
Isolation of Alginate-Degrading Bacteria from the Human Gut Microbiota and Discovery of Bacteroides xylanisolvens AY11-1 as a Novel Anti-Colitis Probiotic Bacterium
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Isolation of Alginate-Degrading Bacteria from Human Gut Microbiota
2.3. Animals and Treatment
2.4. High-Throughput Sequencing and Bioinformatic Analyses
2.5. Statistical Analyses
3. Results
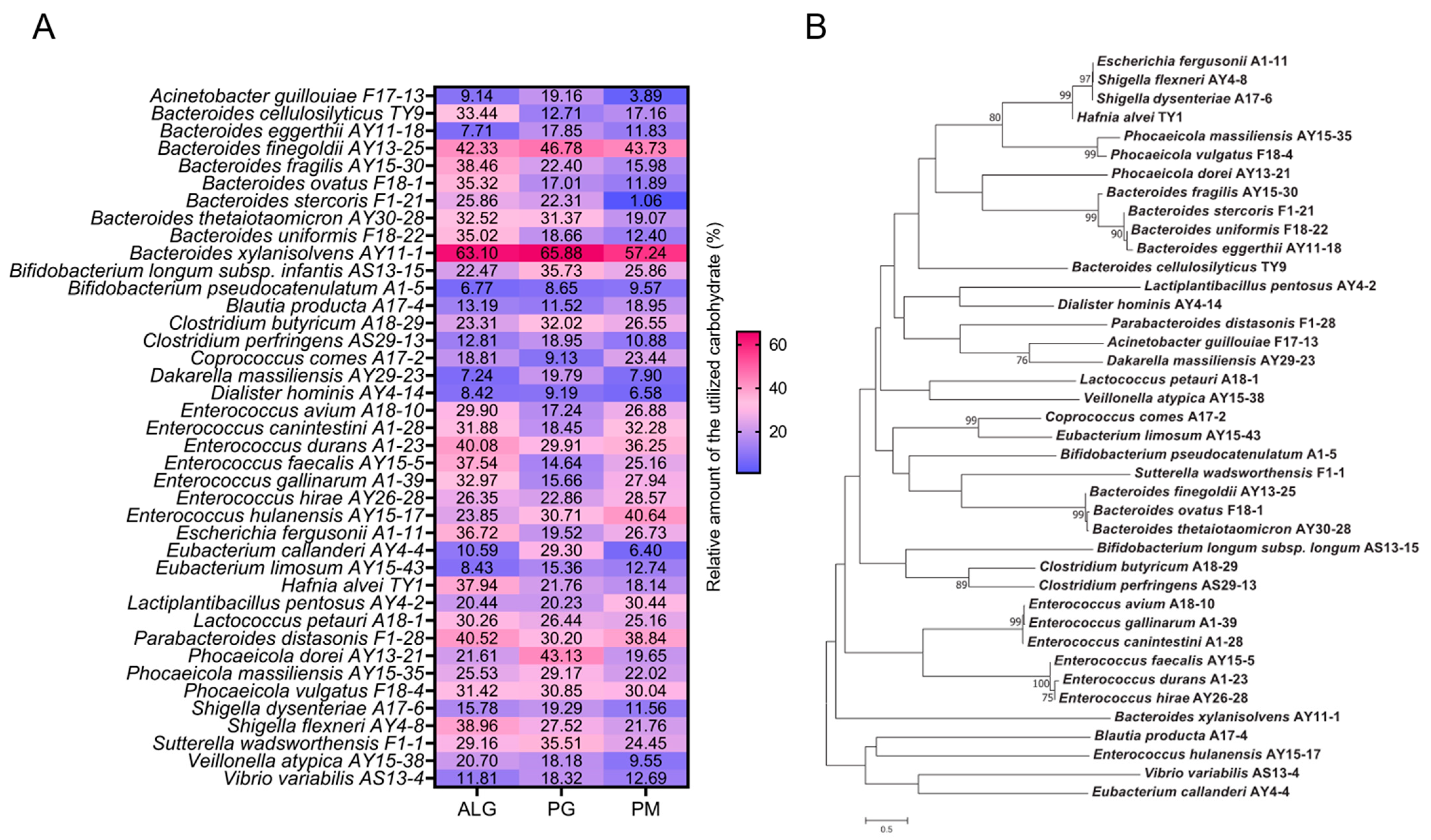
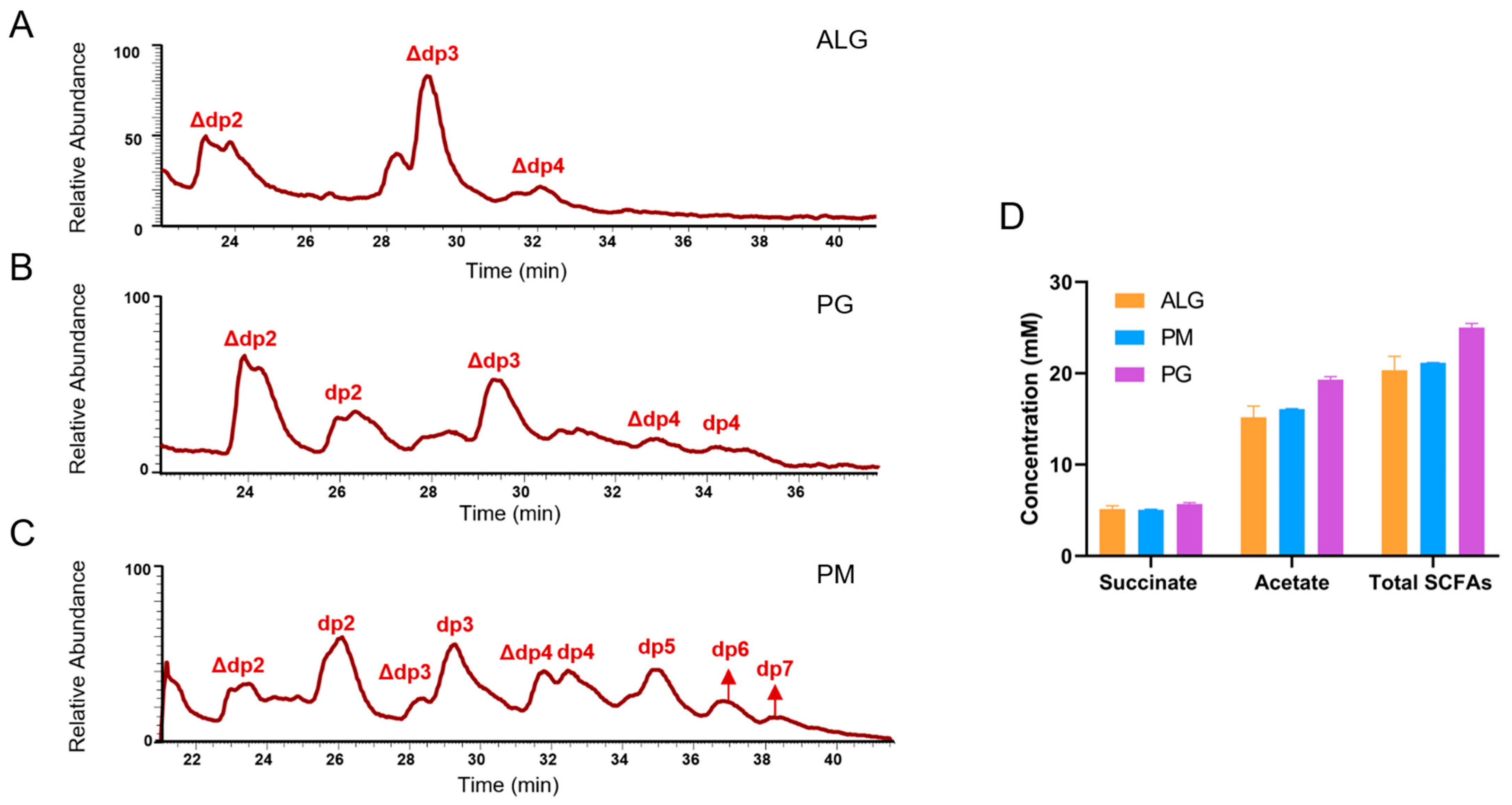
3.1. B. xylanisolvens AY11-1 Is a Potent Alginate-Degrading Bacterium in the Human Gut
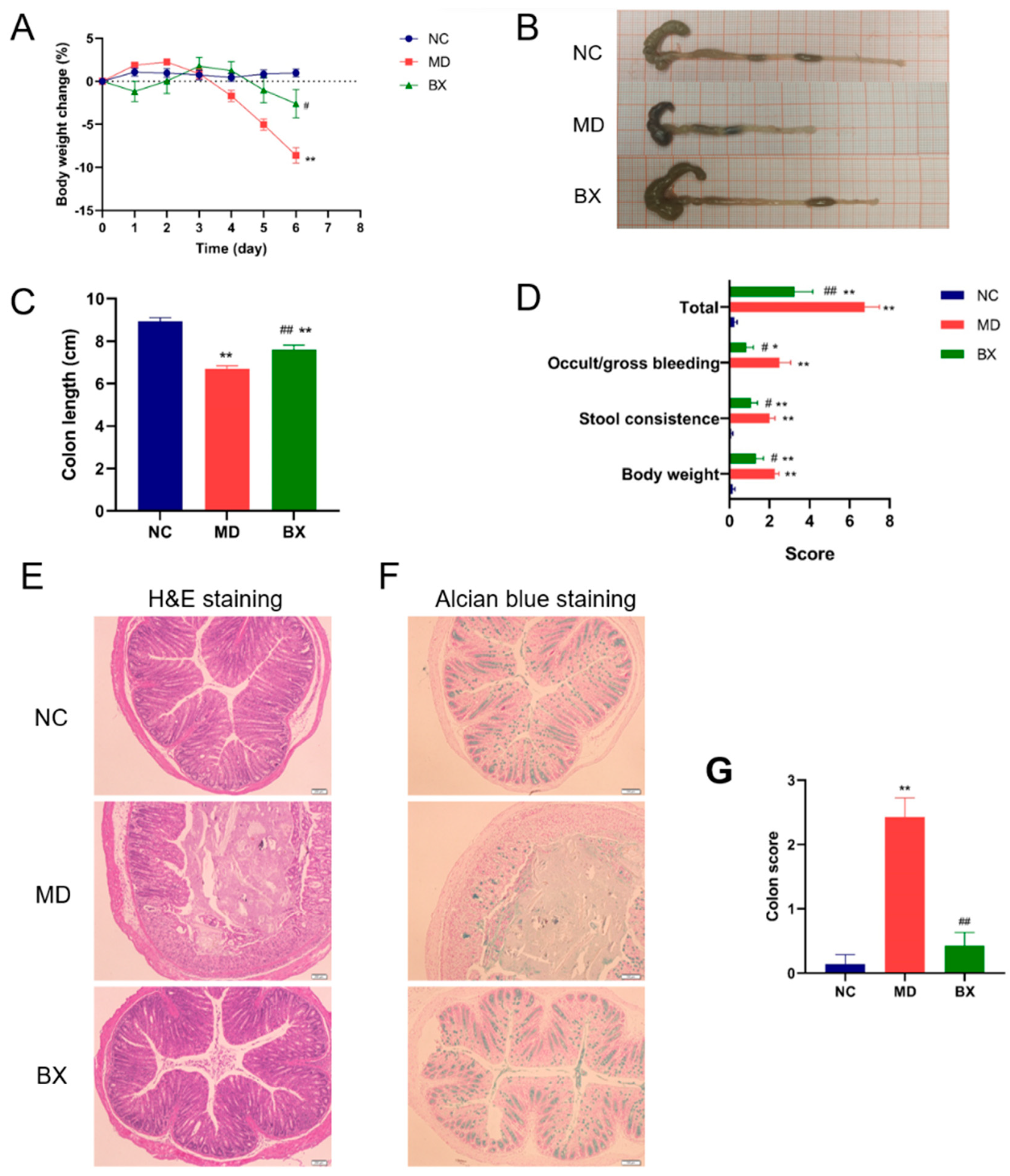
3.2. B. xylanisolvens AY11-1 Ameliorated Ulcerative Colitis in DSS-Fed Mice
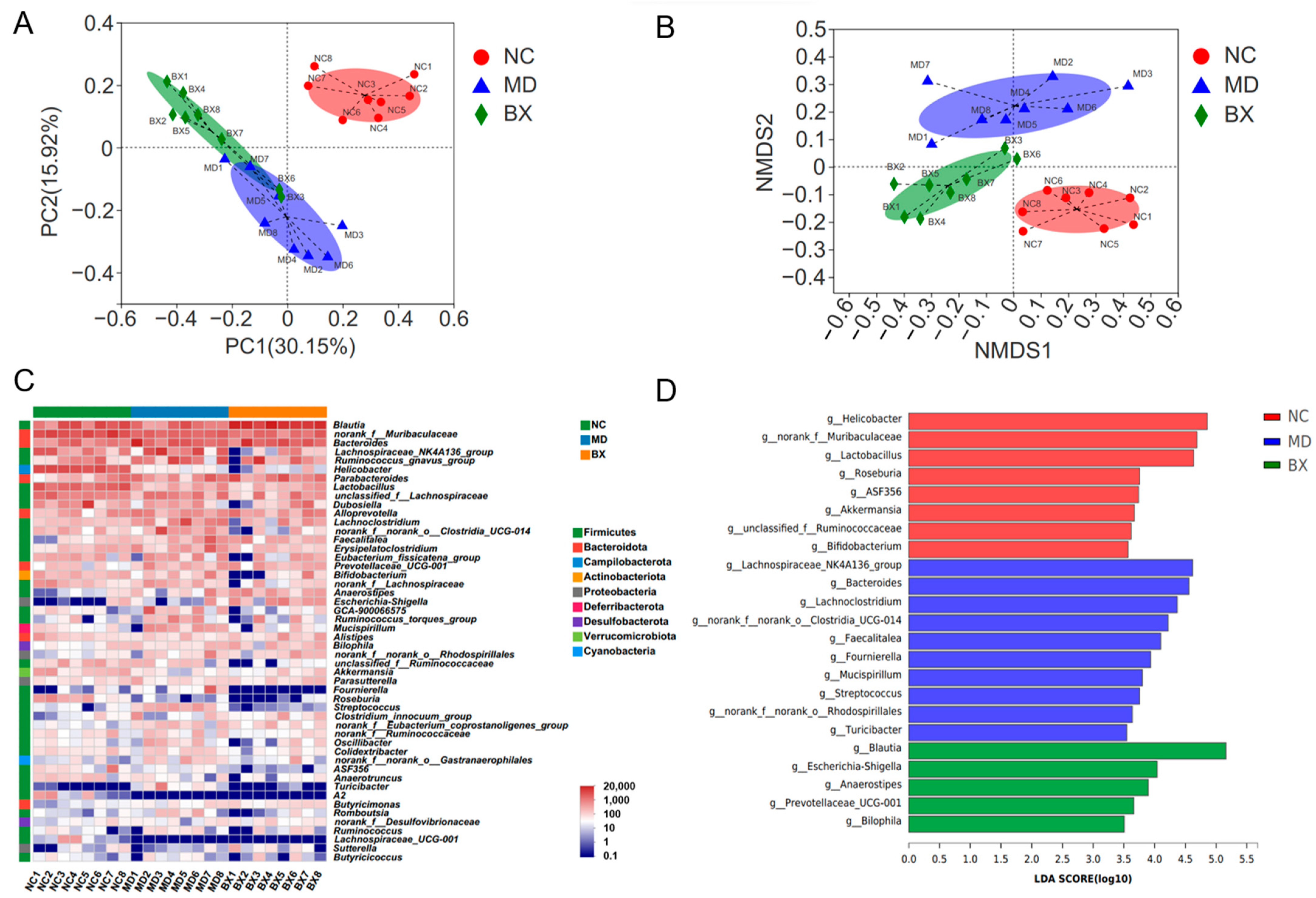
3.3. B. xylanisolvens AY11-1 Modulated the Gut Microbiome by Increasing the Abundances of Blautia spp. and Prevotellaceae UCG-001
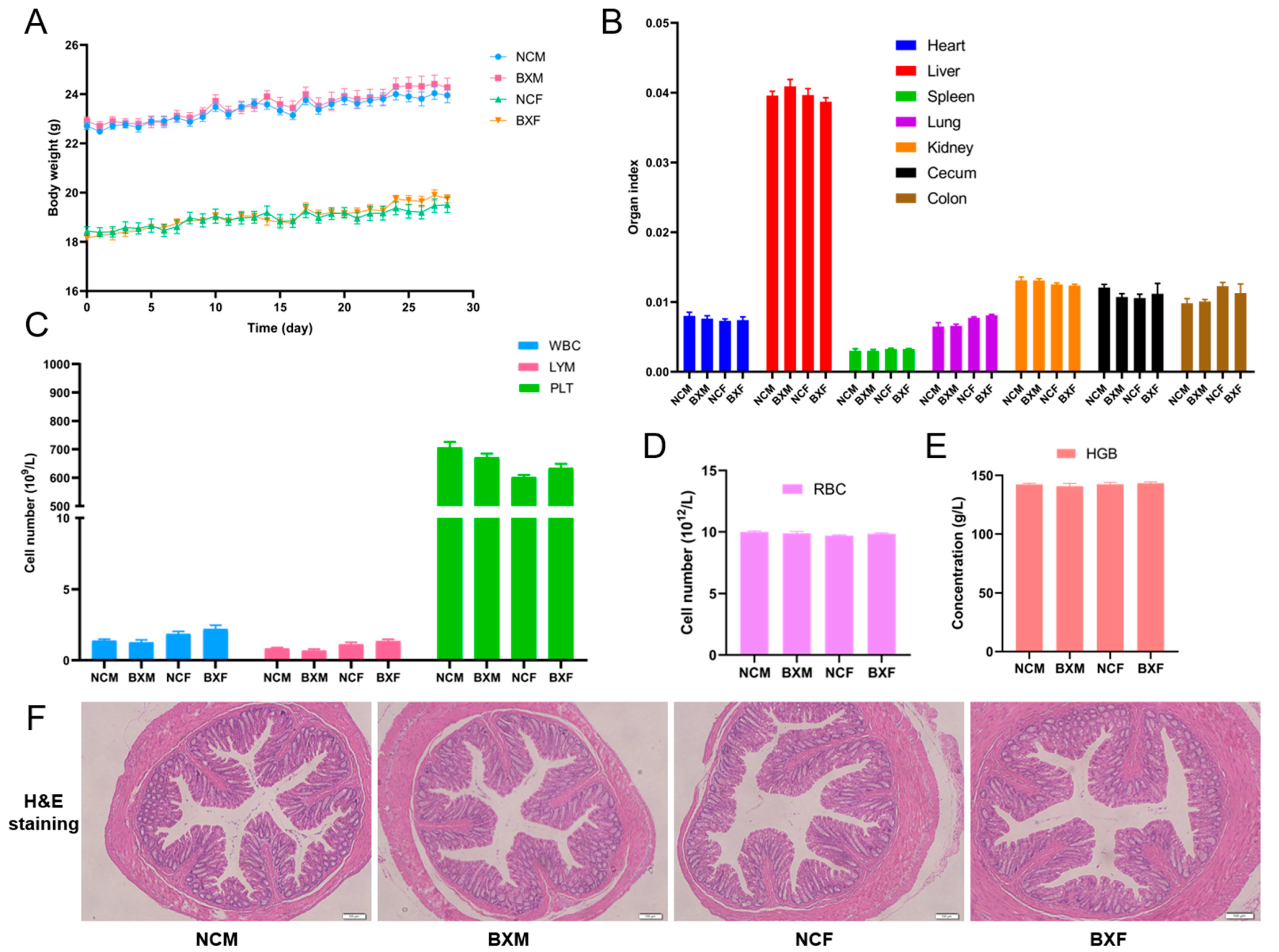
3.4. B. xylanisolvens AY11-1 Showed No Oral Toxicity and Was Well-Tolerated in Both Male and Female Mice
4. Discussion
4.1. Future Perspectives and Potential Applications of B. xylanisolvens AY11-1
4.2. Limitations of the Current Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bi, D.; Yang, X.; Yao, L.; Hu, Z.; Li, H.; Xu, X.; Lu, J. Potential food and nutraceutical applications of alginate: A Review. Mar. Drugs 2022, 20, 564. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: An overview. Carbohydr. Polym. 2018, 179, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Pan, L.; Shang, Q.; Yu, G. Fermentation of alginate and its derivatives by different enterotypes of human gut microbiota: Towards personalized nutrition using enterotype-specific dietary fibers. Int. J. Biol. Macromol. 2021, 183, 1649–1659. [Google Scholar] [CrossRef]
- Manichanh, C.; Borruel, N.; Casellas, F.; Guarner, F. The gut microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 599–608. [Google Scholar] [CrossRef]
- Ma, M.; Fu, T.; Wang, Y.; Zhang, A.; Gao, P.; Shang, Q.; Yu, G. Polysaccharide from edible alga Enteromorpha clathrata improves ulcerative colitis in association with increased abundance of Parabacteroides spp. in the gut microbiota of dextran sulfate sodium-fed mice. Mar. Drugs 2022, 20, 764. [Google Scholar] [CrossRef] [PubMed]
- Mentella, M.C.; Scaldaferri, F.; Pizzoferrato, M.; Gasbarrini, A.; Miggiano, G.A.D. Nutrition, IBD and gut microbiota: A review. Nutrients 2020, 12, 944. [Google Scholar] [CrossRef] [Green Version]
- Pi, Y.; Zhang, X.; Wu, Y.; Wang, Z.; Bai, Y.; Liu, X.; Han, D.; Zhao, J.; Tobin, I.; Zhao, J.; et al. Alginate alleviates dextran sulfate sodium-induced colitis by promoting Bifidobacterium animalis and intestinal hyodeoxycholic acid synthesis in mice. Microbiol Spectr. 2022, 10, e0297922. [Google Scholar] [CrossRef]
- Fu, T.; Zhou, L.; Fu, Z.; Zhang, B.; Li, Q.; Pan, L.; Zhou, C.; Zhao, Q.; Shang, Q.; Yu, G. Enterotype-specific effect of human gut microbiota on the fermentation of marine algae oligosaccharides: A preliminary proof-of-concept in vitro study. Polymers 2022, 14, 770. [Google Scholar] [CrossRef]
- Pan, L.; Ai, X.; Fu, T.; Ren, L.; Shang, Q.; Li, G.; Yu, G. In vitro fermentation of hyaluronan by human gut microbiota: Changes in microbiota community and potential degradation mechanism. Carbohydr. Polym. 2021, 269, 118313. [Google Scholar] [CrossRef]
- Pan, L.; Fu, T.; Cheng, H.; Mi, J.; Shang, Q.; Yu, G. Polysaccharide from edible alga Gloiopeltis furcata attenuates intestinal mucosal damage by therapeutically remodeling the interactions between gut microbiota and mucin O-glycans. Carbohydr. Polym. 2022, 278, 118921. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Won, S. Evaluation of 16S rRNA Databases for Taxonomic Assignments Using Mock Community. Genom. Inform. 2018, 16, e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Q.; Yin, Y.; Zhu, L.; Li, G.; Yu, G.; Wang, X. Degradation of chondroitin sulfate by the gut microbiota of Chinese individuals. Int. J. Biol. Macromol. 2016, 86, 112–118. [Google Scholar] [CrossRef]
- Mirshafiey, A.; Khodadadi, A.; Rehm, B.H.; Khorramizadeh, M.R.; Eslami, M.B.; Razavi, A.; Saadat, F. Sodium alginate as a novel therapeutic option in experimental colitis. Scand. J. Immunol. 2005, 61, 316–321. [Google Scholar] [CrossRef]
- Niu, W.; Chen, Y.; Wang, L.; Li, J.; Cui, Z.; Lv, J.; Yang, F.; Huo, J.; Zhang, Z.; Ju, J. The combination of sodium alginate and chlorogenic acid enhances the therapeutic effect on ulcerative colitis by the regulation of inflammation and the intestinal flora. Food Funct. 2022, 13, 10710–10723. [Google Scholar] [CrossRef]
- Razavi, A.; Khodadadi, A.; Eslami, M.B.; Eshraghi, S.; Mirshafiey, A. Therapeutic effect of sodium alginate in experimental chronic ulcerative colitis. Iran. J. Allergy Asthma Immunol. 2008, 7, 13–18. [Google Scholar]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, M.; Garner, A.; Vlamakis, H.; Xavier, R.J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 2019, 17, 497–511. [Google Scholar] [CrossRef]
- Li, M.; Li, G.; Shang, Q.; Chen, X.; Liu, W.; Pi, X.; Zhu, L.; Yin, Y.; Yu, G.; Wang, X. In vitro fermentation of alginate and its derivatives by human gut microbiota. Anaerobe 2016, 39, 19–25. [Google Scholar] [CrossRef]
- Tan, H.; Zhai, Q.; Chen, W. Investigations of Bacteroides spp. towards next-generation probiotics. Food Res. Int. 2019, 116, 637–644. [Google Scholar] [CrossRef]
- O′Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodmann, T.; Endo, A.; Gueimonde, M.; Vinderola, G.; Kneifel, W.; de Vos, W.M.; Salminen, S.; Gómez-Gallego, C. Safety of novel microbes for human consumption: Practical examples of assessment in the European Union. Front. Microbiol. 2017, 8, 1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhao, J.; Zhang, H.; Lee, Y.K.; Zhai, Q.; Chen, W. Roles of intestinal Bacteroides in human health and diseases. Crit. Rev. Food Sci. Nutr. 2021, 61, 3518–3536. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, T.; Wang, Y.; Ma, M.; Dai, W.; Pan, L.; Shang, Q.; Yu, G. Isolation of Alginate-Degrading Bacteria from the Human Gut Microbiota and Discovery of Bacteroides xylanisolvens AY11-1 as a Novel Anti-Colitis Probiotic Bacterium. Nutrients 2023, 15, 1352. https://doi.org/10.3390/nu15061352
Fu T, Wang Y, Ma M, Dai W, Pan L, Shang Q, Yu G. Isolation of Alginate-Degrading Bacteria from the Human Gut Microbiota and Discovery of Bacteroides xylanisolvens AY11-1 as a Novel Anti-Colitis Probiotic Bacterium. Nutrients. 2023; 15(6):1352. https://doi.org/10.3390/nu15061352
Chicago/Turabian StyleFu, Tianyu, Yamin Wang, Mingfeng Ma, Wei Dai, Lin Pan, Qingsen Shang, and Guangli Yu. 2023. "Isolation of Alginate-Degrading Bacteria from the Human Gut Microbiota and Discovery of Bacteroides xylanisolvens AY11-1 as a Novel Anti-Colitis Probiotic Bacterium" Nutrients 15, no. 6: 1352. https://doi.org/10.3390/nu15061352