] [CrossRef] [PubMed]
- Warstedt, K.; Furuhjelm, C.; Fälth-Magnusson, K.; Fagerås, M.; Duchén, K. High levels of omega-3 fatty acids in milk from omega-3 fatty acid-supplemented mothers are related to less immunoglobulin E-associated disease in infancy. Acta Paediatr. 2016, 105, 1337–1347. [Google Scholar] [CrossRef]
- Kasapidou, E.; Basdagianni, Z.; Papatzimos, G.; Papadopoulos, V.; Tsiftsi, E.; Neki, I.; Nigianni, P.-A.; Mitlianga, P. Chemical composition, antioxidant profile and physicochemical properties of commercial non-cocoa-and cocoa-flavoured plant-based milk alternatives. Eur. Food Res. Technol. 2023, 249, 3011–3026. [Google Scholar] [CrossRef]
- Gaucheron, F. The minerals of milk. Reprod. Nutr. Dev. 2005, 45, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, M.D. Calcium intake and urinary stone disease. Transl. Androl. Urol. 2014, 3, 235. [Google Scholar] [PubMed]
- Heravi, A.S.; Michos, E.D. Vitamin D and calcium supplements: Helpful, harmful, or neutral for cardiovascular risk? Methodist DeBakey Cardiovasc. J. 2019, 15, 207. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Clarke, R.; Halsey, J.; Jackson, R.; Lehman, A.; Prince, R.; Lewis, J.; Baron, J.A.; Kroger, H.; Sund, R.; et al. Calcium Supplements Treatment Trialists’ Collaboration. Calcium Supplements and Risk of CVD: A Meta-Analysis of Randomized Trials. Curr. Dev. Nutr. 2023, 7, 100046. [Google Scholar] [CrossRef]
- Drouin-Chartier, J.P.; Brassard, D.; Tessier-Grenier, M.; Côté, J.A.; Labonté, M.È.; Desroches, S.; Couture, P.; Lamarche, B. Systematic Review of the Association between Dairy Product Consumption and Risk of Cardiovascular-Related Clinical Outcomes. Adv. Nutr. 2016, 7, 1026–1040. [Google Scholar] [CrossRef]
- Gudi, S.K. Dairy consumption and risk of type-2 diabetes: The untold story. Ann. Pediatr. Endocrinol. Metab. 2021, 26, 14. [Google Scholar] [CrossRef]
- Gil, H.; Chen, Q.-Y.; Khil, J.; Park, J.; Na, G.; Lee, D.; Keum, N. Milk intake in early life and later cancer risk: A meta-analysis. Nutrients 2022, 14, 1233. [Google Scholar] [CrossRef]
- Arafat, H.M.; Omar, J.; Shafii, N.; Naser, I.A.; Al Laham, N.A.; Muhamad, R.; Al-Astani, T.A.D.; Shaqaliah, A.J.; Shamallakh, O.M.; Shamallakh, K.M. The association between breast cancer and consumption of dairy products: A systematic review. Ann. Med. 2023, 55, 2198256. [Google Scholar] [CrossRef]
- Kumar, A.; Chinnathambi, S.; Kumar, M.; Pandian, G.N. Food Intake and Colorectal Cancer. Nutr. Cancer 2023, 75, 1710–1742. [Google Scholar] [CrossRef] [PubMed]
- Givens, D. MILK Symposium review: The importance of milk and dairy foods in the diets of infants, adolescents, pregnant women, adults, and the elderly. J. Dairy Sci. 2020, 103, 9681–9699. [Google Scholar] [CrossRef]
- Corsello, A.; Pugliese, D.; Gasbarrini, A.; Armuzzi, A. Diet and Nutrients in Gastrointestinal Chronic Diseases. Nutrients 2020, 12, 2693. [Google Scholar] [CrossRef] [PubMed]
- Schoemaker, A.A.; Sprikkelman, A.B.; Grimshaw, K.E.; Roberts, G.; Grabenhenrich, L.; Rosenfeld, L.; Siegert, S.; Dubakiene, R.; Rudzeviciene, O.; Reche, M.; et al. Incidence and natural history of challenge-proven cow’s milk allergy in European children--EuroPrevall birth cohort. Allergy 2015, 70, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy 2014, 69, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Katz, Y.; Rajuan, N.; Goldberg, M.R.; Eisenberg, E.; Heyman, E.; Cohen, A.; Leshno, M. Early exposure to cow’s milk protein is protective against IgE-mediated cow’s milk protein allergy. J. Allergy Clin. Immunol. 2010, 126, 77–82.e1. [Google Scholar] [CrossRef]
- García-Ara, M.C.; Boyano-Martínez, M.T.; Díaz-Pena, J.M.; Martín-Muñoz, M.F.; Martín-Esteban, M. Cow’s milk-specific immunoglobulin E levels as predictors of clinical reactivity in the follow-up of the cow’s milk allergy infants. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2004, 34, 866–870. [Google Scholar] [CrossRef]
- Fiocchi, A.; Terracciano, L.; Bouygue, G.R.; Veglia, F.; Sarratud, T.; Martelli, A.; Restani, P. Incremental prognostic factors associated with cow’s milk allergy outcomes in infant and child referrals: The Milan Cow’s Milk Allergy Cohort study. Ann. Allergy Asthma Immunol. 2008, 101, 166–173. [Google Scholar] [CrossRef]
- Skripak, J.M.; Matsui, E.C.; Mudd, K.; Wood, R.A. The natural history of IgE-mediated cow’s milk allergy. J. Allergy Clin. Immunol. 2007, 120, 1172–1177. [Google Scholar] [CrossRef]
- Fiocchi, A.; Schünemann, H.J.; Brozek, J.; Restani, P.; Beyer, K.; Troncone, R.; Martelli, A.; Terracciano, L.; Bahna, S.L.; Rancé, F.; et al. Diagnosis and Rationale for Action Against Cow’s Milk Allergy (DRACMA): A summary report. J. Allergy Clin. Immunol. 2010, 126, 1119–1128.e12. [Google Scholar] [CrossRef] [PubMed]
- Tsabouri, S.; Douros, K.; Priftis, K.N. Cow’s milk allergenicity. Endocr. Metab. Immune Disord. Drug Targets 2014, 14, 16–26. [Google Scholar] [CrossRef]
- Sampson, H.A. Food allergy. Part 1: Immunopathogenesis and clinical disorders. J. Allergy Clin. Immunol. 1999, 103, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Vickery, B.P.; Chin, S.; Burks, A.W. Pathophysiology of food allergy. Pediatr. Clin. N. Am. 2011, 58, 363–376. [Google Scholar] [CrossRef]
- Luyt, D.; Ball, H.; Makwana, N.; Green, M.R.; Bravin, K.; Nasser, S.M.; Clark, A.T. BSACI guideline for the diagnosis and management of cow’s milk allergy. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2014, 44, 642–672. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, J.M.; Paul, M.; Rothenberg, M.E. Novel immunologic mechanisms in eosinophilic esophagitis. Curr. Opin. Immunol. 2017, 48, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Giannetti, A.; Toschi Vespasiani, G.; Ricci, G.; Miniaci, A.; di Palmo, E.; Pession, A. Cow’s Milk Protein Allergy as a Model of Food Allergies. Nutrients 2021, 13, 1525. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef]
- Savage, J.H.; Lee-Sarwar, K.A.; Sordillo, J.; Bunyavanich, S.; Zhou, Y.; O’Connor, G.; Sandel, M.; Bacharier, L.B.; Zeiger, R.; Sodergren, E.; et al. A prospective microbiome-wide association study of food sensitization and food allergy in early childhood. Allergy 2018, 73, 145–152. [Google Scholar] [CrossRef]
- Roduit, C.; Frei, R.; Depner, M.; Schaub, B.; Loss, G.; Genuneit, J.; Pfefferle, P.; Hyvärinen, A.; Karvonen, A.M.; Riedler, J.; et al. Increased food diversity in the first year of life is inversely associated with allergic diseases. J. Allergy Clin. Immunol. 2014, 133, 1056–1064. [Google Scholar] [CrossRef]
- Perkin, M.R.; Logan, K.; Marrs, T.; Radulovic, S.; Craven, J.; Flohr, C.; Lack, G. Enquiring About Tolerance (EAT) study: Feasibility of an early allergenic food introduction regimen. J. Allergy Clin. Immunol. 2016, 137, 1477–1486.e8. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Maslin, K.; Holloway, J.W.; Silveira, L.J.; Fleischer, D.M.; Dean, T.; Arshad, S.H. Different Measures of Diet Diversity During Infancy and the Association with Childhood Food Allergy in a UK Birth Cohort Study. J. Allergy Clin. Immunol. Pract. 2020, 8, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Koplin, J.J.; Allen, K.J.; Gurrin, L.C.; Peters, R.L.; Lowe, A.J.; Tang, M.L.; Dharmage, S.C. The impact of family history of allergy on risk of food allergy: A population-based study of infants. Int. J. Environ. Res. Public. Health 2013, 10, 5364–5377. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.J.; Hosking, C.S. Food allergy and atopic dermatitis in infancy: An epidemiologic study. Pediatr. Allergy Immunol. 2004, 15, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Boyano-Martínez, T.; García-Ara, C.; Pedrosa, M.; Díaz-Pena, J.M.; Quirce, S. Accidental allergic reactions in children allergic to cow’s milk proteins. J. Allergy Clin. Immunol. 2009, 123, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Burris, A.D.; Burris, J.; Järvinen, K.M. Cow’s milk protein allergy in term and preterm infants: Clinical manifestations, immunologic pathophysiology, and management strategies. NeoReviews 2020, 21, e795–e808. [Google Scholar] [CrossRef] [PubMed]
- Toro-Monjaraz, E.M.; Fonseca-Camarillo, G.; Zárate-Mondragón, F.; Montijo-Barrios, E.; Cadena-León, J.; Avelar-Rodríguez, D.; Ramírez-Mayans, J.; Cervantes-Bustamante, R.; Yamamoto-Furusho, J.K. Differential cytokine expression in the duodenum and rectum of children with non-immunoglobulin e-mediated cow’s milk protein allergy. Dig. Dis. Sci. 2021, 66, 3769–3775. [Google Scholar] [CrossRef] [PubMed]
- Vitaliti, G.; Cimino, C.; Coco, A.; Praticò, A.D.; Lionetti, E. The immunopathogenesis of cow’s milk protein allergy (CMPA). Ital. J. Pediatr. 2012, 38, 35. [Google Scholar] [CrossRef]
- Athie-Morales, V.; Smits, H.H.; Cantrell, D.A.; Hilkens, C.M. Sustained IL-12 signaling is required for Th1 development. J. Immunol. 2004, 172, 61–69. [Google Scholar] [CrossRef]
- Paajanen, L.; Kokkonen, J.; Karttunen, T.J.; Tuure, T.; Korpela, R.; Vaarala, O. Intestinal cytokine mRNA expression in delayed-type cow’s milk allergy. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 470–476. [Google Scholar] [CrossRef]
- Veres, G.; Westerholm-Ormio, M.; Kokkonen, J.; Arato, A.; Savilahti, E. Cytokines and adhesion molecules in duodenal mucosa of children with delayed-type food allergy. J. Pediatr. Gastroenterol. Nutr. 2003, 37, 27–34. [Google Scholar] [PubMed]
- Paajanen, L.; Vaarala, O.; Karttunen, R.; Tuure, T.; Korpela, R.; Kokkonen, J. Increased IFN-γ secretion from duodenal biopsy samples in delayed-type cow’s milk allergy. Pediatr. Allergy Immunol. 2005, 16, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Monin, L.; Gaffen, S.L. Interleukin 17 family cytokines: Signaling mechanisms, biological activities, and therapeutic implications. Cold Spring Harb. Perspect. Biol. 2018, 10, a028522. [Google Scholar] [CrossRef] [PubMed]
- Clayton, F.; Fang, J.C.; Gleich, G.J.; Lucendo, A.J.; Olalla, J.M.; Vinson, L.A.; Lowichik, A.; Chen, X.; Emerson, L.; Cox, K. Eosinophilic esophagitis in adults is associated with IgG4 and not mediated by IgE. Gastroenterology 2014, 147, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Schuyler, A.J.; Wilson, J.M.; Tripathi, A.; Commins, S.P.; Ogbogu, P.U.; Kruzsewski, P.G.; Barnes, B.H.; McGowan, E.C.; Workman, L.J.; Lidholm, J. Specific IgG4 antibodies to cow’s milk proteins in pediatric patients with eosinophilic esophagitis. J. Allergy Clin. Immunol. 2018, 142, 139–148.e12. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Platts-Mills, T.A.; Rispens, T. The developmental history of IgE and IgG4 antibodies in relation to atopy, eosinophilic esophagitis, and the modified TH 2 response. Curr. Allergy Asthma Rep. 2016, 16, 45. [Google Scholar] [CrossRef]
- Pratelli, G.; Tamburini, B.; Carlisi, D.; De Blasio, A.; D’Anneo, A.; Emanuele, S.; Notaro, A.; Affranchi, F.; Giuliano, M.; Seidita, A. Foodomics-Based Approaches Shed Light on the Potential Protective Effects of Polyphenols in Inflammatory Bowel Disease. Int. J. Mol. Sci. 2023, 24, 14619. [Google Scholar] [CrossRef]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, F.R. Intestinal barrier in human health and disease. Int. J. Environ. Res. Public. Health 2021, 18, 12836. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Milani, C.; Duranti, S.; Lugli, G.A.; Bernasconi, S.; Margolles, A.; Di Pierro, F.; Van Sinderen, D.; Ventura, M. The infant gut microbiome as a microbial organ influencing host well-being. Ital. J. Pediatr. 2020, 46, 16. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Tocaa, M.; Fernándezb, A.; Orsic, M.; Tabaccod, O.; Vinderolae, G. Lactose intolerance: Myths and facts. An update. Arch. Argent. Pediatr. 2022, 120, 59–66. [Google Scholar]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lecuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef]
- Corbett, A.J.; Eckle, S.B.; Birkinshaw, R.W.; Liu, L.; Patel, O.; Mahony, J.; Chen, Z.; Reantragoon, R.; Meehan, B.; Cao, H. T-cell activation by transitory neo-antigens derived from distinct microbial pathways. Nature 2014, 509, 361–365. [Google Scholar] [CrossRef]
- Fernando, M.R.; Saxena, A.; Reyes, J.-L.; McKay, D.M. Butyrate enhances antibacterial effects while suppressing other features of alternative activation in IL-4-induced macrophages. Am. J. Physiol.-Gastrointest. Liver Physiol. 2016, 310, G822–G831. [Google Scholar] [CrossRef]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef]
- Tamburini, B.; La Manna, M.P.; La Barbera, L.; Mohammadnezhad, L.; Badami, G.D.; Shekarkar Azgomi, M.; Dieli, F.; Caccamo, N. Immunity and nutrition: The right balance in inflammatory bowel disease. Cells 2022, 11, 455. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Zhong, Y.; Priebe, M.G.; Vonk, R.J.; Huang, C.-Y.; Antoine, J.-M.; He, T.; Harmsen, H.J.; Welling, G.W. The role of colonic microbiota in lactose intolerance. Dig. Dis. Sci. 2004, 49, 78–83. [Google Scholar] [CrossRef]
- Azcarate-Peril, M.A.; Ritter, A.J.; Savaiano, D.; Monteagudo-Mera, A.; Anderson, C.; Magness, S.T.; Klaenhammer, T.R. Impact of short-chain galactooligosaccharides on the gut microbiome of lactose-intolerant individuals. Proc. Natl. Acad. Sci. USA 2017, 114, E367–E375. [Google Scholar] [CrossRef]
- Arnold, J.W.; Simpson, J.B.; Roach, J.; Bruno-Barcena, J.M.; Azcarate-Peril, M.A. Prebiotics for lactose intolerance: Variability in galacto-oligosaccharide utilization by intestinal Lactobacillus rhamnosus. Nutrients 2018, 10, 1517. [Google Scholar] [CrossRef] [PubMed]
- Misselwitz, B.; Butter, M.; Verbeke, K.; Fox, M.R. Update on lactose malabsorption and intolerance: Pathogenesis, diagnosis and clinical management. Gut 2019, 68, 2080–2091. [Google Scholar] [CrossRef] [PubMed]
- Leis, R.; de Castro, M.-J.; de Lamas, C.; Picáns, R.; Couce, M.L. Effects of prebiotic and probiotic supplementation on lactase deficiency and lactose intolerance: A systematic review of controlled trials. Nutrients 2020, 12, 1487. [Google Scholar] [CrossRef] [PubMed]
- Oak, S.J.; Jha, R. The effects of probiotics in lactose intolerance: A systematic review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Gingold-Belfer, R.; Levy, S.; Layfer, O.; Pakanaev, L.; Niv, Y.; Dickman, R.; Perets, T.T. Use of a novel probiotic formulation to alleviate lactose intolerance symptoms—A pilot study. Probiotics Antimicrob. Proteins 2020, 12, 112–118. [Google Scholar] [CrossRef]
- He, T.; Venema, K.; Priebe, M.; Welling, G.; Brummer, R.J.; Vonk, R. The role of colonic metabolism in lactose intolerance. Eur. J. Clin. Investig. 2008, 38, 541–547. [Google Scholar] [CrossRef]
- Di Costanzo, M.; Canani, R.B. Lactose intolerance: Common misunderstandings. Ann. Nutr. Metab. 2018, 73, 30–37. [Google Scholar] [CrossRef]
- D’Auria, E.; Venter, C. Precision medicine in cow’s milk allergy. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Verduci, E.; Zuccotti, G.V.; Peroni, D.G. New Insights in Cow’s Milk and Allergy: Is the Gut Microbiota the Missing Link? Nutrients 2022, 14, 1631. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, S.; Niggemann, B.; Arató, A.; Dias, J.; Heuschkel, R.; Husby, S.; Mearin, M.; Papadopoulou, A.; Ruemmele, F.; Staiano, A. Diagnostic approach and management of cow’s-milk protein allergy in infants and children: ESPGHAN GI Committee practical guidelines. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 221–229. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, E.; Salvatore, S.; Pozzi, E.; Mantegazza, C.; Sartorio, M.U.A.; Pensabene, L.; Baldassarre, M.E.; Agosti, M.; Vandenplas, Y.; Zuccotti, G. Cow’s milk allergy: Immunomodulation by dietary intervention. Nutrients 2019, 11, 1399. [Google Scholar] [CrossRef] [PubMed]
- Forsgård, R.A. Lactose digestion in humans: Intestinal lactase appears to be constitutive whereas the colonic microbiome is adaptable. Am. J. Clin. Nutr. 2019, 110, 273–279. [Google Scholar] [CrossRef]
- Toca, M.; Fernández, A.; Orsi, M.; Tabacco, O.; Vinderola, G. Intolerancia a la lactosa: Mitos y verdades. Actualización. Arch. Argent. Pediatría 2022, 120, 101–110. [Google Scholar]
- Facioni, M.S.; Raspini, B.; Pivari, F.; Dogliotti, E.; Cena, H. Nutritional management of lactose intolerance: The importance of diet and food labelling. J. Transl. Med. 2020, 18, 260. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.; Friesen, C.A. Colonic mucosal inflammatory cells in children and adolescents with lactase deficiency. Pathol.-Res. Pract. 2020, 216, 152971. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Gyawali, R.; Awaisheh, S.S.; Ayivi, R.D.; Silva, R.C.; Subedi, K.; Aljaloud, S.O.; Siddiqui, S.A.; Krastanov, A. Fermented foods and probiotics: An approach to lactose intolerance. J. Dairy Res. 2021, 88, 357–365. [Google Scholar] [CrossRef]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on gut microbiota in patients with lactose intolerance and persisting functional gastrointestinal symptoms: A randomised, double-blind, cross-over study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef]
- Cancarevic, I.; Rehman, M.; Iskander, B.; Lalani, S.; Malik, B.H. Is there a correlation between irritable bowel syndrome and lactose intolerance? Cureus 2020, 12, e6710. [Google Scholar] [CrossRef] [PubMed]
- Alkalay, M.J. Nutrition in patients with lactose malabsorption, celiac disease, and related disorders. Nutrients 2021, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Jansson-Knodell, C.L.; White, M.; Lockett, C.; Xu, H.; Shin, A. Associations of food intolerance with irritable bowel syndrome, psychological symptoms, and quality of life. Clin. Gastroenterol. Hepatol. 2022, 20, 2121–2131.e3. [Google Scholar] [CrossRef] [PubMed]
- Usai-Satta, P.; Lai, M.; Oppia, F. Lactose malabsorption and presumed related disorders: A review of current evidence. Nutrients 2022, 14, 584. [Google Scholar] [CrossRef]
- Efremova, I.; Maslennikov, R.; Poluektova, E.; Vasilieva, E.; Zharikov, Y.; Suslov, A.; Letyagina, Y.; Kozlov, E.; Levshina, A.; Ivashkin, V. Epidemiology of small intestinal bacterial overgrowth. World J. Gastroenterol. 2023, 29, 3400. [Google Scholar] [CrossRef] [PubMed]
- Hodges, J.K.; Cao, S.; Cladis, D.P.; Weaver, C.M. Lactose intolerance and bone health: The challenge of ensuring adequate calcium intake. Nutrients 2019, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- Treister-Goltzman, Y.; Peleg, R. Primary lactase deficiency and bone mineral density in postmenopausal women. Osteoporos. Int. 2019, 30, 527. [Google Scholar] [CrossRef]
- Ratajczak, A.E.; Rychter, A.M.; Zawada, A.; Dobrowolska, A.; Krela-Kaźmierczak, I. Lactose intolerance in patients with inflammatory bowel diseases and dietary management in prevention of osteoporosis. Nutrition 2021, 82, 111043. [Google Scholar] [CrossRef]
- Zingone, F.; Bertin, L.; Maniero, D.; Palo, M.; Lorenzon, G.; Barberio, B.; Ciacci, C.; Savarino, E.V. Myths and Facts about Food Intolerance: A Narrative Review. Nutrients 2023, 15, 4969. [Google Scholar] [CrossRef]
- Szilagyi, A.; Ishayek, N. Lactose intolerance, dairy avoidance, and treatment options. Nutrients 2018, 10, 1994. [Google Scholar] [CrossRef]
- Malik, T.F.; Panuganti, K.K. Lactose intolerance. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Anguita-Ruiz, A.; Aguilera, C.M.; Gil, Á. Genetics of lactose intolerance: An updated review and online interactive world maps of phenotype and genotype frequencies. Nutrients 2020, 12, 2689. [Google Scholar] [CrossRef] [PubMed]
- Wanes, D.; Husein, D.M.; Naim, H.Y. Congenital lactase deficiency: Mutations, functional and biochemical implications, and future perspectives. Nutrients 2019, 11, 461. [Google Scholar] [CrossRef] [PubMed]
- De Luca, P.; Iaconis, D.; Biffali, E.; Enza, C.; de Magistris, L.; Riegler, G.; Pappalardo, D.; Amato, M.R.; Iardino, P.; Montanino, C. Development of a novel SNP assay to detect lactase persistence associated genetic variants. Mol. Biol. Rep. 2021, 48, 7087–7093. [Google Scholar] [CrossRef] [PubMed]
- Marten, L.M.; Wanes, D.; Stellbrinck, T.; Santer, R.; Naim, H.Y. Hypomorphic variants of lactase-phlorizin hydrolase in congenital lactase deficiency are trafficking incompetent and functionally inactive. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2022, 1868, 166338. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.T.; Lei, Y.; Mitchell, L.E.; Sharma, S.V.; Swartz, M.D.; Waller, D.K.; Finnell, R.H.; Benjamin, R.H.; Browne, M.L.; Canfield, M.A. Maternal lactase polymorphism (rs4988235) is associated with neural tube defects in offspring in the National Birth Defects Prevention Study. J. Nutr. 2019, 149, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Kowalówka, M.; Kosewski, G.; Lipiński, D.; Przysławski, J. A Comprehensive Look at the-13910 C>T LCT Gene Polymorphism as a Molecular Marker for Vitamin D and Calcium Levels in Young Adults in Central and Eastern Europe: A Preliminary Study. Int. J. Mol. Sci. 2023, 24, 10191. [Google Scholar] [CrossRef] [PubMed]
- Domżał-Magrowska, D.; Kowalski, M.K.; Małecka-Wojciesko, E. The incidence of adult type hypolactasia in patients with irritable bowel syndrome. Gastroenterol. Rev. Przegląd Gastroenterol. 2023, 18, 110–114. [Google Scholar] [CrossRef]
- Comerford, K.B.; Pasin, G. Gene–dairy food interactions and health outcomes: A review of nutrigenetic studies. Nutrients 2017, 9, 710. [Google Scholar] [CrossRef]
- Aruoma, O.I.; Hausman-Cohen, S.; Pizano, J.; Schmidt, M.A.; Minich, D.M.; Joffe, Y.; Brandhorst, S.; Evans, S.J.; Brady, D.M. Personalized nutrition: Translating the science of nutrigenomics into practice: Proceedings from the 2018 American College of Nutrition Meeting. J. Am. Coll. Nutr. 2019, 38, 287–301. [Google Scholar] [CrossRef]
- Ferguson, L.R.; De Caterina, R.; Görman, U.; Allayee, H.; Kohlmeier, M.; Prasad, C.; Choi, M.S.; Curi, R.; De Luis, D.A.; Gil, Á. Guide and position of the international society of nutrigenetics/nutrigenomics on personalised nutrition: Part 1-fields of precision nutrition. J. Nutr. Nutr. 2016, 9, 12–27. [Google Scholar] [CrossRef]
- Robles, L.; Priefer, R. Lactose intolerance: What your breath can tell you. Diagnostics 2020, 10, 412. [Google Scholar] [CrossRef] [PubMed]
- Tomczonek-Moruś, J.; Wojtasik, A.; Zeman, K.; Smolarz, B.; Bąk-Romaniszyn, L. 13910C>T and 22018G>A LCT gene polymorphisms in diagnosing hypolactasia in children. United Eur. Gastroenterol. J. 2019, 7, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Mattar, R.; de Campos Mazo, D.F.; Carrilho, F.J. Lactose intolerance: Diagnosis, genetic, and clinical factors. Clin. Exp. Gastroenterol. 2012, 5, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Floris, M.; Cano, A.; Porru, L.; Addis, R.; Cambedda, A.; Idda, M.L.; Steri, M.; Ventura, C.; Maioli, M. Direct-to-consumer nutrigenetics testing: An overview. Nutrients 2020, 12, 566. [Google Scholar] [CrossRef]
- Guasch-Ferré, M.; Dashti, H.S.; Merino, J. Nutritional genomics and direct-to-consumer genetic testing: An overview. Adv. Nutr. 2018, 9, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Marietta, C.; McGuire, A.L. Currents in contemporary ethics Direct-to-consumer genetic testing: Is it the practice of medicine? J. Law Med. Ethics 2009, 37, 369–374. [Google Scholar] [CrossRef] [PubMed]
- De, S.; Pietilä, A.-M.; Iso-Touru, T.; Hopia, A.; Tahvonen, R.; Vähäkangas, K. Information provided to consumers about direct-to-consumer nutrigenetic testing. Public Health Genom. 2020, 22, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Misselwitz, B.; Pohl, D.; Frühauf, H.; Fried, M.; Vavricka, S.R.; Fox, M. Lactose malabsorption and intolerance: Pathogenesis, diagnosis and treatment. United Eur. Gastroenterol. J. 2013, 1, 151–159. [Google Scholar] [CrossRef]
- Zheng, X.; Chu, H.; Cong, Y.; Deng, Y.; Long, Y.; Zhu, Y.; Pohl, D.; Fried, M.; Dai, N.; Fox, M. Self-reported lactose intolerance in clinic patients with functional gastrointestinal symptoms: Prevalence, risk factors, and impact on food choices. Neurogastroenterol. Motil. 2015, 27, 1138–1146. [Google Scholar] [CrossRef]
- Tandy-Connor, S.; Guiltinan, J.; Krempely, K.; LaDuca, H.; Reineke, P.; Gutierrez, S.; Gray, P.; Davis, B.T. False-positive results released by direct-to-consumer genetic tests highlight the importance of clinical confirmation testing for appropriate patient care. Genet. Med. 2018, 20, 1515–1521. [Google Scholar] [CrossRef]
- Kuchay, R.A.H. New insights into the molecular basis of lactase non-persistence/persistence: A brief review. Drug Discov. Ther. 2020, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Labrie, V.; Buske, O.J.; Oh, E.; Jeremian, R.; Ptak, C.; Gasiūnas, G.; Maleckas, A.; Petereit, R.; Žvirbliene, A.; Adamonis, K. Lactase nonpersistence is directed by DNA-variation-dependent epigenetic aging. Nat. Struct. Mol. Biol. 2016, 23, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Leseva, M.N.; Grand, R.J.; Klett, H.; Boerries, M.; Busch, H.; Binder, A.M.; Michels, K.B. Differences in DNA methylation and functional expression in lactase persistent and non-persistent individuals. Sci. Rep. 2018, 8, 5649. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Goda, T.; Motohashi, Y.; Sakuma, K. The specific expression patterns of lactase, sucrase and calbindin-D9k in weaning rats are regulated at the transcriptional level. J. Nutr. Sci. Vitaminol. 2004, 50, 265–271. [Google Scholar] [CrossRef]
- Motohashi, Y.; Fukushima, A.; Kondo, T.; Sakuma, K. Lactase decline in weaning rats is regulated at the transcriptional level and not caused by termination of milk ingestion. J. Nutr. 1997, 127, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Maiuri, L.; Fusco, M.I.; Salvati, V.M.; Fuccio, A.; Auricchio, S.; Mantei, N.; Zecca, L.; Gloor, S.M.; Semenza, G. Lactase persistence versus decline in human adults: Multifactorial events are involved in down-regulation after weaning. Gastroenterology 1997, 112, 1506–1514. [Google Scholar] [CrossRef]
- Xu, Y.-J.; Wu, X. Foodomics in microbiological investigations. Curr. Opin. Food Sci. 2015, 4, 51–55. [Google Scholar] [CrossRef]
- Pimentel, G.; Burton, K.J.; Rosikiewicz, M.; Freiburghaus, C.; von Ah, U.; Münger, L.H.; Pralong, F.P.; Vionnet, N.; Greub, G.; Badertscher, R. Blood lactose after dairy product intake in healthy men. Br. J. Nutr. 2017, 118, 1070–1077. [Google Scholar] [CrossRef]
- Vionnet, N.; Münger, L.H.; Freiburghaus, C.; Burton, K.J.; Pimentel, G.; Pralong, F.P.; Badertscher, R.; Vergères, G. Assessment of lactase activity in humans by measurement of galactitol and galactonate in serum and urine after milk intake. Am. J. Clin. Nutr. 2019, 109, 470–477. [Google Scholar] [CrossRef]
- Bush, C.L.; Blumberg, J.B.; El-Sohemy, A.; Minich, D.M.; Ordovás, J.M.; Reed, D.G.; Behm, V.A.Y. Toward the definition of personalized nutrition: A proposal by the American Nutrition Association. J. Am. Coll. Nutr. 2020, 39, 5–15. [Google Scholar] [CrossRef]
- Toro-Martín, D.; Arsenault, B.J.; Després, J.-P.; Vohl, M.-C. Precision nutrition: A review of personalized nutritional approaches for the prevention and management of metabolic syndrome. Nutrients 2017, 9, 913. [Google Scholar] [CrossRef] [PubMed]
- Sales, N.M.R.; Pelegrini, P.B.; Goersch, M. Nutrigenomics: Definitions and advances of this new science. J. Nutr. Metab. 2014, 2014, 202759. [Google Scholar] [CrossRef] [PubMed]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361, bmj.k2173. [Google Scholar] [CrossRef] [PubMed]
- Authority, E.F.S. Dietary Reference Values for Nutrients Summary Report; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 2397–8325. [Google Scholar]
- Dekker, P.J.; Koenders, D.; Bruins, M.J. Lactose-free dairy products: Market developments, production, nutrition and health benefits. Nutrients 2019, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Fassio, F.; Facioni, M.S.; Guagnini, F. Lactose maldigestion, malabsorption, and intolerance: A comprehensive review with a focus on current management and future perspectives. Nutrients 2018, 10, 1599. [Google Scholar] [CrossRef]
- Suchy, F.J.; Brannon, P.M.; Carpenter, T.O.; Fernandez, J.R.; Gilsanz, V.; Gould, J.B.; Hall, K.; Hui, S.L.; Lupton, J.; Mennella, J.; et al. NIH consensus development conference statement: Lactose intolerance and health. NIH Consens. State Sci. Statements 2010, 27, 1–27. [Google Scholar] [PubMed]
- Nicklas, T.A.; Qu, H.; Hughes, S.O.; He, M.; Wagner, S.E.; Foushee, H.R.; Shewchuk, R.M. Self-perceived lactose intolerance results in lower intakes of calcium and dairy foods and is associated with hypertension and diabetes in adults. Am. J. Clin. Nutr. 2011, 94, 191–198. [Google Scholar] [CrossRef]
- Heaney, R.P. Calcium, dairy products and osteoporosis. J. Am. Coll. Nutr. 2000, 19, 83S–99S. [Google Scholar] [CrossRef]
- Lactose Intolerance—Treatment—NHS. 2020. Available online: https://www.nhs.uk/conditions/lactose-intolerance/treatment/ (accessed on 27 December 2023).
- Guide: Nutritional Characteristics—Parmigiano Reggiano. 2020. Available online: https://www.parmigianoreggiano.com/product-guide-nutritional-characteristics/#2 (accessed on 27 December 2023).
- Szilagyi, A.; Shrier, I.; Heilpern, D.; Je, J.S.; Park, S.; Chong, G.; Lalonde, C.; Cote, L.-F.; Lee, B. Differential impact of lactose/lactase phenotype on colonic microflora. Can. J. Gastroenterol. Hepatol. 2010, 24, 373–379. [Google Scholar] [CrossRef]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef]
- Hertzler, S.; Savaiano, D.A.; Dilk, A.; Jackson, K.A.; Bhriain, S.N.; Suarez, F.L. Nutrient considerations in lactose intolerance. Nutr. Prev. Treat. Dis. 2017, 40, 875–892. [Google Scholar]
- Silanikove, N.; Leitner, G.; Merin, U. The interrelationships between lactose intolerance and the modern dairy industry: Global perspectives in evolutional and historical backgrounds. Nutrients 2015, 7, 7312–7331. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.C.; Lerebours, E.C.; Lin, W.J.; Harlander, S.K.; Berrada, N.M.; Antoine, J.M.; Savaiano, D.A. Strains and species of lactic acid bacteria in fermented milks (yogurts): Effect on in vivo lactose digestion. Am. J. Clin. Nutr. 1991, 54, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Kies, A.K. Authorised EU health claims related to the management of lactose intolerance: Reduced lactose content, dietary lactase supplements and live yoghurt cultures. In Foods, Nutrients and Food Ingredients with Authorised EU Health Claims; Woodhead Publishing: Sawston, UK, 2014. [Google Scholar]
- Mutch, D.M.; Wahli, W.; Williamson, G. Nutrigenomics and nutrigenetics: The emerging faces of nutrition. FASEB J. 2005, 19, 1602–1616. [Google Scholar] [CrossRef] [PubMed]
- Röttger-Wirtz, S.; Alie, D. Personalised Nutrition: The EU’s Fragmented Legal Landscape and the Overlooked Implications of EU Food Law. Eur. J. Risk Regul. 2021, 12, 212–235. [Google Scholar] [CrossRef]
- Nordström, K.; Goossens, J. Personalized nutrition and social justice: Ethical considerations within four future scenarios applying the perspective of Nussbaum’s capabilities approach. J. Agric. Environ. Ethics 2016, 29, 5–22. [Google Scholar] [CrossRef]
- Casellas, F.; Aparici, A.; Pérez, M.; Rodríguez, P. Perception of lactose intolerance impairs health-related quality of life. Eur. J. Clin. Nutr. 2016, 70, 1068–1072. [Google Scholar] [CrossRef]
- Enko, D.; Meinitzer, A.; Brandmayr, W.; Halwachs-Baumann, G.; Schnedl, W.J.; Kriegshäuser, G. Association between increased plasma levels of homocysteine and depression observed in individuals with primary lactose malabsorption. PLoS ONE 2018, 13, e0202567. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TEST | APPLICATION | TEST PRINCIPLE |
|---|---|---|
| Hydrogen breath test (HBT) | Test of choice for the diagnosis of LM/LI | Detection of increase H2 in expiratory air after lactose intake |
| Lactose tolerance test | LM epidemiology, low sensitivity and specificity | Increase in blood sugar after lactose intake |
| Lactase activity of jejunum | If gastroscopy is carried out for other investigations, invasive and expensive | Evaluation of lactase enzymatic activity in duodenal biopsy sample |
| Genetic test | LD/LNP | Test for detection of −13910:C>T polymorphism |
| Serum gaxilose or urine galactose test | Used for the diagnosis of small intestine diseases (intestinal malabsorption) | Evaluation of D-xylose in plasma or galactose in urine after cleavage by lactase of 4-galactosylxylose oral administration |
| SNP | VARIANT | GEOGRAPHIC REGION |
|---|---|---|
| rs4988235 | −13910:C>T | Europe |
| rs869051967 | −14009:T>G | Middle East |
| rs4988236 | −13908:G>A | Far East |
| rs773131166 | −13914:C>T | East Europe |
| rs41380347 | −13915:A>C | Middle East |
| rs41525747 | −13907:G>C | Middle East |
| rs820486563 | −14009:T>G | Ethiopia and Sudan |
| rs182549 | –22018:G>A | Europe |
| rs41456145 | −13913:A>G | Africa |
| rs145946881 | −14010:C>G | Kenya, Tanzania and South Africa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pratelli, G.; Tamburini, B.; Badami, G.D.; Lo Pizzo, M.; De Blasio, A.; Carlisi, D.; Di Liberto, D. Cow’s Milk: A Benefit for Human Health? Omics Tools and Precision Nutrition for Lactose Intolerance Management. Nutrients 2024, 16, 320. https://doi.org/10.3390/nu16020320
Pratelli G, Tamburini B, Badami GD, Lo Pizzo M, De Blasio A, Carlisi D, Di Liberto D. Cow’s Milk: A Benefit for Human Health? Omics Tools and Precision Nutrition for Lactose Intolerance Management. Nutrients. 2024; 16(2):320. https://doi.org/10.3390/nu16020320
Chicago/Turabian StylePratelli, Giovanni, Bartolo Tamburini, Giusto Davide Badami, Marianna Lo Pizzo, Anna De Blasio, Daniela Carlisi, and Diana Di Liberto. 2024. "Cow’s Milk: A Benefit for Human Health? Omics Tools and Precision Nutrition for Lactose Intolerance Management" Nutrients 16, no. 2: 320. https://doi.org/10.3390/nu16020320