
Molecular Targets and Mechanisms of 6,7-Dihydroxy-2,4-dimethoxyphenanthrene from Chinese Yam Modulating NF-κB/COX-2 Signaling Pathway: The Application of Molecular Docking and Gene Silencing
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Anti-Inflammatory Activity Test for PC2 and PC4
2.3. Molecular Docking
2.4. Molecular Dynamics Simulation
2.5. Drug Intervention on Caco-2 Cells and Gene Silenced Caco-2 Cells
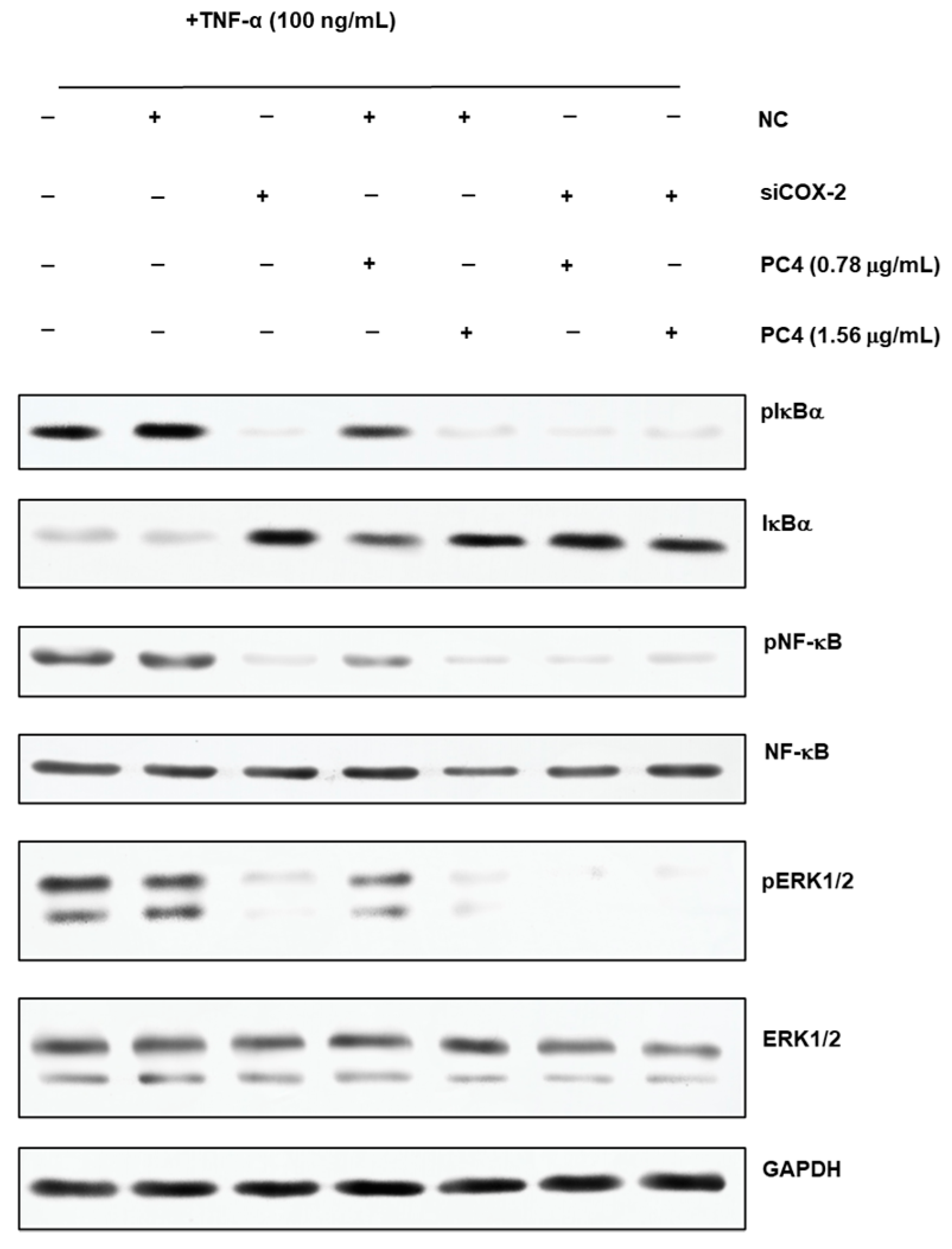
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
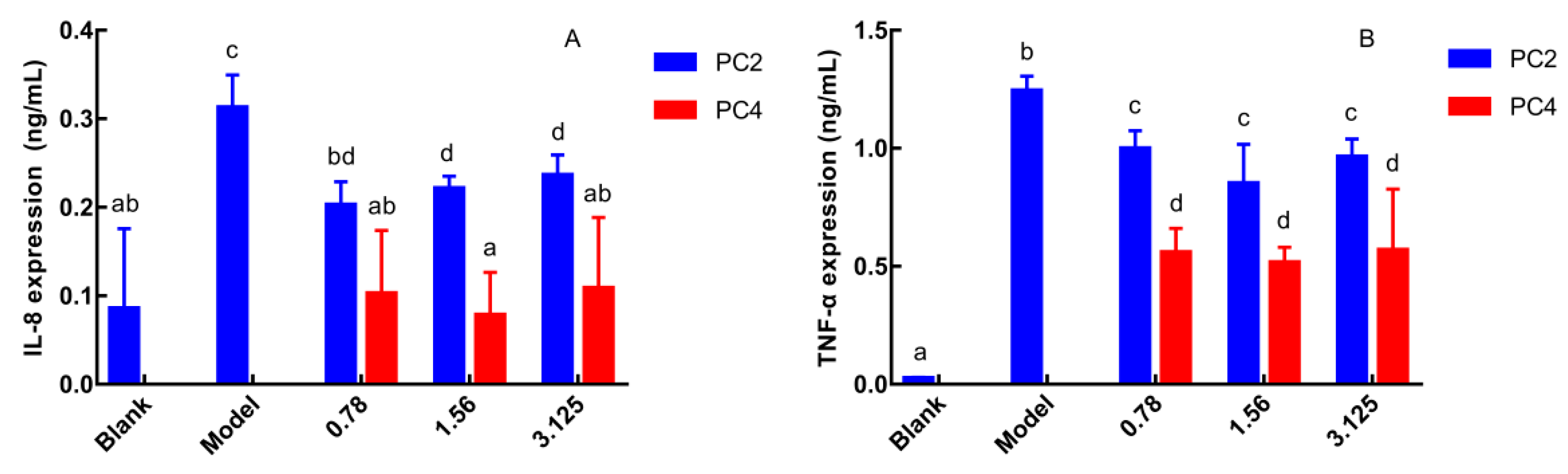
3.1. Comparison of PC2 and PC4 in Anti-Inflammatory Activities
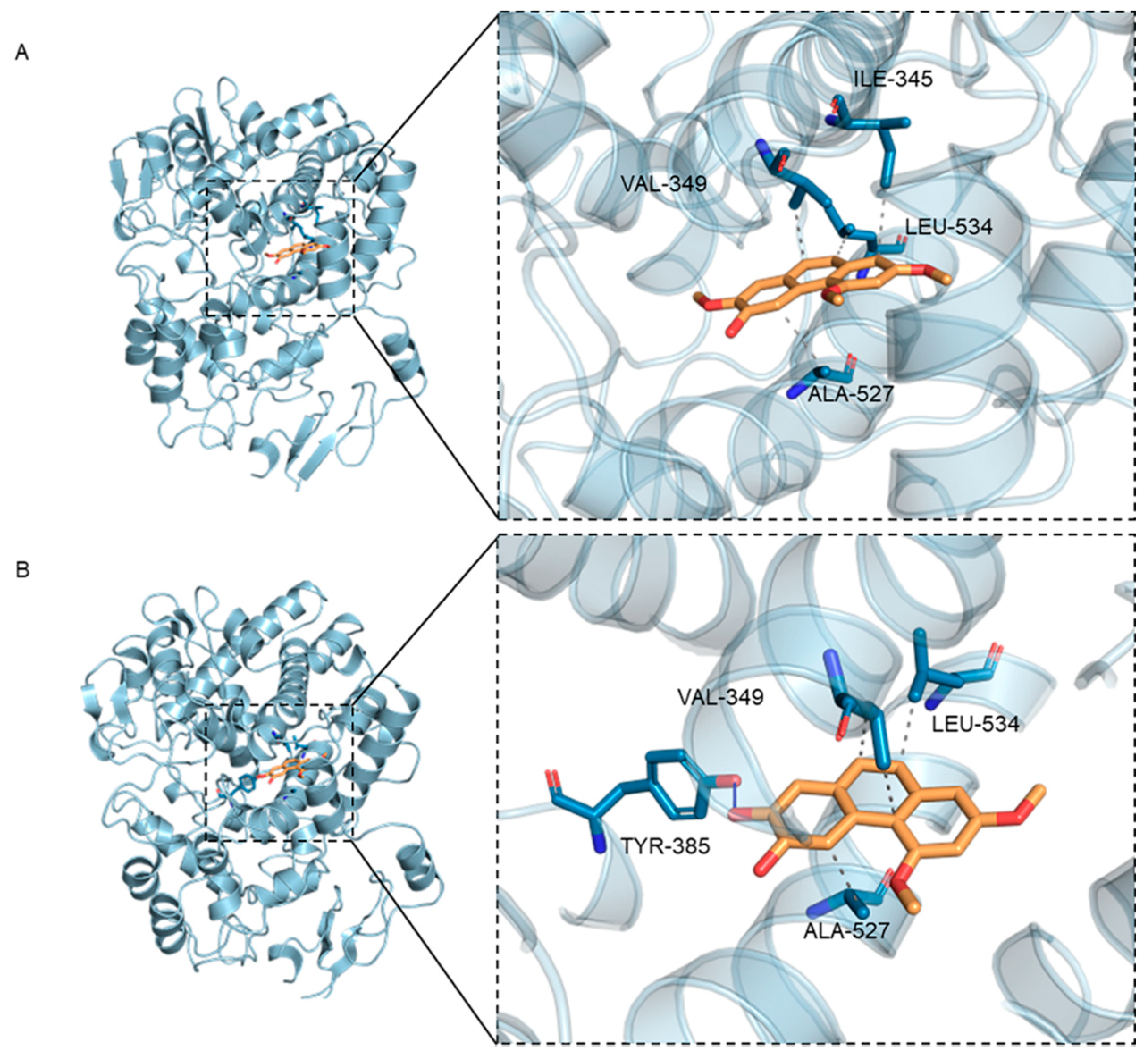
3.2. Molecular Docking
3.2.1. Molecular Docking Analysis
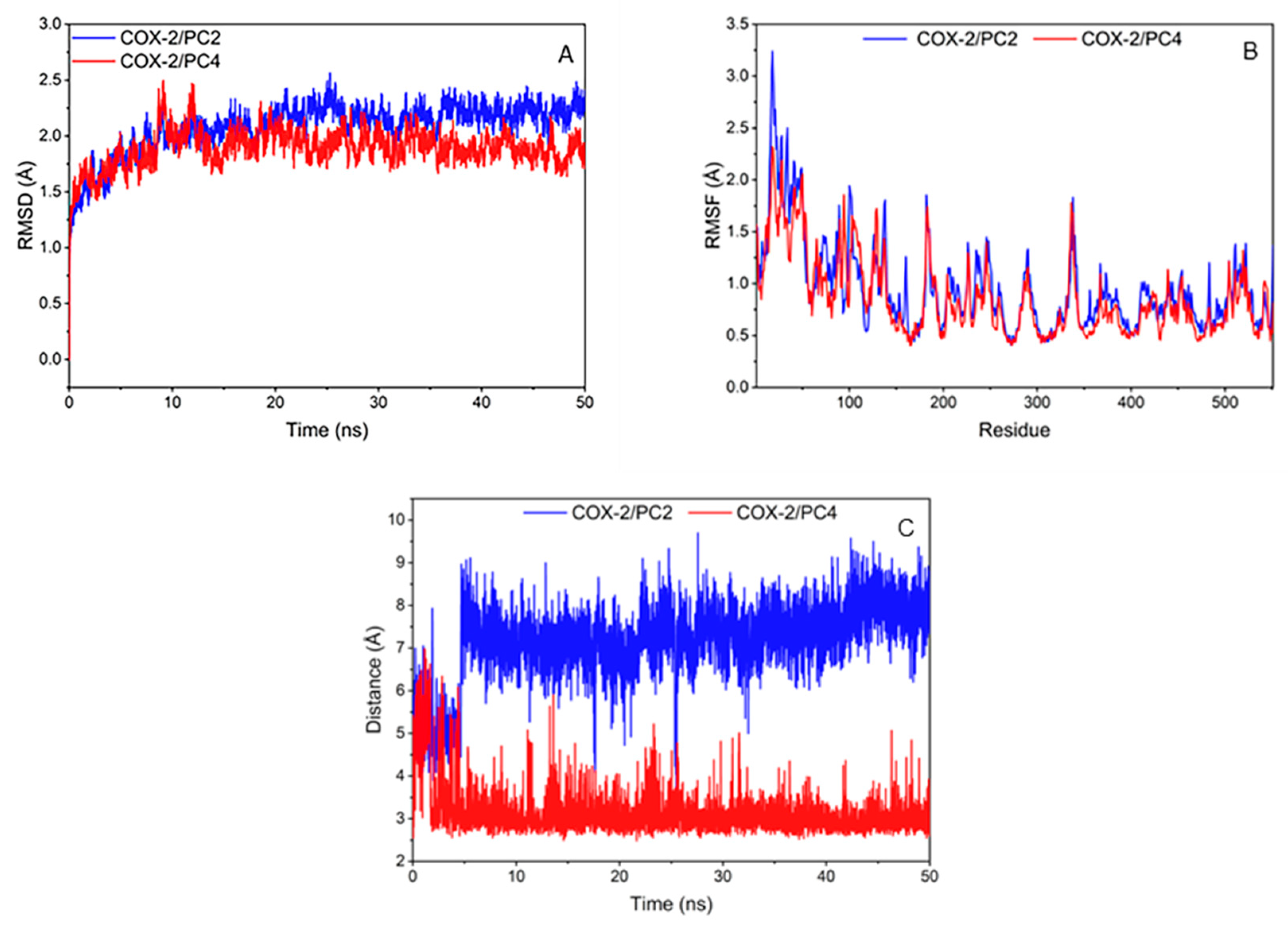
3.2.2. Molecular Dynamics Simulation Analysis
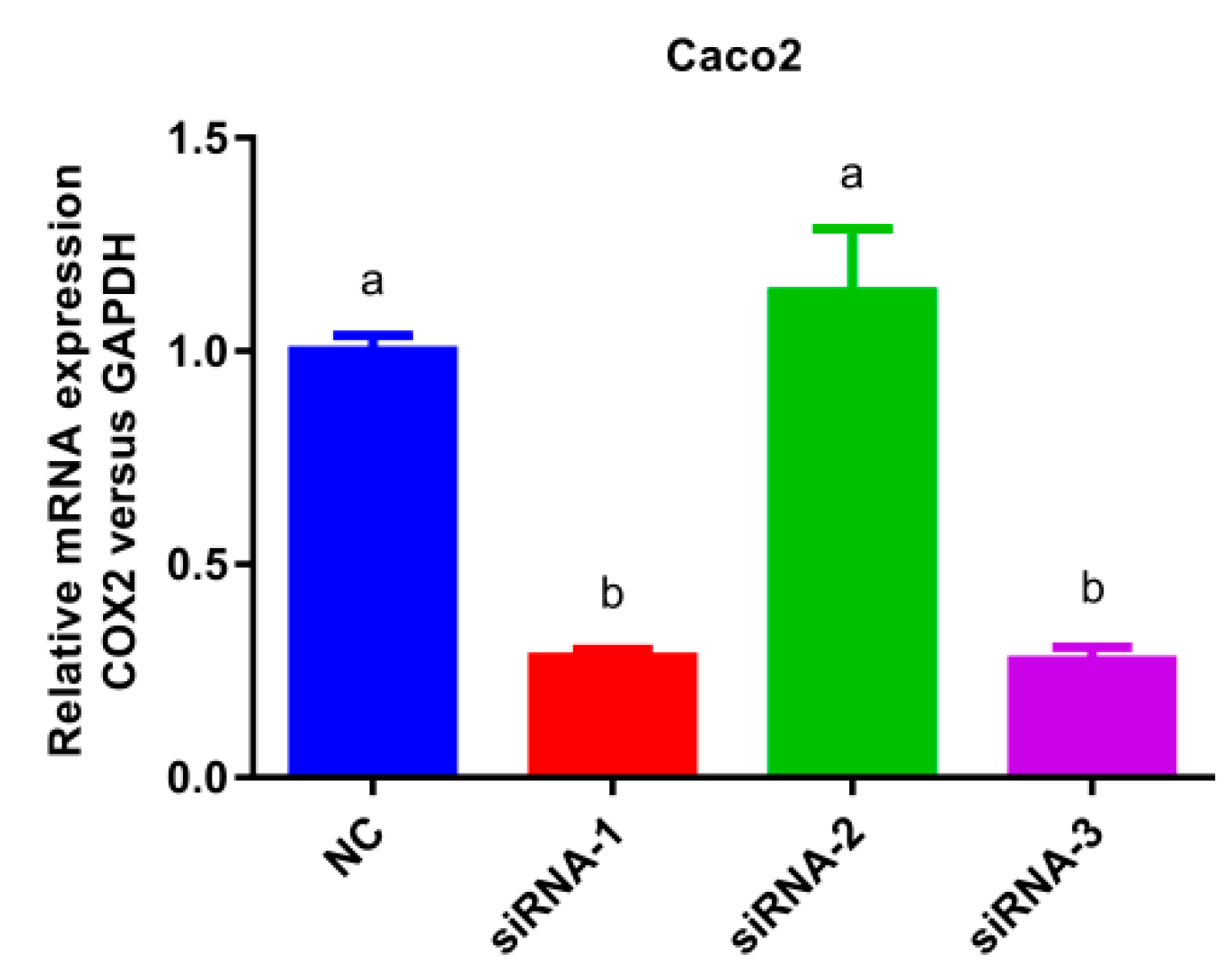
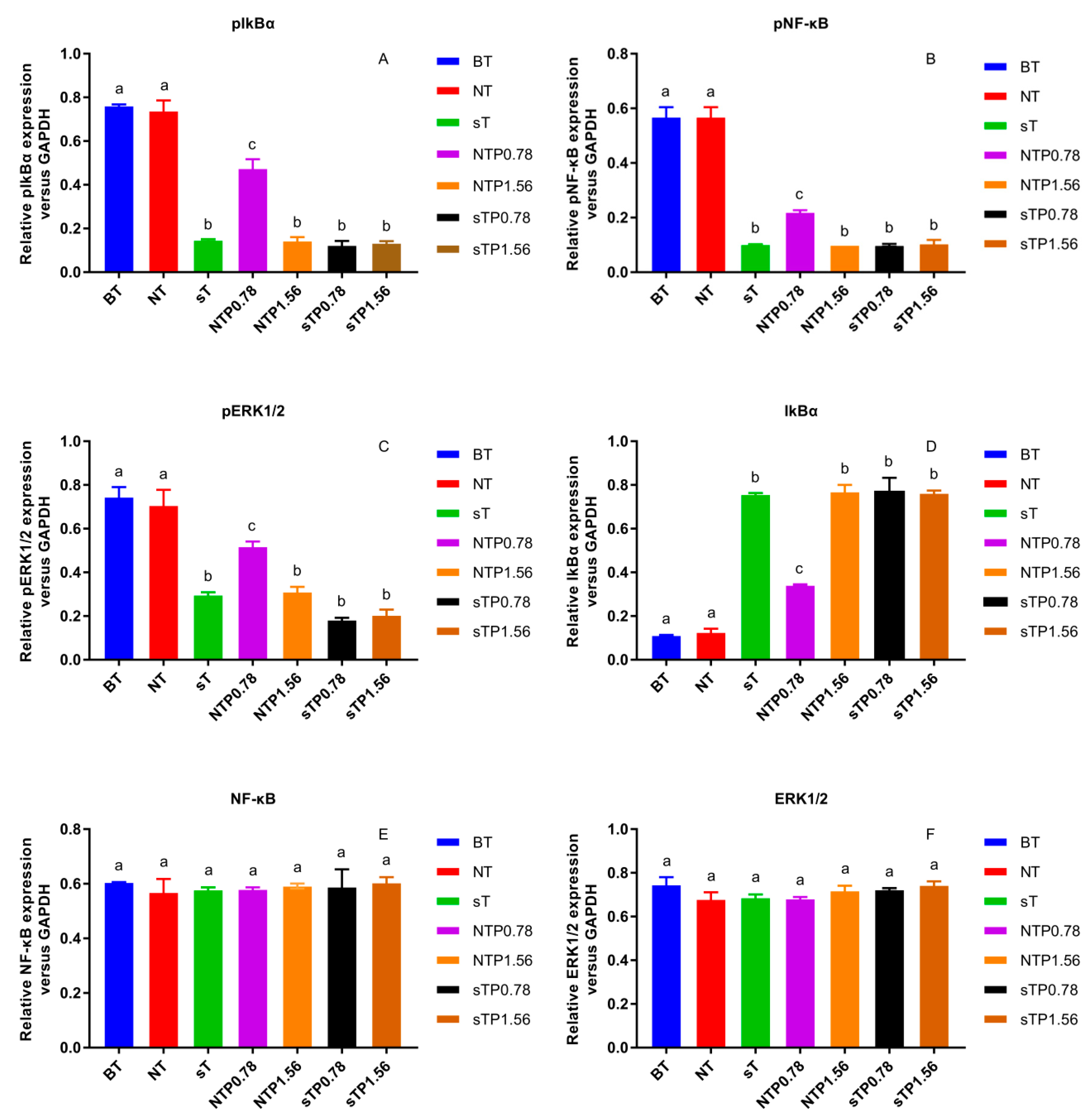
3.3. COX-2 Gene Silencing
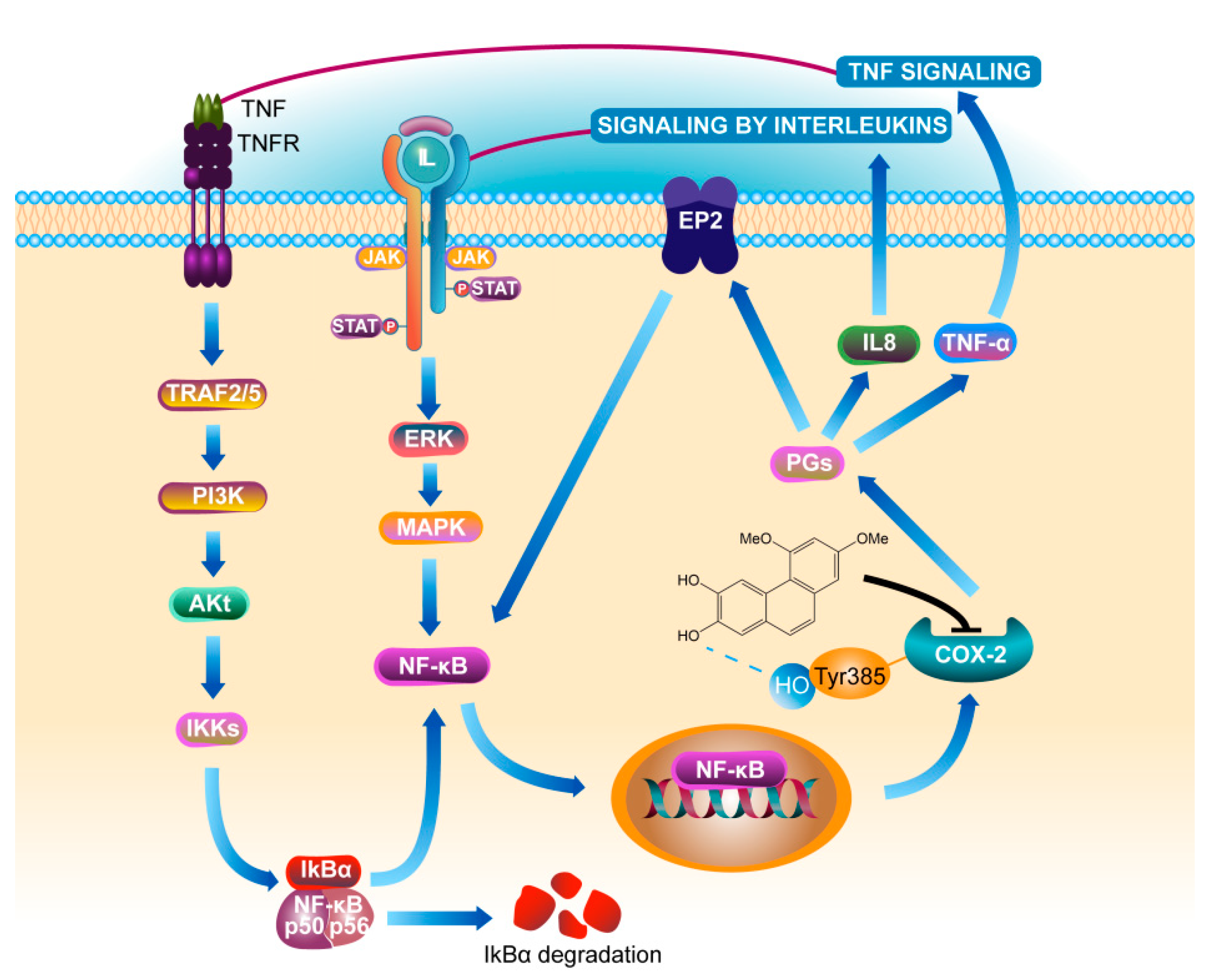
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Epping, J.; Laibach, N. An underutilized orphan tuber crop-Chinese yam: A review. Planta 2020, 252, 58. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Li, J.; Tao, H.; Zhao, H.; Liu, P.; Cui, B. Physicochemical properties and in vitro digestibility of hydrothermal treated Chinese yam (Dioscorea opposita Thunb.) starch and flour. Int. J. Biol. Macromol. 2021, 176, 177–185. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Chen, G. Extraction, structural analysis, derivatization and antioxidant activity of polysaccharide from Chinese yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef] [PubMed]
- Woo, K.W.; Kwon, O.W.; Kim, S.Y.; Choi, S.Z.; Son, M.W.; Kim, K.H.; Lee, K.R. Phenolic derivatives from the rhizomes of Dioscorea nipponica and their anti-neuroinflammatory and neuroprotective activities. J. Ethnopharmacol. 2014, 155, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.Y.-Z.; Zhang, Z.-X.; Zhou, W.; Cao, H.; Xiang, F.-J. New phenanthrene glycosides from Dioscorea opposita. J. Asian Nat. Prod. Res. 2014, 16, 148–152. [Google Scholar] [CrossRef]
- Yang, M.H.; Yoon, K.D.; Chin, Y.-W.; Park, J.H.; Kim, S.H.; Kim, Y.C.; Kim, J. Neuroprotective effects of Dioscorea opposita on scopolamine-induced memory impairment in in vivo behavioral tests and in vitro assays. J. Ethnopharmacol. 2009, 121, 130–134. [Google Scholar] [CrossRef]
- Tóth, B.; Hohmann, J.; Vasas, A. Phenanthrenes: A promising group of plant secondary metabolites. J. Nat. Prod. 2018, 81, 661–678. [Google Scholar] [CrossRef]
- Dissanayake, A.A.; Mmongoyo, J.A.; Nair, M.G. Zanzibar yam (Dioscorea sansibarensis Pax) isolates exhibit cyclooxygenase enzyme and lipid peroxidation inhibitory activities. Nat. Prod. Res. 2022, 36, 4719–4723. [Google Scholar] [CrossRef]
- Lim, J.S.; Hahn, D.; Gu, M.J.; Oh, J.; Lee, J.S.; Kim, J.-S. Anti-inflammatory and antioxidant effects of 2, 7-dihydroxy-4, 6-dimethoxy phenanthrene isolated from Dioscorea batatas Decne. Appl. Biol. Chem. 2019, 62, 29. [Google Scholar] [CrossRef]
- Lim, J.S.; Oh, J.; Yun, H.S.; Lee, J.S.; Hahn, D.; Kim, J.-S. Anti-neuroinflammatory activity of 6,7-dihydroxy-2,4-dimethoxy phenanthrene isolated from Dioscorea batatas Decne partly through suppressing the p38 MAPK/NF-κB pathway in BV2 microglial cells. J. Ethnopharmacol. 2022, 282, 114633. [Google Scholar] [CrossRef]
- Abbasi-Parizad, P.; De Nisi, P.; Scaglia, B.; Scarafoni, A.; Pilu, S.; Adani, F. Recovery of phenolic compounds from agro-industrial by-products: Evaluating antiradical activities and immunomodulatory properties. Food Bioprod. Process. 2021, 127, 338–348. [Google Scholar] [CrossRef]
- Huarte, E.; Serra, G.; Monteagudo-Mera, A.; Spencer, J.; Cid, C.; De Pena, M.-P. Raw and sous-vide-cooked red cardoon stalks (Cynara cardunculus L. var. altilis DC): (Poly)phenol bioaccessibility, anti-inflammatory activity in the gastrointestinal tract, and prebiotic activity. J. Agric. Food. Chem. 2021, 69, 9270–9286. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Zhang, H.; Fan, W.Y.; Mats, L.; Liu, R.H.; Deng, Z.Y.; Tsao, R. Anti-inflammatory effect and cellular transport mechanism of phenolics from common bean (Phaseolus vulga L.) milk and yogurts in Caco-2 mono- and Caco-2/EA.hy926 co-culture models. J. Agric. Food. Chem. 2021, 69, 1513–1523. [Google Scholar] [CrossRef]
- Zhao, Z.-H.; Xin, F.-Z.; Xue, Y.; Hu, Z.; Han, Y.; Ma, F.; Zhou, D.; Liu, X.-L.; Cui, A.; Liu, Z.; et al. Indole-3-propionic acid inhibits gut dysbiosis and endotoxin leakage to attenuate steatohepatitis in rats. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef]
- Jeong, J.B.; Hong, S.C.; Jeong, H.J.; Koo, J.S. Anti-inflammatory Effect of 2-methoxy-4-vinylphenol via the suppression of NF-kappa B and MAPK activation, and acetylation of histone H3. Arch. Pharmacal Res. 2011, 34, 2109–2116. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Huynh, D.T.N.; Jin, Y.; Jeon, H.; Heo, K.S. Protective effects of ginsenoside-Rg2 and-Rh1 on liver function through inhibiting TAK1 and STAT3-mediated inflammatory activity and Nrf2/ARE-mediated antioxidant signaling pathway. Arch. Pharmacal Res. 2021, 44, 241–252. [Google Scholar] [CrossRef]
- Chun, K.S.; Keum, Y.S.; Han, S.S.; Song, Y.S.; Kim, S.H.; Surh, Y.J. Curcumin inhibits phorbol ester-induced expression of cyclooxygenase-2 in mouse skin through suppression of extracellular signal-regulated kinase activity and NF-kappa B activation. Carcinogenesis 2003, 24, 1515–1524. [Google Scholar] [CrossRef]
- Aoki, T.; Narumiya, S. Prostaglandins and chronic inflammation. Trends Pharmacol. Sci. 2012, 33, 304–311. [Google Scholar] [CrossRef]
- Kikut, J.; Konecka, N.; Zietek, M.; Kulpa, D.; Szczuko, M. Diet supporting therapy for inflammatory bowel diseases. Eur. J. Nutr. 2021, 60, 2275–2291. [Google Scholar] [CrossRef]
- Shin, E.M.; Zhou, H.Y.; Guo, L.Y.; Kim, J.A.; Lee, S.H.; Merfort, I.; Kang, S.S.; Kim, H.S.; Kim, S.; Kim, Y.S. Anti-inflammatory effects of glycyrol isolated from Glycyrrhiza uralensis in LPS-stimulated RAW264.7 macrophages. Int. Immunopharmacol. 2008, 8, 1524–1532. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, C.; Dissanayake, A.A.; Gao, Q.; Nair, M.G. Phenanthrenes in chinese yam peel exhibit antiinflammatory activity, as shown by strong in vitro cyclooxygenase enzyme inhibition. Nat. Prod. Commun. 2016, 11, 1313–1316. [Google Scholar] [CrossRef]
- Li, Q.; Li, K.; Hu, T.; Liu, F.; Liao, S.; Zou, Y. 6,7-Dihydroxy-2,4-dimethoxyphenanthrene from Chinese yam peels alleviates DSS-induced intestinal mucosal injury in mice via modulation of the NF-kappa B/COX-2 signaling pathway. J. Agric. Food Chem. 2021, 69, 4720–4731. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, C.; Liu, F.; Hu, T.; Shen, W.; Li, E.; Liao, S.; Zou, Y. Mulberry leaf polyphenols attenuated postprandial glucose absorption via inhibition of disaccharidases activity and glucose transport in Caco-2 cells. Food Funct. 2020, 11, 1835–1844. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, Y.X.; Fong, P.; Mao, S.J.; Wang, Q.T. The application of the MM/GBSA method in the binding pose prediction of FGFR inhibitors. PCCP 2020, 22, 9656–9663. [Google Scholar] [CrossRef]
- Gao, J.; Chen, F.; Fang, H.; Mi, J.; Qi, Q.; Yang, M. Daphnetin inhibits proliferation and inflammatory response in human HaCaT keratinocytes and ameliorates imiquimod-induced psoriasis-like skin lesion in mice. Biol. Res. 2020, 53, 48. [Google Scholar] [CrossRef]
- Das, S.K.; Mahanta, S.; Tanti, B.; Tag, H.; Hui, P.K. Identification of phytocompounds from Houttuynia cordata Thunb. as potential inhibitors for SARS-CoV-2 replication proteins through GC-MS/LC-MS characterization, molecular docking and molecular dynamics simulation. Mol. Divers. 2022, 26, 365–388. [Google Scholar] [CrossRef]
- Meshram, R.J.; Bagul, K.T.; Pawnikar, S.P.; Barage, S.H.; Kolte, B.S.; Gacche, R.N. Known compounds and new lessons: Structural and electronic basis of flavonoid-based bioactivities. J. Biomol. Struct. Dyn. 2020, 38, 1168–1184. [Google Scholar] [CrossRef]
- Erez, N.; Truitt, M.; Olson, P.; Hanahan, D. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappa B-dependent manner. Cancer Cell 2010, 17, 135–147. [Google Scholar] [CrossRef]
- Guo, C.; Wang, Y.; Zhang, S.; Zhang, X.; Du, Z.; Li, M.; Ding, K. Crataegus pinnatifida polysaccharide alleviates colitis via modulation of gut microbiota and SCFAs metabolism. Int. J. Biol. Macromol. 2021, 181, 357–368. [Google Scholar] [CrossRef]
- Shen, Y.; Teng, L.; Qu, Y.H.; Liu, J.; Zhu, X.D.; Chen, S.; Yang, L.F.; Huang, Y.H.; Song, Q.; Fu, Q. Anti-proliferation and anti-inflammation effects of corilagin in rheumatoid arthritis by downregulating NF-kappa B and MAPK signaling pathways. J. Ethnopharmacol. 2022, 284, 114791. [Google Scholar] [CrossRef]
- Danese, S.; Fiocchi, C. Medical progress: Ulcerative colitis. N. Engl. J. Med. 2011, 365, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Clemente, M.G.; Tripathi, A.; Sapone, A.; Thakar, M.; Iacono, G.; Carroccio, A.; D’Agate, C.; et al. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Cho, S.O.; Kim, H. alpha-Lipoic acid inhibits expression of IL-8 by suppressing activation of MAPK, Jak/Stat, and NF-kappa B in H. pylori-infected gastric epithelial AGS cells. Yonsei Med. J. 2016, 57, 260–264. [Google Scholar] [CrossRef]
- Goh, J.; Baird, A.W.; O’Keane, C.; Watson, R.W.G.; Cottell, D.; Bernasconi, G.; Petasis, N.A.; Godson, C.; Brady, H.R.; MacMathuna, P. Lipoxin A(4) and aspirin-triggered 15-epi-lipoxin A(4) antagonize TNF-alpha-stimulated neutrophil-enterocyte interactions in vitro and attenuate TNF-alpha-induced chemokine release and colonocyte apoptosis in human intestinal mucosa ex vivo. J. Immunol. 2001, 167, 2772–2780. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.E.; Lapointe, G.R.; Feucht, P.H.; Hilt, S.; Gallegos, C.A.; Gordon, C.A.; Giedlin, M.A.; Mullenbach, G.; Tekamp-Olson, P. IL-8 induces neutrophil chemotaxis predominantly via type I IL-8 receptors. J. Immunol. 1995, 155, 1428–1433. [Google Scholar] [CrossRef]
- Vecchio, A.J.; Simmons, D.M.; Malkowski, M.G. Structural basis of fatty acid substrate binding to cyclooxygenase-2. J. Biol. Chem. 2010, 285, 22152–22163. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Marnett, L.J. Cyclooxygenases: Structural and functional insights. J. Lipid Res. 2009, 50, S29–S34. [Google Scholar] [CrossRef]
- Tang, X.B.; Huang, G.Z.; Zhang, T.; Li, S.Q. Elucidation of colon-protective efficacy of diosgenin in experimental TNBS-induced colitis: Inhibition of NF-kappa B/IkB-alpha and Bax/Caspase-1 signaling pathways. Biosci. Biotechnol. Biochem. 2020, 84, 1903–1912. [Google Scholar] [CrossRef]
- Miller, S.C.; Huang, R.; Sakamuru, S.; Shukla, S.J.; Attene-Ramos, M.S.; Shinn, P.; Van Leer, D.; Leister, W.; Austin, C.P.; Xia, M. Identification of known drugs that act as inhibitors of NF-kappa B signaling and their mechanism of action. Biochem. Pharmacol. 2010, 79, 1272–1280. [Google Scholar] [CrossRef] [Green Version]
- Nam, N.-H. Naturally occurring NF-kappa B inhibitors. Mini-Rev. Med. Chem. 2006, 6, 945–951. [Google Scholar] [CrossRef]
- Yuan, R.; Huang, L.; Du, L.-J.; Feng, J.-F.; Li, J.; Luo, Y.-Y.; Xu, Q.-M.; Yang, S.-L.; Gao, H.; Feng, Y.-L. Dihydrotanshinone exhibits an anti-inflammatory effect in vitro and in vivo through blocking TLR4 dimerization. Pharmacol. Res. 2019, 142, 102–114. [Google Scholar] [CrossRef]
- Aoki, T.; Nishimura, M.; Matsuoka, T.; Yamamoto, K.; Furuyashiki, T.; Kataoka, H.; Kitaoka, S.; Ishibashi, R.; Ishibazawa, A.; Miyamoto, S.; et al. PGE(2)-EP2 signalling in endothelium is activated by haemodynamic stress and induces cerebral aneurysm through an amplifying loop via NF-kappa B. Br. J. Pharmacol. 2011, 163, 1237–1249. [Google Scholar] [CrossRef]
- Maybin, J.A.; Hirani, N.; Jabbour, H.N.; Critchley, H.O.D. Novel roles for hypoxia and prostaglandin E-2 in the regulation of IL-8 during endometrial repair. Am. J. Pathol. 2011, 178, 1245–1256. [Google Scholar] [CrossRef]
- Kundu, J.K.; Na, H.K.; Chun, K.S.; Kim, Y.K.; Lee, S.J.; Lee, S.S.; Lee, O.S.; Sim, Y.C.; Surh, Y.J. Inhibition of phorbol ester-induced COX-2 expression by epigallocatechin gallate in mouse skin and cultured human mammary epithelial cells. J. Nutr. 2003, 133, 3805S–3810S. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) | |||
|---|---|---|---|
| Compounds | G-Score/(kJ·mol−1) | Hydrogen Binding | Hydrophobic Interaction |
| PC4 | −8.109 | Tyr385 | Val349, Ala527, Leu534 |
| PC2 | −7.997 | / | Ile345, Val349, Ala527, Leu534 |
| (B) | |||
| System Name | COX-2/PC2 | COX-2/PC4 | |
| ΔEvdw (van der Waals energy) | −39.88 ± 1.16 | −38.46 ± 2.18 | |
| ΔEelec (electrostatic energy) | −5.47 ± 2.22 | −9.73 ± 2.03 | |
| ΔGGB (electrostatic contribution to solvation) | 21.05 ± 1.71 | 21.37 ± 1.10 | |
| ΔGSA (non-polar contribution to solvation) | −5.34 ± 0.07 | −5.31 ± 0.11 | |
| ΔGbind (binding free energy/Gibbs free energy/) | −29.65 ± 1.77 | −32.14 ± 1.94 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, C.; Zou, Y.; Liao, S.; Gao, Q.; Li, Q. Molecular Targets and Mechanisms of 6,7-Dihydroxy-2,4-dimethoxyphenanthrene from Chinese Yam Modulating NF-κB/COX-2 Signaling Pathway: The Application of Molecular Docking and Gene Silencing. Nutrients 2023, 15, 883. https://doi.org/10.3390/nu15040883
Nie C, Zou Y, Liao S, Gao Q, Li Q. Molecular Targets and Mechanisms of 6,7-Dihydroxy-2,4-dimethoxyphenanthrene from Chinese Yam Modulating NF-κB/COX-2 Signaling Pathway: The Application of Molecular Docking and Gene Silencing. Nutrients. 2023; 15(4):883. https://doi.org/10.3390/nu15040883
Chicago/Turabian StyleNie, Congyi, Yuxiao Zou, Sentai Liao, Qunyu Gao, and Qian Li. 2023. "Molecular Targets and Mechanisms of 6,7-Dihydroxy-2,4-dimethoxyphenanthrene from Chinese Yam Modulating NF-κB/COX-2 Signaling Pathway: The Application of Molecular Docking and Gene Silencing" Nutrients 15, no. 4: 883. https://doi.org/10.3390/nu15040883