
The Association of the Hypothalamic-Pituitary-Adrenal Axis with Appetite Regulation in Children with Fetal Alcohol Spectrum Disorders (FASDs)


Abstract
:1. Introduction
2. Materials and Methods
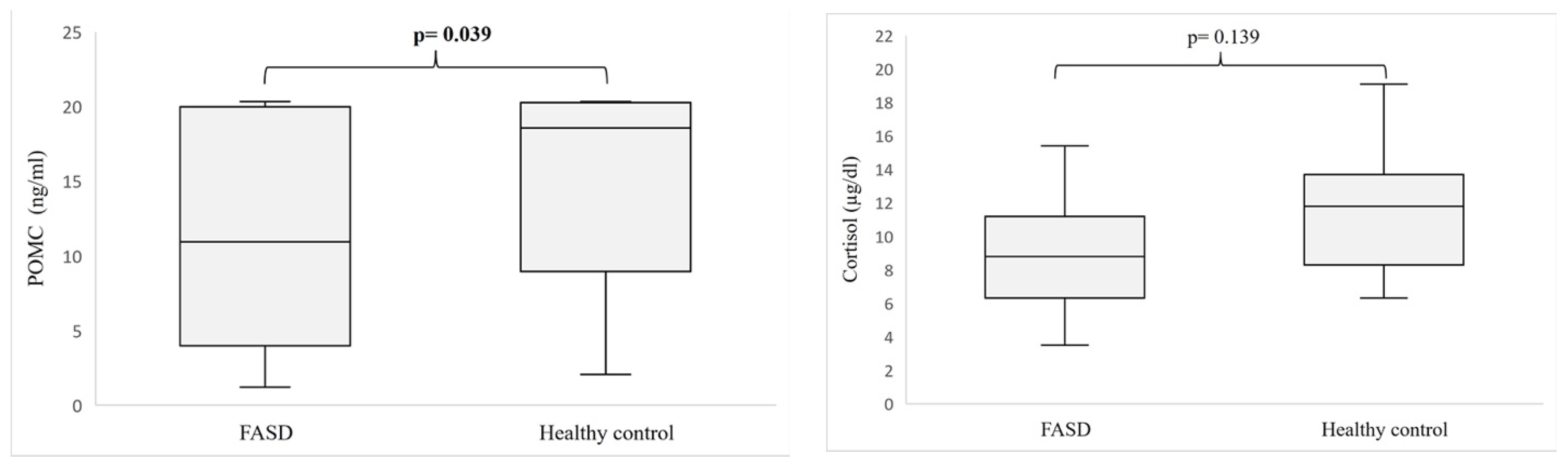
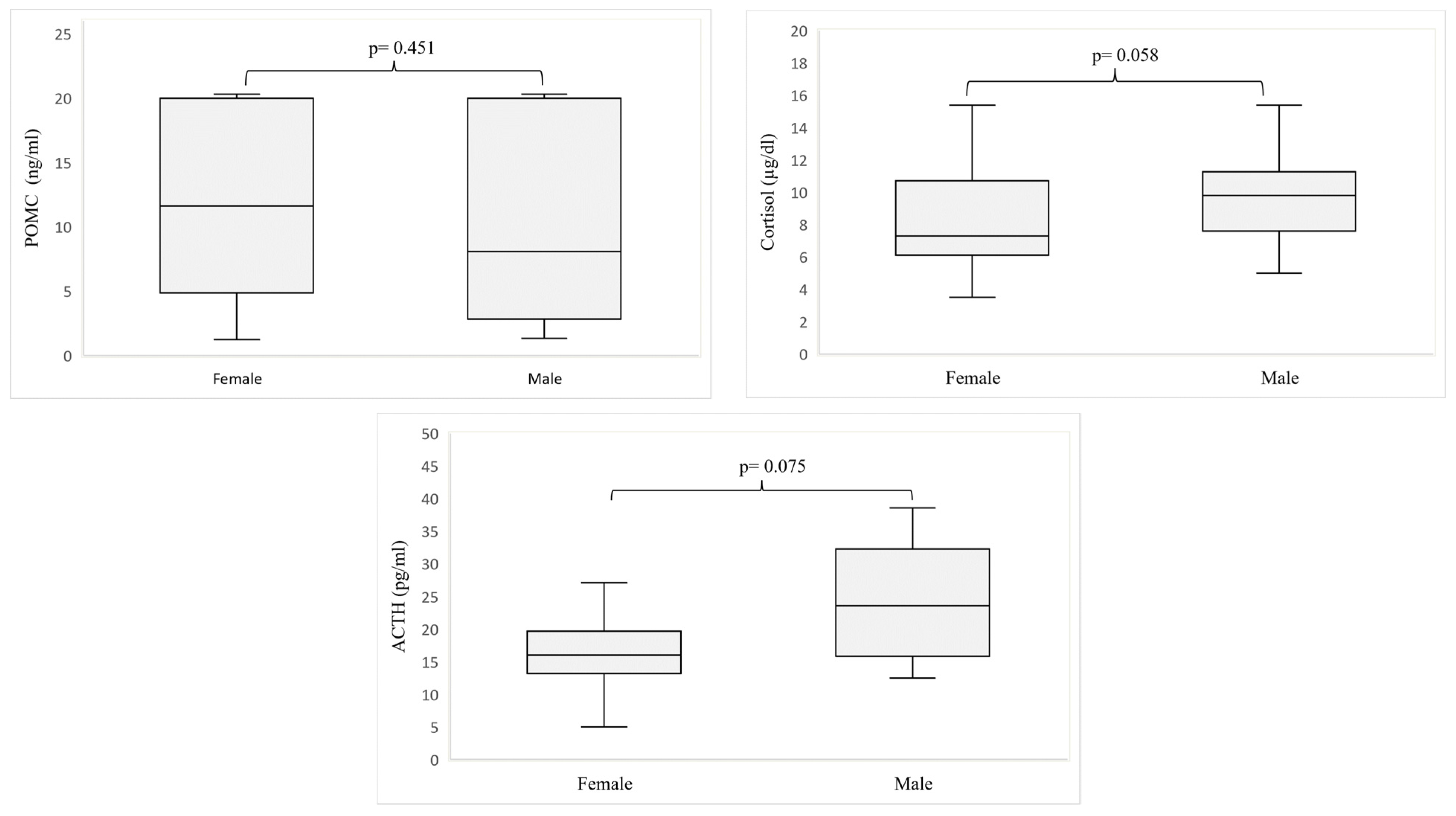
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lange, S.; Probst, C.; Gmel, G.; Rehm, J.; Burd, L.; Popova, S. Global Prevalence of Fetal Alcohol Spectrum Disorder Among Children and Youth: A Systematic Review and Meta-Analysis. JAMA Pediatr. 2017, 171, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.L.; Smith, D.W.; Ulleland, C.N.; Streissguth, P. Pattern of Malformation in Offspring of Chronic Alcoholic Mothers. Lancet 1973, 1, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- O’Keeffe, L.M.; Kearney, P.M.; McCarthy, F.P.; Khashan, A.S.; Greene, R.A.; North, R.A.; Poston, L.; McCowan, L.M.E.; Baker, P.N.; Dekker, G.A.; et al. Prevalence and Predictors of Alcohol Use during Pregnancy: Findings from International Multicentre Cohort Studies. BMJ Open 2015, 5, e006323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, S.; Lange, S.; Probst, C.; Gmel, G.; Rehm, J. Estimation of National, Regional, and Global Prevalence of Alcohol Use during Pregnancy and Fetal Alcohol Syndrome: A Systematic Review and Meta-Analysis. Lancet Glob. Health 2017, 5, e290–e299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chasnoff, I.J.; Wells, A.M.; King, L. Misdiagnosis and Missed Diagnoses in Foster and Adopted Children with Prenatal Alcohol Exposure. Pediatrics 2015, 135, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Hoyme, H.E.; Kalberg, W.O.; Elliott, A.J.; Blankenship, J.; Buckley, D.; Marais, A.-S.; Manning, M.A.; Robinson, L.K.; Adam, M.P.; Abdul-Rahman, O.; et al. Updated Clinical Guidelines for Diagnosing Fetal Alcohol Spectrum Disorders. Pediatrics 2016, 138, e20154256. [Google Scholar] [CrossRef] [Green Version]
- Astley, S.J. Validation of THE fetal alcohol spectrum Disorder (FASD) 4-digit diagnostic code. J. Popul. Ther. Clin. Pharmacol. 2013, 20, e416–e467. [Google Scholar]
- Cook, J.L.; Green, C.R.; Lilley, C.M.; Anderson, S.M.; Baldwin, M.E.; Chudley, A.E.; Conry, J.L.; LeBlanc, N.; Loock, C.A.; Lutke, J.; et al. Fetal Alcohol Spectrum Disorder: A Guideline for Diagnosis across the Lifespan. CMAJ 2016, 188, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, J.; Floyd, R.L.; Weber, M.K. Guidelines for Identifying and Referring Persons with Fetal Alcohol Syndrome. Morb. Mortal. Wkly. Rep. Recomm. Rep. 2005, 54, 1-CE-4. [Google Scholar]
- Werts, R.L.; Van Calcar, S.C.; Wargowski, D.S.; Smith, S.M. Inappropriate Feeding Behaviors and Dietary Intakes in Children with Fetal Alcohol Spectrum Disorder or Probable Prenatal Alcohol Exposure. Alcohol Clin. Exp. Res. 2014, 38, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Amos-Kroohs, R.M.; Fink, B.A.; Smith, C.J.; Chin, L.; Van Calcar, S.C.; Wozniak, J.R.; Smith, S.M. Abnormal Eating Behaviors Are Common in Children with Fetal Alcohol Spectrum Disorder. J. Pediatr. 2016, 169, 194–200.e1. [Google Scholar] [CrossRef] [Green Version]
- Sampson, P.D.; Bookstein, F.L.; Barr, H.M.; Streissguth, A.P. Prenatal Alcohol Exposure, Birthweight, and Measures of Child Size from Birth to Age 14 Years. Am. J. Public Health 1994, 84, 1421–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, R.C.; Jacobson, J.L.; Sokol, R.J.; Avison, M.J.; Jacobson, S.W. Fetal Alcohol-Related Growth Restriction from Birth through Young Adulthood and Moderating Effects of Maternal Prepregnancy Weight. Alcohol Clin. Exp. Res. 2013, 37, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Day, N.L.; Leech, S.L.; Richardson, G.A.; Cornelius, M.D.; Robles, N.; Larkby, C. Prenatal Alcohol Exposure Predicts Continued Deficits in Offspring Size at 14 Years of Age. Alcohol. Clin. Exp. Res. 2002, 26, 1584–1591. [Google Scholar] [CrossRef] [PubMed]
- Fuglestad, A.J.; Boys, C.J.; Chang, P.-N.; Miller, B.S.; Eckerle, J.K.; Deling, L.; Fink, B.A.; Hoecker, H.L.; Hickey, M.K.; Jimenez-Vega, J.M.; et al. Overweight and Obesity Among Children and Adolescents with Fetal Alcohol Spectrum Disorders. Alcohol. Clin. Exp. Res. 2014, 38, 2502–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druce, M. The Regulation of Appetite. Arch. Dis. Child. 2005, 91, 183–187. [Google Scholar] [CrossRef]
- Biebermann, H.; Kühnen, P.; Kleinau, G.; Krude, H. The Neuroendocrine Circuitry Controlled by POMC, MSH, and AGRP. In Appetite Control; Joost, H.-G., Ed.; Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 47–75. ISBN 978-3-642-24716-3. [Google Scholar]
- Wynne, K.; Stanley, S.; McGowan, B.; Bloom, S. Appetite Control. J. Endocrinol. 2005, 184, 291–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, E.K.; Quinn, M.E.; Tavernier, R.; McQuillan, M.T.; Dahlke, K.A.; Gilbert, K.E. Diurnal Cortisol Slopes and Mental and Physical Health Outcomes: A Systematic Review and Meta-Analysis. Psychoneuroendocrinology 2017, 83, 25–41. [Google Scholar] [CrossRef]
- Okulicz-Kozaryn, K.; Maryniak, A.; Borkowska, M.; Śmigiel, R.; Dylag, K.A. Diagnosis of Fetal Alcohol Spectrum Disorders (FASDs): Guidelines of Interdisciplinary Group of Polish Professionals. Int. J. Environ. Res. Public. Health 2021, 18, 7526. [Google Scholar] [CrossRef]
- Wardlaw, S.L. Hypothalamic Proopiomelanocortin Processing and the Regulation of Energy Balance. Eur. J. Pharmacol. 2011, 660, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Kim, A.; Chua, S.C.; Obici, S.; Wardlaw, S.L. Transgenic MSH Overexpression Attenuates the Metabolic Effects of a High-Fat Diet. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E121–E131. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, S.L. Clinical Review 127: Obesity as a Neuroendocrine Disease: Lessons to Be Learned from Proopiomelanocortin and Melanocortin Receptor Mutations in Mice and Men. J. Clin. Endocrinol. Metab. 2001, 86, 1442–1446. [Google Scholar] [CrossRef]
- Zhan, C.; Zhou, J.; Feng, Q.; Zhang, J.; Lin, S.; Bao, J.; Wu, P.; Luo, M. Acute and Long-Term Suppression of Feeding Behavior by POMC Neurons in the Brainstem and Hypothalamus, Respectively. J. Neurosci. 2013, 33, 3624–3632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, A.A. The Melanocortin System and Energy Balance. Peptides 2006, 27, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaswen, L.; Diehl, N.; Brennan, M.B.; Hochgeschwender, U. Obesity in the Mouse Model of Pro-Opiomelanocortin Deficiency Responds to Peripheral Melanocortin. Nat. Med. 1999, 5, 1066–1070. [Google Scholar] [CrossRef] [PubMed]
- Coll, A.P.; Farooqi, I.S.; Challis, B.G.; Yeo, G.S.H.; O’Rahilly, S. Proopiomelanocortin and Energy Balance: Insights from Human and Murine Genetics. J. Clin. Endocrinol. Metab. 2004, 89, 2557–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Cabrera, M.A.; Boyadjieva, N.I.; Berger, G.; Rousseau, B.; Sarkar, D.K. Alcohol Increases Exosome Release from Microglia to Promote Complement C1q-Induced Cellular Death of Proopiomelanocortin Neurons in the Hypothalamus in a Rat Model of Fetal Alcohol Spectrum Disorders. J. Neurosci. 2020, 40, 7965. [Google Scholar] [CrossRef]
- Bekdash, R.; Zhang, C.; Sarkar, D. Fetal Alcohol Programming of Hypothalamic Proopiomelanocortin System by Epigenetic Mechanisms and Later Life Vulnerability to Stress. Alcohol. Clin. Exp. Res. 2014, 38, 2323–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govorko, D.; Bekdash, R.A.; Zhang, C.; Sarkar, D.K. Male Germline Transmits Fetal Alcohol Adverse Effect on Hypothalamic Proopiomelanocortin Gene across Generations. Biol. Psychiatry 2012, 72, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Galiniak, S.; Podgórski, R.; Rachel, M.; Mazur, A. Serum Kisspeptin and Proopiomelanocortin in Cystic Fibrosis: A Single Study. Sci. Rep. 2022, 12, 17669. [Google Scholar] [CrossRef]
- Escelsior, A.; Cogorno, L.; Sukkar, S.G.; Amerio, A.; Donini, L.M.; Bellomo, M.; Iervasi, E.; Amore, M.; Saverino, D. Anti-Hypothalamus Autoantibodies in Anorexia Nervosa: A Possible New Mechanism in Neuro-Physiological Derangement? Eat. Weight Disord.-Stud. Anorex. Bulim. Obes. 2022, 27, 2481–2496. [Google Scholar] [CrossRef] [PubMed]
- Page-Wilson, G.; Freda, P.U.; Jacobs, T.P.; Khandji, A.G.; Bruce, J.N.; Foo, S.T.; Meece, K.; White, A.; Wardlaw, S.L. Clinical Utility of Plasma POMC and AgRP Measurements in the Differential Diagnosis of ACTH-Dependent Cushing’s Syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E1838–E1845. [Google Scholar] [CrossRef]
- Page-Wilson, G.; Meece, K.; White, A.; Rosenbaum, M.; Leibel, R.L.; Smiley, R.; Wardlaw, S.L. Proopiomelanocortin, Agouti-Related Protein, and Leptin in Human Cerebrospinal Fluid: Correlations with Body Weight and Adiposity. Am. J. Physiol.-Endocrinol. Metab. 2015, 309, E458–E465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, D.K.; Kuhn, P.; Marano, J.; Chen, C.; Boyadjieva, N. Alcohol Exposure during the Developmental Period Induces Beta-Endorphin Neuronal Death and Causes Alteration in the Opioid Control of Stress Axis Function. Endocrinology 2007, 148, 2828–2834. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, L.; Fish, E.W.; O’Leary-Moore, S.K.; Parnell, S.E.; Sulik, K.K. Hypothalamic-Pituitary-Adrenal Axis and Behavioral Dysfunction Following Early Binge-like Prenatal Alcohol Exposure in Mice. Alcohol Fayettev. N 2015, 49, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Gangisetty, O.; Bekdash, R.; Maglakelidze, G.; Sarkar, D.K. Fetal Alcohol Exposure Alters Proopiomelanocortin Gene Expression and Hypothalamic-Pituitary-Adrenal Axis Function via Increasing MeCP2 Expression in the Hypothalamus. PLoS ONE 2014, 9, e113228. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, A.; Priftis, K.N. Regulation of the Hypothalamic-Pituitary-Adrenal Axis. Neuroimmunomodulation 2009, 16, 265–271. [Google Scholar] [CrossRef]
- de Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the Brain: From Adaptation to Disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Podgórski, R.P.; Aebisher, D.; Stompor, M.; Podgórska, D.; Mazur, A. Congenital Adrenal Hyperplasia: Clinical Symptoms and Diagnostic Methods. Acta Biochim. Pol. 2018, 65, 25–33. [Google Scholar] [CrossRef]
- Roy, M.P.; Kirschbaum, C.; Steptoe, A. Psychological, Cardiovascular, and Metabolic Correlates of Individual Differences in Cortisol Stress Recovery in Young Men. Psychoneuroendocrinology 2001, 26, 375–391. [Google Scholar] [CrossRef]
- Riad, M.; Mogos, M.; Thangathurai, D.; Lumb, P.D. Steroids. Curr. Opin. Crit. Care 2002, 8, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Jirikowic, T.; Chen, M.; Nash, J.; Gendler, B.; Carmichael Olson, H. Regulatory Behaviors and Stress Reactivity among Infants at High Risk for Fetal Alcohol Spectrum Disorders: An Exploratory Study. J. Ment. Health Res. Intellect. Disabil. 2016, 9, 171–188. [Google Scholar] [CrossRef]
- Keiver, K.; Bertram, C.P.; Orr, A.P.; Clarren, S. Salivary Cortisol Levels Are Elevated in the Afternoon and at Bedtime in Children with Prenatal Alcohol Exposure. Alcohol 2015, 49, 79–87. [Google Scholar] [CrossRef]
- Ramsay, D.S.; Bendersky, M.I.; Lewis, M. Effect of Prenatal Alcohol and Cigarette Exposure on Two- and Six-Month-Old Infants’ Adrenocortical Reactivity to Stress1. J. Pediatr. Psychol. 1996, 21, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Hanlon-Dearman, A.; Chen, M.L.; Olson, H.C. Understanding and Managing Sleep Disruption in Children with Fetal Alcohol Spectrum Disorder. Biochem. Cell Biol. 2018, 96, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Schuetze, P.; Lopez, F.A.; Granger, D.A.; Eiden, R.D. The Association between Prenatal Exposure to Cigarettes and Cortisol Reactivity and Regulation in 7-Month-Old Infants. Dev. Psychobiol. 2008, 50, 819–834. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Flores, B.; Gomez, R.G.; Solvason, H.B.; Kenna, H.; Williams, G.H.; Schatzberg, A.F. Cortisol Circadian Rhythm Alterations in Psychotic Major Depression. Biol. Psychiatry 2006, 60, 275–281. [Google Scholar] [CrossRef]
- Tataranni, P.A.; Larson, D.E.; Snitker, S.; Young, J.B.; Flatt, J.P.; Ravussin, E. Effects of Glucocorticoids on Energy Metabolism and Food Intake in Humans. Am. J. Physiol. 1996, 271, E317–E325. [Google Scholar] [CrossRef]
- Epel, E.; Lapidus, R.; McEwen, B.; Brownell, K. Stress May Add Bite to Appetite in Women: A Laboratory Study of Stress-Induced Cortisol and Eating Behavior. Psychoneuroendocrinology 2001, 26, 37–49. [Google Scholar] [CrossRef]
- Jéquier, E. Leptin Signaling, Adiposity, and Energy Balance. Ann. N. Y. Acad. Sci. 2002, 967, 379–388. [Google Scholar] [CrossRef]
- Sinha, R.; Jastreboff, A.M. Stress as a Common Risk Factor for Obesity and Addiction. Biol. Psychiatry 2013, 73, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Newell-Price, J.; Bertagna, X.; Grossman, A.B.; Nieman, L.K. Cushing’s Syndrome. Lancet 2006, 367, 1605–1617. [Google Scholar] [CrossRef]
- Rosmond, R.; Dallman, M.F.; Björntorp, P. Stress-Related Cortisol Secretion in Men: Relationships with Abdominal Obesity and Endocrine, Metabolic and Hemodynamic Abnormalities. J. Clin. Endocrinol. Metab. 1998, 83, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Poggioli, R.; Vergoni, A.V.; Bertolini, A. ACTH-(1-24) and Alpha-MSH Antagonize Feeding Behavior Stimulated by Kappa Opiate Agonists. Peptides 1986, 7, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Rivier, C. Alcohol Stimulates ACTH Secretion in the Rat: Mechanisms of Action and Interactions with Other Stimuli. Alcohol. Clin. Exp. Res. 1996, 20, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Fehm, H.L.; Smolnik, R.; Kern, W.; McGregor, G.P.; Bickel, U.; Born, J. The Melanocortin Melanocyte-Stimulating Hormone/Adrenocorticotropin(4-10) Decreases Body Fat in Humans. J. Clin. Endocrinol. Metab. 2001, 86, 1144–1148. [Google Scholar] [CrossRef]
- Krude, H.; Biebermann, H.; Schnabel, D.; Tansek, M.Z.; Theunissen, P.; Mullis, P.E.; Grüters, A. Obesity Due to Proopiomelanocortin Deficiency: Three New Cases and Treatment Trials with Thyroid Hormone and ACTH4-10. J. Clin. Endocrinol. Metab. 2003, 88, 4633–4640. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, K.; Rasmussen, C.; Oberlander, T.F.; Loock, C.; Pei, J.; Andrew, G.; Reynolds, J.; Weinberg, J. Dysregulation of the Cortisol Diurnal Rhythm Following Prenatal Alcohol Exposure and Early Life Adversity. Alcohol 2016, 53, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Belgardt, B.F.; Okamura, T.; Brüning, J.C. Hormone and Glucose Signalling in POMC and AgRP Neurons. J. Physiol. 2009, 587, 5305–5314. [Google Scholar] [CrossRef]
- Calixto, C.; Martinez, F.E.; Jorge, S.M.; Moreira, A.C.; Martinelli, C.E. Correlation between Plasma and Salivary Cortisol Levels in Preterm Infants. J. Pediatr. 2002, 140, 116–118. [Google Scholar] [CrossRef]
- Gallo-Payet, N. 60 YEARS OF POMC: Adrenal and Extra-Adrenal Functions of ACTH. J. Mol. Endocrinol. 2016, 56, T135–T156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.M.; Clark, P. The Short Synacthen Test: Is Less Best? Clin. Endocrinol. 1999, 51, 151–152. [Google Scholar] [CrossRef]
- Knutsson, U.; Dahlgren, J.; Marcus, C.; Rosberg, S.; Brönnegård, M.; Stierna, P.; Albertsson-Wikland, K. Circadian Cortisol Rhythms in Healthy Boys and Girls: Relationship with Age, Growth, Body Composition, and Pubertal Development*. J. Clin. Endocrinol. Metab. 1997, 82, 536–540. [Google Scholar] [CrossRef]
- Dahl, R.E.; Siegel, S.F.; Williamson, D.E.; Lee, P.A.; Perel, J.; Birmaher, B.; Ryan, N.D. Corticotropin Releasing Hormone Stimulation Test and Nocturnal Cortisol Levels in Normal Children. Pediatr. Res. 1992, 32, 64–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martens, A.; Duran, B.; Vanbesien, J.; Verheyden, S.; Rutteman, B.; Staels, W.; Anckaert, E.; Gies, I.; De Schepper, J. Clinical and Biological Correlates of Morning Serum Cortisol in Children and Adolescents with Overweight and Obesity. PLoS ONE 2021, 16, e0258653. [Google Scholar] [CrossRef]
- Jonetz-Mentzel, L.; Wiedemann, G. Establishment of Reference Ranges for Cortisol in Neonates, Infants, Children and Adolescents. Eur. J. Clin. Chem. Clin. Biochem. J. Forum Eur. Clin. Chem. Soc. 1993, 31, 525–529. [Google Scholar] [CrossRef] [Green Version]
- Kiess, W.; Meidert, A.; Dressendörfer, R.A.; Schriever, K.; Kessler, U.; König, A.; Schwarz, H.P.; Strasburger, C.J. Salivary Cortisol Levels throughout Childhood and Adolescence: Relation with Age, Pubertal Stage, and Weight. Pediatr. Res. 1995, 37, 502–506. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| FASD | Healthy Controls | p-Value | ||
|---|---|---|---|---|
| Sex (F/M) | 31/31 | 7/16 | ||
| Age (years) | mean ± SD | 7.52 ± 4.16 | 7.45 ± 5.12 | 0.847 |
| range | 0.42–16.5 | 0.42–17 | ||
| BMI percentile | mean ± SD | 32.38 ± 31.24 | 60.71 ± 27.03 | 0.035 |
| range | 0.1–99.9 | 12.0–99 | ||
| Clinical laboratory markers | ||||
| Cholesterol (mg/dL) Norm < 190 | median | 150 | 155 | 0.971 |
| range | 76–244 | 126–191 | ||
| LDL (mg/dL) Norm < 135 | median | 90 | 95 | 0.827 |
| range | 31–163 | 72–104 | ||
| HDL (mg/dL) Norm > 40 | median | 53 | 53 | 0.856 |
| range | 24–108 | 42–59 | ||
| Triglycerides (mg/dL) Norm < 150 | median | 74 | 65 | 0.753 |
| range | 30–241 | 38–141 | ||
| Glucose (mg/dL) Norm (70–99) | median | 84 | 87 | 0.669 |
| range | 72–99 | 68–94 | ||
| Insulin (mIU/mL) Norm < 15 | median | 4.85 | 2.05 | 0.167 |
| range | 1.25–17 | 1.0–9.03 | ||
| HbA1c (%) Normal range (4–6) | median | 5.36 | 5.41 | 0.774 |
| range | 4.71–5.86 | 5.26–5.55 | ||
| HOMA-IR Norm < 2.5 | median | 0.96 | - | - |
| range | 0.23–3.62 | - | ||
| FAS | ND-PAE | FASD Risk | p-Value | p-Value * | ||
|---|---|---|---|---|---|---|
| Sex (F/M) | 14/12 | 15/16 | 2/3 | |||
| Age (years) | mean ± SD | 7.91 ± 4.77 | 8.13 ± 3.32 | 2.25 ± 1.26 | 0.004 | 0.843 |
| range | 0.42–16.5 | 2.08–13.5 | 1.17–4.42 | |||
| BMI percentile | mean ± SD | 22.12 ± 27.51 | 42.04 ± 33.02 | 27.33 ± 13.87 | 0.053 | 0.020 |
| range | 0.1–78 | 0.1–99.9 | 12–39 | |||
| Clinical laboratory markers | ||||||
| Cholesterol (mg/dL) Norm < 190 | median | 154.5 | 161 | 141 | 0.277 | 0.110 |
| range | 76–238 | 114–244 | 104–185 | |||
| LDL (mg/dL) Norm < 135 | median | 86 | 75 | 84 | 0.598 | 0.365 |
| range | 31–143 | 114–244 | 33–119 | |||
| HDL (mg/dL) Norm > 40 | median | 49.5 | 53 | 46 | 0.515 | 0.382 |
| range | 33–80 | 24–108 | 33–71 | |||
| Triglycerides (mg/dL) Norm < 150 | median | 64 | 75 | 75 | 0.590 | 0.607 |
| range | 30.0–229 | 34–241 | 55–99 | |||
| Glucose (mg/dL) Norm (70–99) | median | 82 | 87 | 80 | 0.111 | 0.211 |
| range | 72–99 | 74–99 | 76–91 | |||
| Insulin (mIU/mL) Norm < 15 mIU/mL | median | 5.10 | 4.22 | 3.22 | 0.596 | 0.623 |
| range | 1.41–16.46 | 1.56–13.97 | 1.25–17 | |||
| HbA1c (%) Normal range (4–6) | median | 5.24 | 5.45 | 5.35 | 0.076 | 0.039 |
| range | 4.81–5.86 | 4.89–5.85 | 4.71–5.53 | |||
| HOMA-IR Norm < 2.5 | median | 1.07 | 1.29 | 1.73 | 0.591 | 0.790 |
| range | 0.27–3.62 | 0.31–3.53 | 0.23–3.36 | |||
| Hormone | FAS | ND-PAE | FASD Risk | p-Value | p-Value * | |
|---|---|---|---|---|---|---|
| POMC (ng/mL) | median | 9.35 | 15.93 | 11.22 | 0.725 | 0.479 |
| range | 1.22–20.32 | 1.51–20.32 | 3.92–16.84 | |||
| ACTH (pg/mL) | median | 24.9 | 17.7 | 14.3 | 0.942 | 0.401 |
| range | 12.3–38.6 | 5.0–37.6 | 12.5–66.1 | |||
| Cortisol (µg/dL) | median | 9.9 | 7.95 | 9.0 | 0.649 | 0.370 |
| range | 4.30–18.9 | 3.5–26.0 | 6.1–23.2 |
| Age | BMI Percentile | Cortisol | ACTH | Cholesterol | LDL | HDL | TGL | Glucose | Insulin | HOMA-IR | HbA1c | POMC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POMC | R | 0.370 | 0.285 | 0.015 | 0.386 | 0.193 | 0.126 | 0.154 | 0.177 | 0.292 | 0.475 | 0.473 | 0.076 | |
| p | 0.003 | 0.030 | 0.912 | 0.020 | 0.137 | 0.333 | 0.236 | 0.173 | 0.026 | <0.001 | <0.001 | 0.587 | ||
| ACTH | R | 0.304 | 0.306 | 0.607 | 0.335 | 0.262 | 0.208 | 0.175 | 0.152 | 0.183 | 0.189 | 0.045 | 0.386 | |
| p | 0.071 | 0.083 | <0.001 | 0.046 | 0.141 | 0.245 | 0.33 | 0.398 | 0.307 | 0.292 | 0.805 | 0.020 | ||
| Cortisol | R | 0.071 | −0.093 | 0.607 | 0.106 | 0.122 | −0.024 | 0.188 | −0.158 | −0.089 | −0.127 | −0.116 | 0.015 | |
| p | 0.592 | 0.497 | <0.001 | 0.444 | 0.379 | 0.866 | 0.172 | 0.268 | 0.543 | 0.386 | 0.424 | 0.912 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podgórski, R.; Galiniak, S.; Mazur, A.; Domin, A. The Association of the Hypothalamic-Pituitary-Adrenal Axis with Appetite Regulation in Children with Fetal Alcohol Spectrum Disorders (FASDs). Nutrients 2023, 15, 1366. https://doi.org/10.3390/nu15061366
Podgórski R, Galiniak S, Mazur A, Domin A. The Association of the Hypothalamic-Pituitary-Adrenal Axis with Appetite Regulation in Children with Fetal Alcohol Spectrum Disorders (FASDs). Nutrients. 2023; 15(6):1366. https://doi.org/10.3390/nu15061366
Chicago/Turabian StylePodgórski, Rafał, Sabina Galiniak, Artur Mazur, and Agnieszka Domin. 2023. "The Association of the Hypothalamic-Pituitary-Adrenal Axis with Appetite Regulation in Children with Fetal Alcohol Spectrum Disorders (FASDs)" Nutrients 15, no. 6: 1366. https://doi.org/10.3390/nu15061366