
Molecular Oxygen Levels and Percentages of DNA Damage in TPN Patients


 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oxygen Consumption Assay
2.2. Estimation of Oxidative Damage Based on Genomic DNA Digestion by Fpg Protein
3. Statistical Analysis
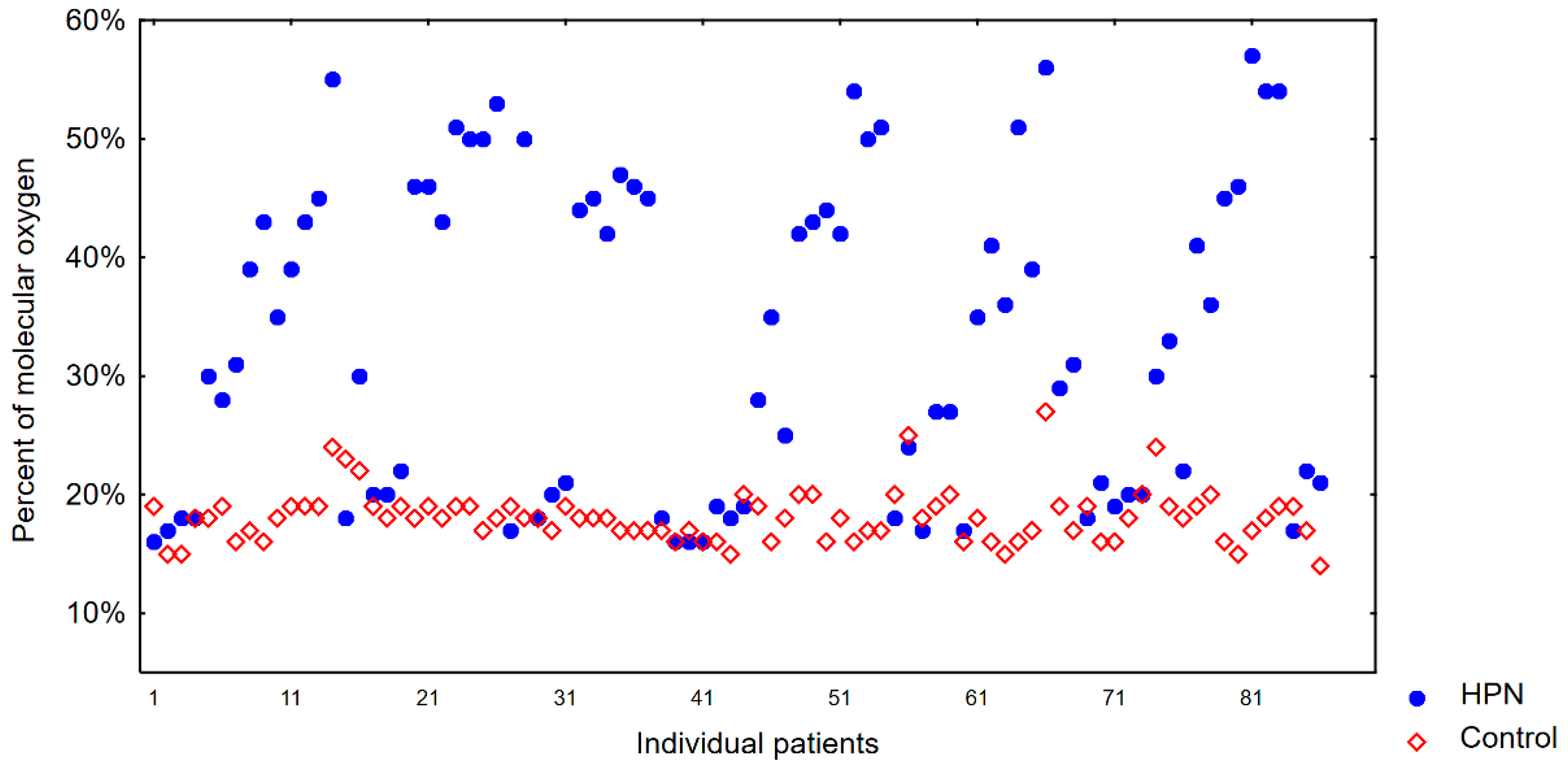
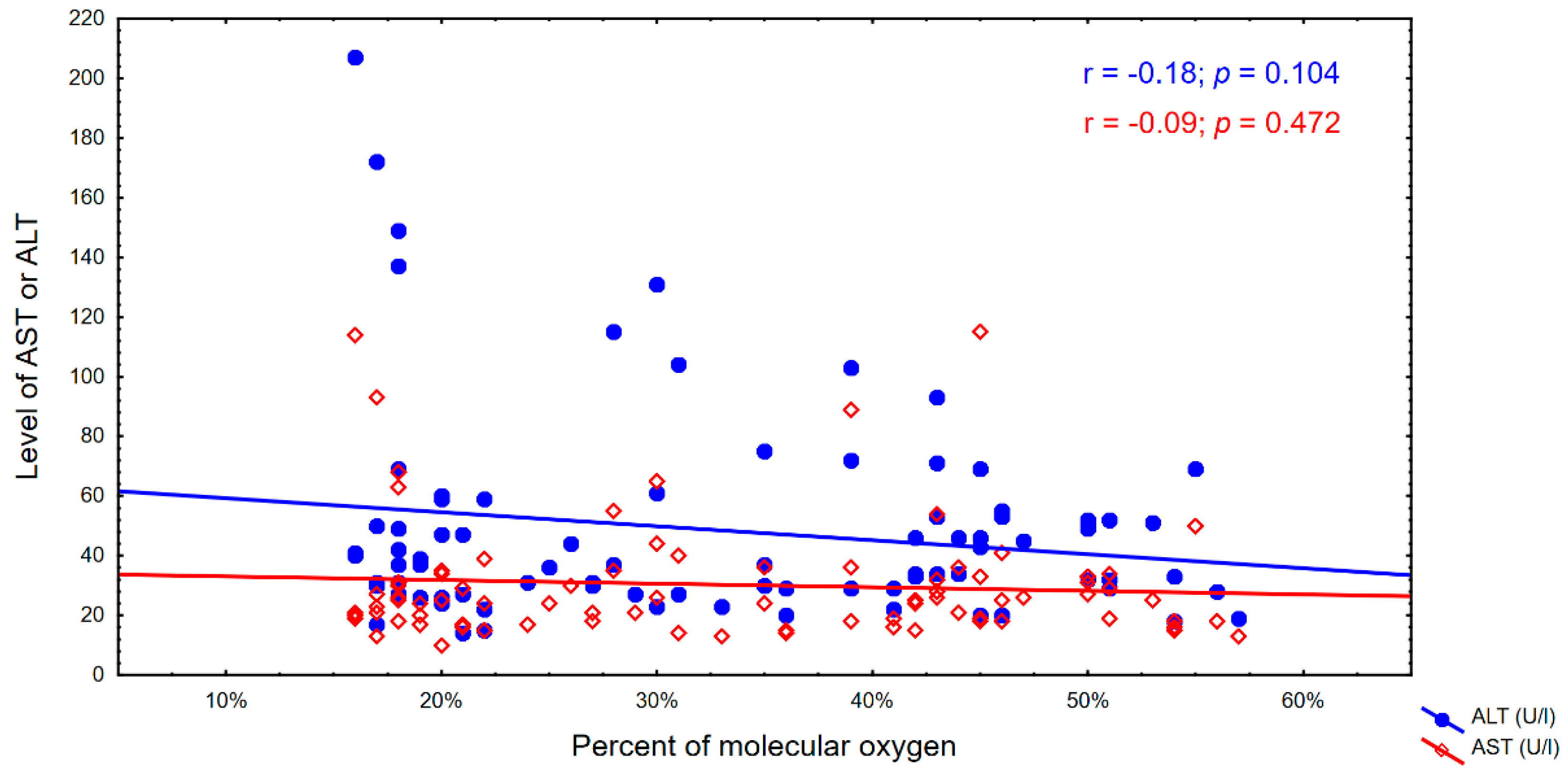
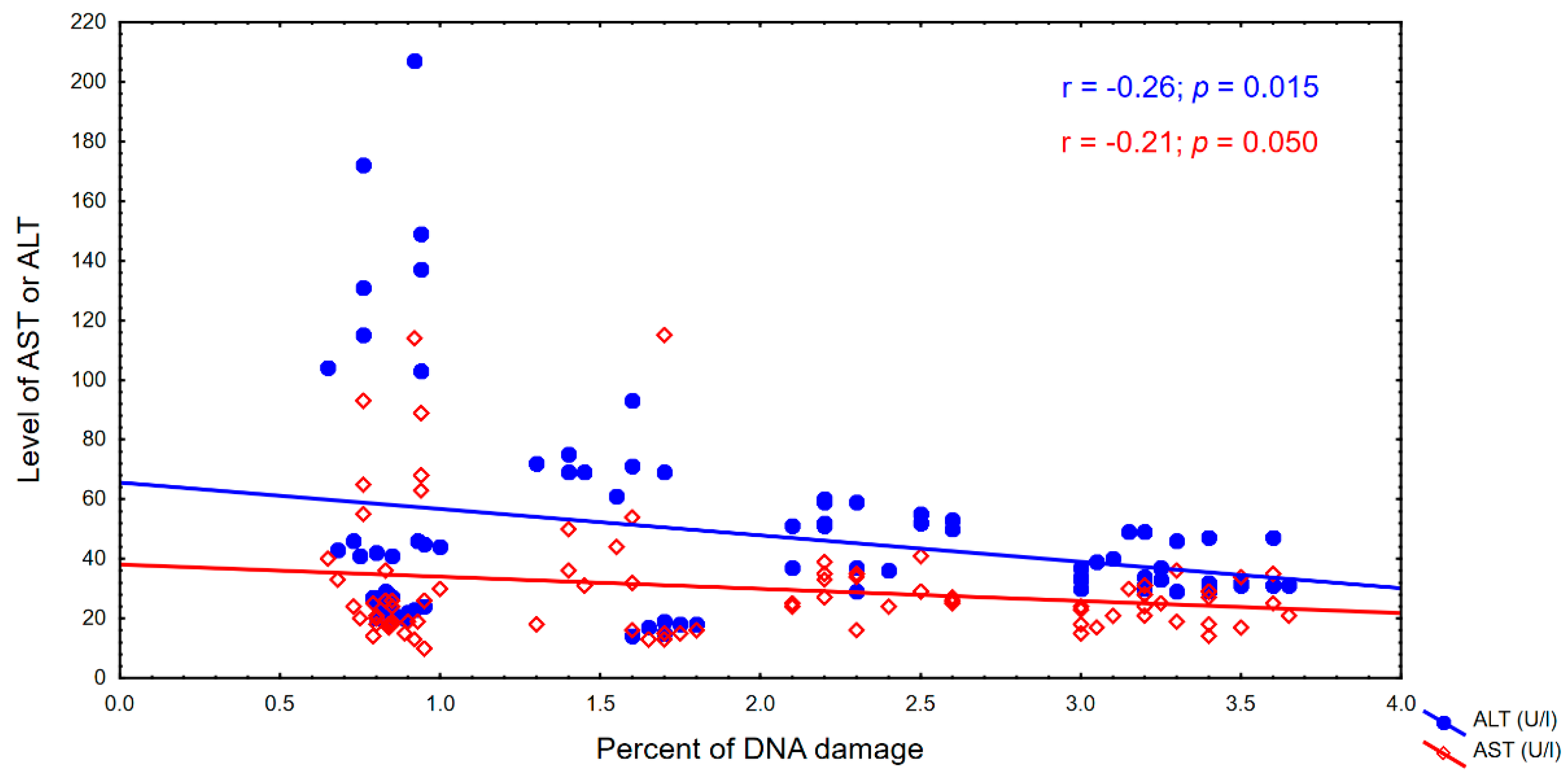
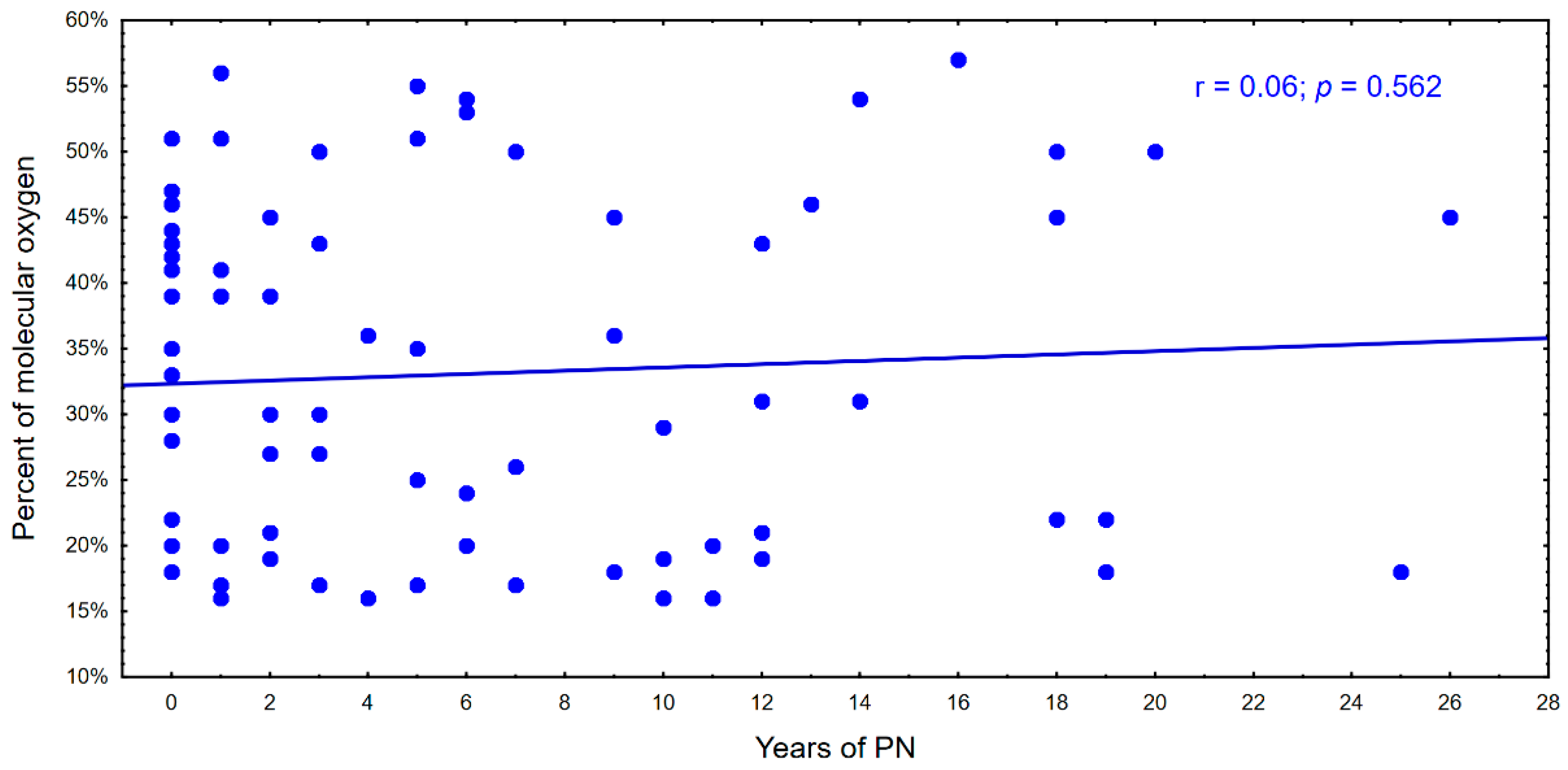
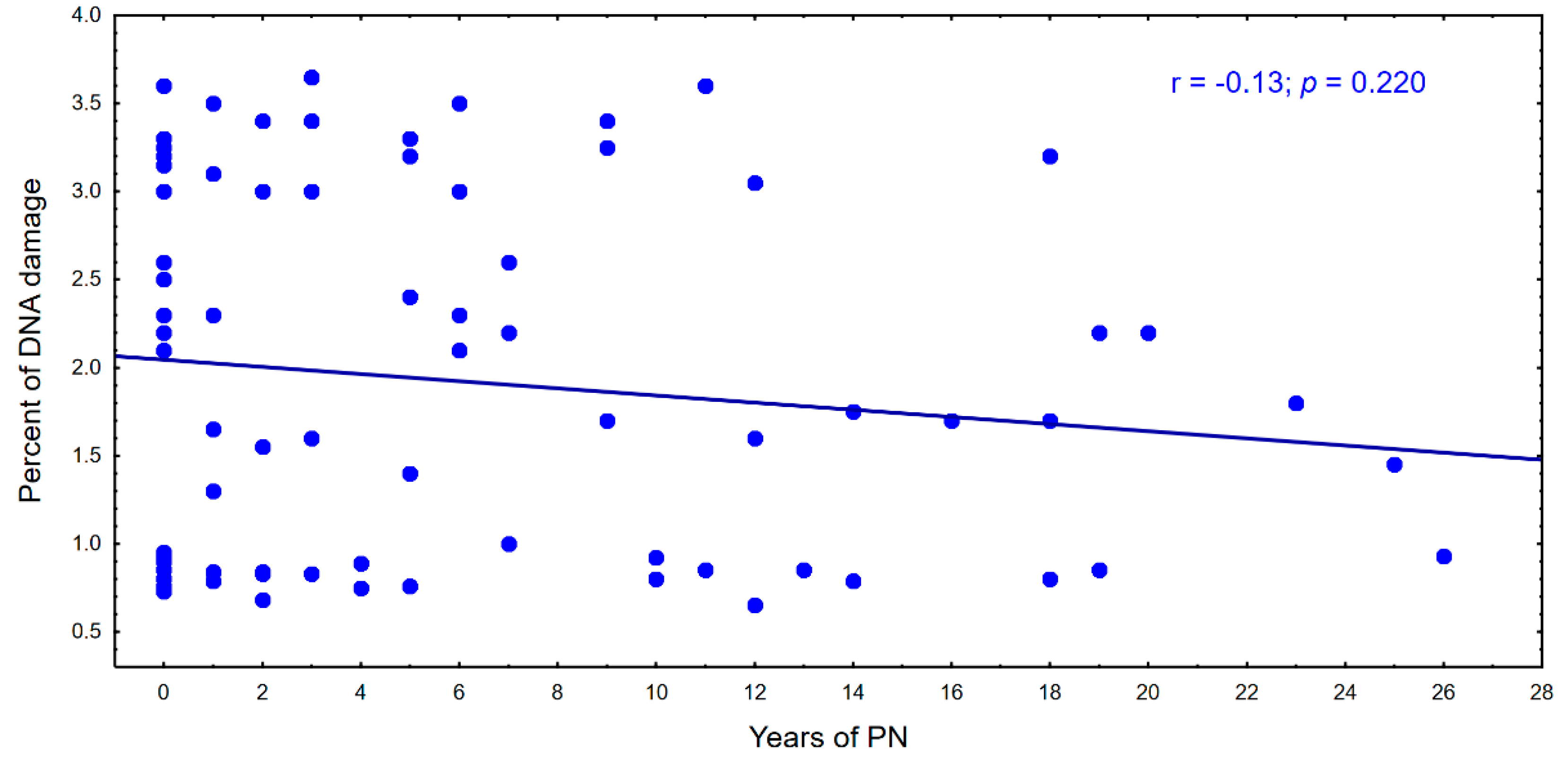
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cederholm, T.; Barazzoni, R.; Austin, P.; Ballmer, P.; Biolo, G.; Bischoff, S.C.; Compher, C.; Correia, I.; Higashiguchi, T.; Holst, M.; et al. ESPEN guidelines on definitions and terminology of clinical nutrition. Clin. Nutr. 2017, 36, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Triantafillidis, J.K.; Papalois, A.E. The role of total parenteral nutrition in inflammatory bowel disease: Current aspects. Scand. J. Gastroenterol. 2014, 49, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Chew, K.S.; Ng, R.T.; El Kasmi, K.; Sokol, R.J. Intestinal failure-associated liver disease (IFALD): Insights into pathogenesis and advances in management. Hepatol. Int. 2020, 14, 305–316. [Google Scholar] [CrossRef]
- Elfawy, H.A.; Das, B. Life Sciences Crosstalk between mitochondrial dysfunction, oxidative stress, and age related neurodegenerative disease: Etiologies and therapeutic strategies. Life Sci. 2019, 218, 165–184. [Google Scholar] [CrossRef]
- Güiza, F.; Vanhorebeek, I.; Verstraete, S.; Verlinden, I.; Derese, I.; Ingels, C.; Dulfer, K.; Verbruggen, S.C.; Garcia Guerra, G.; Joosten, K.F.; et al. Effect of early parenteral nutrition during paediatric critical illness on DNA methylation as a potential mediator of impaired neurocognitive developement: A pre-planned secondary analysis of the PEPaNIC international, randomised controlled trial. Lancet Respir. Med. 2020, 8, 288–303. [Google Scholar] [CrossRef]
- Lejri, I.; Agapouda, A.; Grimm, A.; Eckert, A. Mitochondria- and Oxidative Stress-Targeting Substances in Cognitive Decline-Related Disorders: From Molecular Mechanisms to Clinical Evidence. Oxidative Med. Cell Longev. 2019, 2019, 9695412. [Google Scholar] [CrossRef]
- Wesselink, E.; Koekkoek, W.A.C.; Grefte, S.; Witkamp, R.F.; van Zanten, A.R.H. Feeding mitochondria: Potential role of nutritional components to improve critical illness convalescence. Clin. Nutr. 2019, 38, 982–995. [Google Scholar] [CrossRef]
- Ferrucci-Da Silva, C.; Zhan, L.; Shen, J.; Kong, B.; Campbell, M.J.; Memon, N.; Hegyi, T.; Lu, L.; Guo, G.L. Effects of total parenteral nutrition on drug metabolism gene expression in mice. Acta Pharm. Sin. B 2020, 10, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, J.-C.; Kang, W.-M.; Ma, Z.-Q. Superiority of a fish oil–enriched emulsion to medium-chain triacylglycerols/long-chain triacylglycerols in gastrointestinal surgery patients: A randomized clinical trial. Nutrition 2012, 28, 623–629. [Google Scholar] [CrossRef]
- Navarro, A.; Boveris, A. Rat brain and liver mitochondria develop oxidative stress and lose enzymatic activities on aging. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2004, 287, R1244–R1249. [Google Scholar] [CrossRef]
- Navarro, A.; Boveris, A. The mitochondrial energy transduction system and the aging process. Am. J. Physiol.-Cell Physiol. 2007, 292, C670–C686. [Google Scholar] [CrossRef] [PubMed]
- Kosek, V.; Heczkova, M.; Novak, F.; Meisnerova, E.; Novákova, O.; Zelenka, J.; Bechynska, K.; Vrzacova, N.; Suttnar, J.; Hlavackova, A.; et al. The ω-3 Polyunsaturated Fatty Acids and Oxidative Stress in Long-Term Parenteral Nutrition Dependent Adult Patients: Functional Lipidomics Approach. Nutrients 2020, 12, 2351. [Google Scholar] [CrossRef] [PubMed]
- Unal, S.; Demirel, N.; Erol, S.; Isik, D.U.; Kulali, F.; Iyigun, F.; Bas, A.Y. Effects of two different lipid emulsions on morbidities and oxidant stress statuses in preterm infants: An observational study. J. Matern.-Fetal Neonatal Med. 2018, 31, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Maehre, H.K.; Jensen, I.J.; Elvevoll, E.O.; Eilertsen, K.E. Omega-3 Fatty Acids and Cardiovascular Diseases: Effects, Mechanisms and Dietary Relevance. Int. J. Mol. Sci. 2015, 16, 22636–22661. [Google Scholar] [CrossRef] [PubMed]
- Repetto, M.G.; Ossani, G.; Monserrat, A.J.; Boveris, A. Oxidative damage: The biochemical mechanism of cellular injury and necrosis in choline deficiency. Exp. Mol. Pathol. 2010, 88, 143–149. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Cieśla, J.; Komisarski, M.; Kuśmierek, J.T.; Tudek, B. Long-chain adducts of trans-4-hydroxy-2-nonenal to DNA bases cause recombination, base substitutions and frameshift mutations in M13 phage. Mutat. Res. Mol. Mech. Mutagen. 2004, 550, 33–48. [Google Scholar] [CrossRef]
- Konieczka, P.; Barszcz, M.; Kowalczyk, P.; Szlis, M.; Jankowski, J. The potential of acetylsalicylic acid and vitamin E in modulating inflammatory cascades in chickens under lipopolysaccharide-induced inflammation. Vet. Res. 2019, 50, 65. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Koszelewski, D.; Gawdzik, B.; Samsonowicz-Górski, J.; Kramkowski, K.; Wypych, A.; Lizut, R.; Ostaszewski, R. Promiscuous Lipase-Catalyzed Markovnikov Addition of H-Phosphites to Vinyl Esters for the Synthesis of Cytotoxic α-Acyloxy Phosphonate Derivatives. Materials 2022, 15, 1975. [Google Scholar] [CrossRef]
- Koszelewski, D.; Kowalczyk, P.; Śmigielski, P.; Samsonowicz-Górski, J.; Kramkowski, K.; Wypych, A.; Szymczak, M.; Ostaszewski, R. Relationship between Structure and Antibacterial Activity of α-Aminophosphonate Derivatives Obtained via Lipase-Catalyzed Kabachnik−Fields Reaction. Materials 2022, 15, 3846. [Google Scholar] [CrossRef]
- Sahrawat, P.; Kowalczyk, P.; Koszelewski, D.; Szymczak, M.; Kramkowski, K.; Wypych, A.; Ostaszewski, R. Influence of Open Chain and Cyclic Structure of Peptidomimetics on Antibacterial Activity in E. coli Strains. Molecules 2022, 27, 3633. [Google Scholar] [CrossRef]
- Koszelewski, D.; Kowalczyk, P.; Samsonowicz-Górski, J.; Hrunyk, A.; Brodzka, A.; Łęcka, J.; Kramkowski, K.; Ostaszewski, R. Synthesis and Antimicrobial Activity of the Pathogenic E. coli Strains of p-Quinols: Additive Effects of Copper-Catalyzed Addition of Aryl Boronic Acid to Benzoquinones. Int. J. Mol. Sci. 2023, 24, 1623. [Google Scholar] [CrossRef]
- Villafranca, J.J.A.; Guindo, M.N.; Sanz, E.; Santamaria, M.M.; Siles, M.G.; Abilés, J. Effects of cyclic parenteral nutrition on parenteral-associated liver dysfunction parameters. Nutr. J. 2017, 16, 66. [Google Scholar] [CrossRef] [PubMed]
- Smolina, N.; Bruton, J.; Kostareva, A.; Sejersen, T. Assaying Mitochondrial Respiration as an Indicator of Cellular Metabolism and Fitness. Cell Viability Assays Methods Protoc. 2017, 1601, 79–87. [Google Scholar] [CrossRef]
- Halliwell, B.; Chirico, S. Lipid peroxidation: Its mechanism, measurement, and significance. Am. J. Clin. Nutr. 1993, 57, 715S–725S; discussion 724S–725S. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.H.; Kohli, R.; Gores, G.J. Mechanisms of lipotoxicity in NAFLD and clinical implications. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 131–140. [Google Scholar] [CrossRef]
- McGarry, J.D.; Foster, D.W. Regulation of hepatic fatty acid oxidation and ketone body production. Annu. Rev. Biochem. 1980, 49, 395–420. [Google Scholar] [CrossRef] [PubMed]
- Chahal, N.; Manlhiot, C.; Wong, H.; McCrindle, B.W. Effectiveness of Omega-3 Polysaturated Fatty Acids (Fish Oil) Supplementation for Treating Hypertriglyceridemia in Children and Adolescents. Clin. Pediatr. 2014, 53, 645–651. [Google Scholar] [CrossRef]
- Federation of American Societies for Experimental Biology. Anti-Inflammatory Effects of Omega 3 Fatty Acid in Fish Oil Linked to Lowering of Prostaglandin; Science Daily: Rockville, MD, USA, 2006. [Google Scholar]
- Grau-Carmona, T.; Bonet-Saris, A.; García-de-Lorenzo, A.; Sánchez-Alvarez, C.; Rodríguez-Pozo, A.; Acosta-Escribano, J.; Miñambres, E.; Herrero-Meseguer, J.I.; Mesejo, A. Influence of n-3 polyunsaturated fatty acids enriched lipid emulsions on nosocomial infections and clinical outcomes in critically ill patients: ICU lipids study. Crit. Care Med. 2015, 43, 31–39. [Google Scholar] [CrossRef]
- Llop-Talaveron, J.; Badia-Tahull, M.B.; Lozano-Andreu, T.; Suarez-Lledo, A.; Leiva-Badosa, E. Risk factors of hepatic function alterations in hospitalized adult patients treated with short-term parenteral nutrition receiving the same lipid composition at the same dose. Lipids Health Dis. 2018, 17, 267. [Google Scholar] [CrossRef]
- Kumpf, V.J. Parenteral nutrition-associated liver disease in adult and pediatric patients. Nutr. Clin. Pract. 2006, 21, 279–290. [Google Scholar] [CrossRef]
- Piper, S.N.; Schade, I.; Beschmann, R.B.; Maleck, W.H.; Boldt, J.; Röhm, K.D. Hepatocellular integrity after parenteral nutrition: Comparison of a fish-oil-containing lipid emulsion with an olive-soybean oil-based lipid emulsion. Eur. J. Anaesthesiol. 2009, 26, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Delgado Roche, L. Oxidative stress: The dark side of soybean-oil-based emulsions used in parenteral nutrition. Oxid. Antioxid. Med. Sci. 2012, 1, 11–14. [Google Scholar] [CrossRef]
- Mitra, A.; Ahn, J. Liver Disease in Patients on Total Parenteral Nutrition. Clin. Liver Dis. 2017, 21, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Rolim, A.E.H.; Henrique-Araújo, R.; Ferraz, E.G.; Dultra, F.K.D.A.A.; Fernandez, L.G. Lipidomics in the study of lipid metabolism: Current perspectives in the omic sciences. Gene 2015, 554, 131–139. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Mohamed, I.; Nuyt, A.M.; Elremaly, W.; Rouleau, T. Impact of SMOFLipid on Pulmonary Alveolar Development in Newborn Guinea Pigs. J. Parenter. Enteral. Nutr. 2018, 42, 1314–1321. [Google Scholar] [CrossRef]
- Divakaruni, A.S.; Paradyse, A.; Ferrick, D.A.; Murphy, A.N.; Jastroch, M. Analysis and interpretation of microplate-based oxygen consumption and pH data. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2014; Volume 547, pp. 309–354. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Madej, A.; Szymczak, M.; Ostaszewski, R. α-Amidoamids as New Replacements of Antibiotics—Research on the Chosen K12, R2–R4 E. coli Strains. Materials 2020, 13, 5169. [Google Scholar] [CrossRef]
- Langie, S.A.; Kowalczyk, P.; Tomaszewski, B.; Vasilaki, A.; Maas, L.M.; Moonen, E.J.; Palagani, A.; Godschalk, R.W.; Tudek, B.; van Schooten, F.J.; et al. Redox and epigenetic regulation of the APE1 gene in the hippocampus of piglets: The effect of early life exposures. DNA Repair 2014, 18, 52–62. [Google Scholar] [CrossRef]
- Langie, S.A.; Kowalczyk, P.; Tudek, B.; Zabielski, R.; Dziaman, T.; Oliński, R.; van Schooten, F.J.; Godschalk, R.W. The effect of oxidative stress on nucleotide-excision repair in colon tissue of newborn piglets. Mutat. Res. Toxicol. Environ. Mutagen. 2010, 695, 75–80. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Jaworek, J.; Kot, M.; Sokolowska, B.; Bielen, A.; Janowska, B.; Ciesla, J.M.; Szparecki, G.; Sados, B.; Tudek, B. Inflammation increases oxidative DNA damage repair and stimulates preneoplastic changes in colons of newborn rats. J. Physiol. Pharmacol. 2016, 67, 277–286. [Google Scholar]
- Pessayre, D.; Berson, A.; Fromenty, B.; Mansouri, A. Mitochondria in Steatohepatitis. Semin. Liver Dis. 2001, 21, 57–70. [Google Scholar] [CrossRef]
- YoYounossi, Z.M.; Diehl, A.M.; Ong, J.P. Nonalcoholic fatty liver disease: An agenda for clinical research. Hepatology 2002, 35, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The role of oxidative stress during inflammatory processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Sjöberg, F.; Singer, M. The medical use of oxygen: A time for critical reappraisal. J. Intern. Med. 2013, 274, 505–528. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Young, P.J.; Laffey, J.G.; Asfar, P.; Taccone, F.S.; Skrifvars, M.B.; Meyhoff, C.S.; Radermacher, P. Dangers of hyperoxia. Crit. Care 2021, 25, 440. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.G.; Herrera, E.A.; Carrasco-Pozo, C.; Sotomayor-Zárate, R.; Cruz, G.; Morales, P.; Castillo, R.L. Pharmacological models and approaches for pathophysiological conditions associated with hypoxia and oxidative stress. Pharmacol. Ther. 2016, 158, 1–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | Female (n = 44) | Male (n = 42) | tdf=84 | p-Value * | ||||
|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |||
| Age | 56.67 | 17.42 | 57.48 | 15.30 | 55.83 | 19.56 | 0.435 | 0.664 |
| TPN [years] | 5.69 | 6.75 | 5.25 | 6.64 | 6.14 | 6.92 | −0.611 | 0.543 |
| Total | Female (n = 44) | Male (n = 42) | tdf=84 | p-Value * | ||||
|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |||
| Lipid emulsion/week (mL) | 766.00 | 340.00 | 697.95 | 349.48 | 838.10 | 318.11 | −1.942 | 0.056 |
| Lipid/week (g) | 151.00 | 61.92 | 136.86 | 57.56 | 166.79 | 63.28 | −2.296 | 0.024 |
| Control Group | TPN Group | ||||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | t * | df | P | |
| AST [U/L] | 24.24 | 15.87 | 48.44 | 34.78 | 5.423 | 170 | 0 |
| ALT [U/L] | 26.6 | 13.3 | 30.26 | 20.06 | 2.18 | 170 | 0.031 |
| CRP [mg/L] | 4.24 | 4.98 | 10.23 | 15.18 | 3.641 | 170 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dąbrowska, K.; Zaczek, Z.; Panczyk, M.; Osowska, S.; Kowalczyk, P.; Kramkowski, K.; Sobocki, J. Molecular Oxygen Levels and Percentages of DNA Damage in TPN Patients. Nutrients 2023, 15, 2206. https://doi.org/10.3390/nu15092206
Dąbrowska K, Zaczek Z, Panczyk M, Osowska S, Kowalczyk P, Kramkowski K, Sobocki J. Molecular Oxygen Levels and Percentages of DNA Damage in TPN Patients. Nutrients. 2023; 15(9):2206. https://doi.org/10.3390/nu15092206
Chicago/Turabian StyleDąbrowska, Karolina, Zuzanna Zaczek, Mariusz Panczyk, Sylwia Osowska, Paweł Kowalczyk, Karol Kramkowski, and Jacek Sobocki. 2023. "Molecular Oxygen Levels and Percentages of DNA Damage in TPN Patients" Nutrients 15, no. 9: 2206. https://doi.org/10.3390/nu15092206