
Polyunsaturated Fatty Acids and Their Immunomodulatory Actions in Periodontal Disease

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
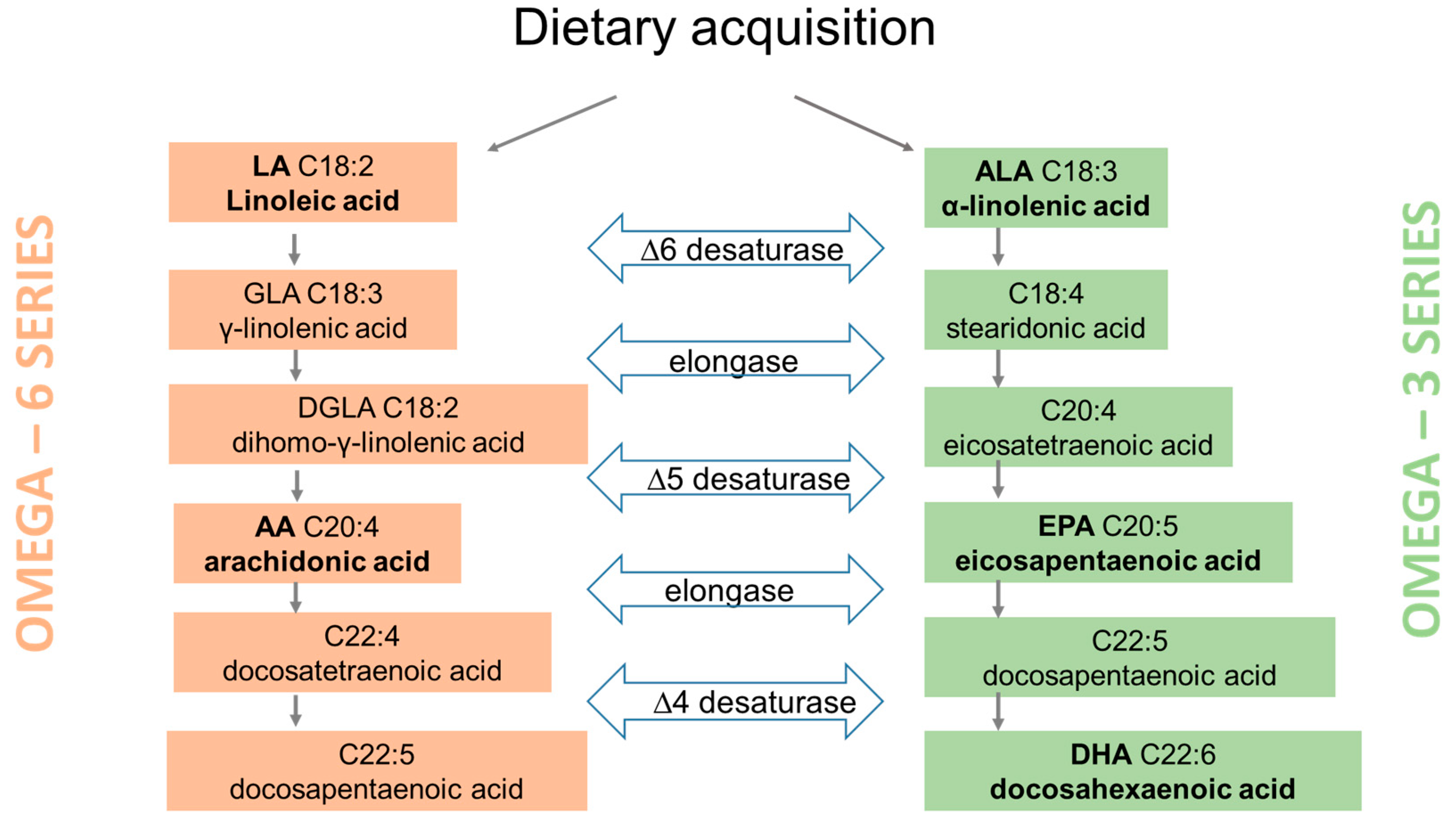
2. Fatty Acids
2.1. Metabolism
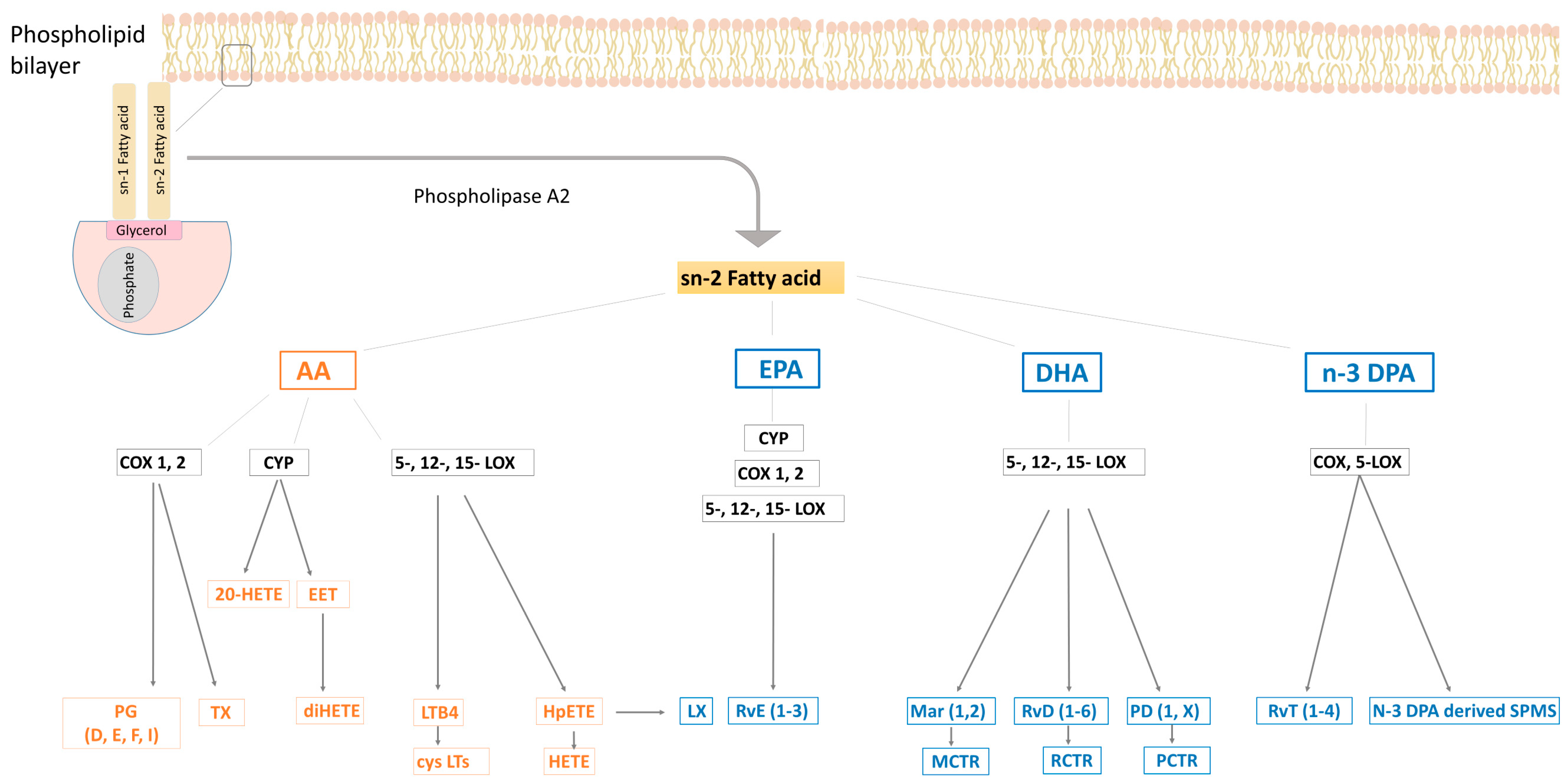
2.2. Bioactive Metabolites: Eicosanoids and SPMs
3. PUFAs in Immunity and Inflammation
3.1. N−6 Fatty Acids
3.2. N−3 Fatty Acids and SPMs
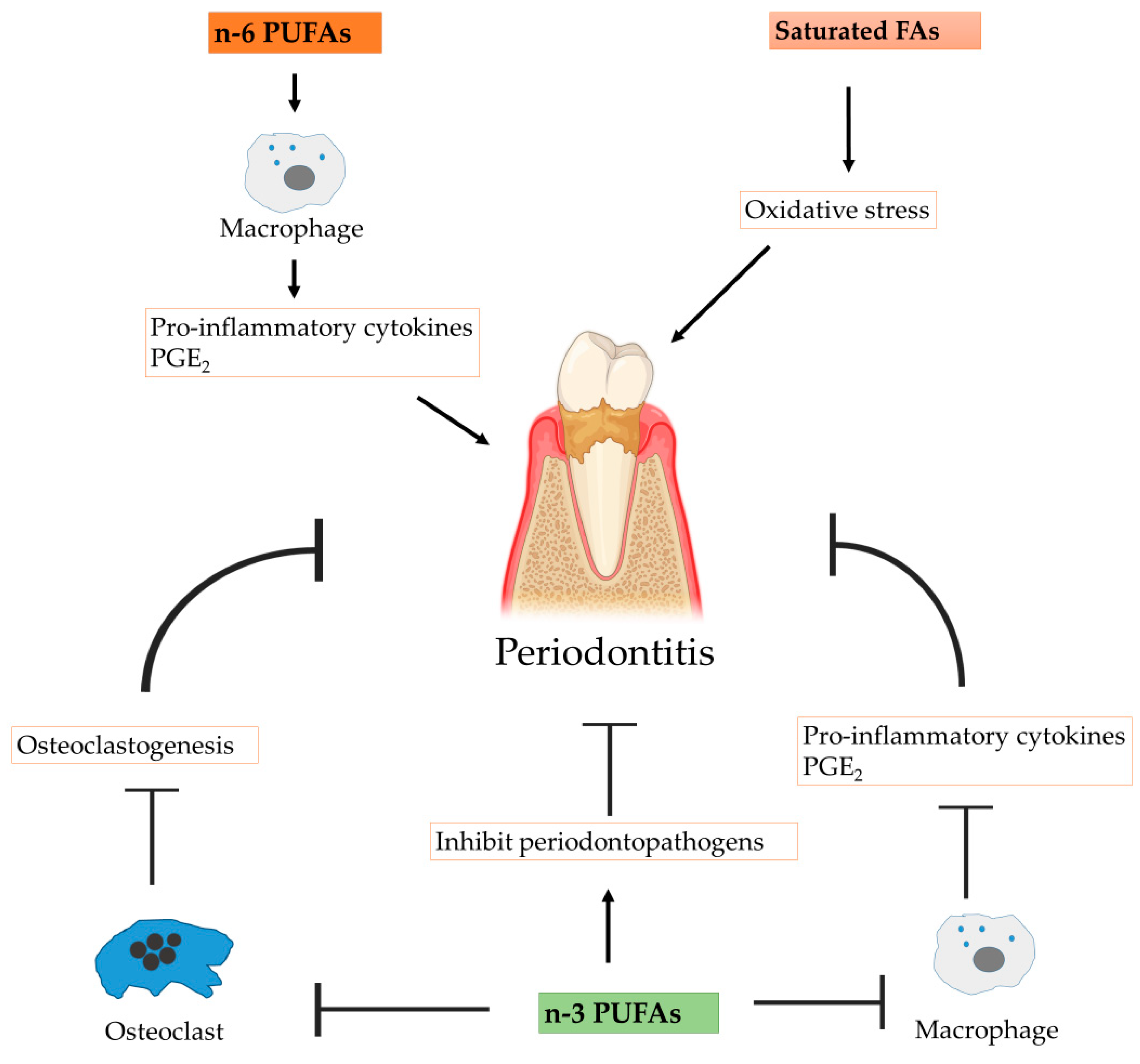
4. Immunomodulatory Impact in Periodontitis
4.1. Periodontitis
4.2. Anti-Inflammatory Actions
4.3. Protective Functions in Bone Metabolism
4.4. Antimicrobial Actions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef]
- Panezai, J.; Van Dyke, T.E. Resolution of inflammation: Intervention strategies and future applications. Toxicol. Appl. Pharmacol. 2022, 449, 116089. [Google Scholar] [CrossRef] [PubMed]
- Wiktorowska-Owczarek, A.; Berezinska, M.; Nowak, J.Z. PUFAs: Structures, metabolism and functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.L.; Leonard, A.E.; Mukerji, P. Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes. Prostaglandins Leukot Essent Fat. Acids 2003, 68, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C. Metabolism of alpha-linolenic acid in humans. Prostaglandins Leukot Essent Fat. Acids 2006, 75, 161–168. [Google Scholar] [CrossRef]
- Abeywardena, M.Y.; Patten, G.S. Role of ω3 long chain polyunsaturated fatty acids in reducing cardio-metabolic risk factors. Endocr. Metab. Immune Disord 2011, 11, 232–246. [Google Scholar] [CrossRef]
- Das, U.N. Essential fatty acids—A review. Curr. Pharm. Biotechnol. 2006, 7, 467–482. [Google Scholar] [CrossRef]
- Giltay, E.J.; Gooren, L.J.; Toorians, A.W.; Katan, M.B.; Zock, P.L. Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Am. J. Clin. Nutr. 2004, 80, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Khanapure, S.P.; Garvey, D.S.; Janero, D.R.; Gordon Letts, L. Eicosanoids in inflammation: Biosynthesis, pharmacology, and therapeutic frontiers. Curr. Top. Med. Chem. 2007, 7, 311–340. [Google Scholar] [CrossRef] [PubMed]
- von Moltke, J.; Trinidad, N.J.; Moayeri, M.; Kintzer, A.F.; Wang, S.B.; van Rooijen, N.; Brown, C.R.; Krantz, B.A.; Leppla, S.H.; Gronert, K.; et al. Rapid Induction of Inflammatory Lipid Mediators by the Inflammasome in Vivo. Nature 2012, 490, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Bozza, P.T.; Bakker-Abreu, I.; Navarro-Xavier, R.A.; Bandeira-Melo, C. Lipid body function in eicosanoid synthesis: An update. Prostagl. Leukot.Essent. Fat. Acids 2011, 85, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenases 1 and 2. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, dihommo-gamma linolenic, eicosanoids and inflammatory processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef]
- Serhan, C.N.; Petasis, N.A. Resolvins and protectins in inflammation resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.; Banthiya, S.; van Leyen, K. Mammalian lipoxygenases and their biological relevance. Biochim. Biophys. Acta 2015, 1851, 308–330. [Google Scholar] [CrossRef]
- Feinmark, S.J.; Cannon, P.J. Endothelial cell leukotriene C4 synthesis results from intercellular transfer of leukotriene A4 synthesized polymorphonuclear leukocytes. J. Biol. Chem. 1986, 261, 16466–16472. [Google Scholar] [CrossRef]
- Serhan, C.N. Pro-Resolving Lipid Mediators are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef]
- Spiecker, M.; Liao, J.K. Vascular protective effects of cytochrome p450 epoxygenase-derived eicosanoids. Arch. Biochem. Biophys. 2015, 433, 413–420. [Google Scholar] [CrossRef]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid Mediator Class Switching During Acute Inflammation: Signals in Resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef]
- Mori, T.A.; Hodgson, J.M. Fatty acids. In Encyclopedia of Human Nutrition; Elsevier: Amsterdam, The Netherlands, 2013; pp. 209–214. [Google Scholar]
- Thies, F.; Nebe-von-Caron, G.; Powell, J.R.; Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Dietary supplementation with γ -linolenic acid or fish oil decreases T lymphocyte proliferation in healthy older humans. J. Nutr. 2001, 131, 1918–1927. [Google Scholar] [CrossRef]
- Kelley, D.S.; Taylor, P.C.; Nelson, G.J.; Mackey, B.E. Arachidonic acid supplementation enhances synthesis of eicosanoids without suppressing immune functions in young healthy men. Lipids 1998, 33, 125–130. [Google Scholar] [CrossRef]
- Kew, S.; Banerjee, T.; Minihane, A.M.; Finnegan, Y.E.; Williams, C.M.; Calder, P.C. Relation between the fatty acid composition of peripheral blood mononuclear cells and measures of immune cell function in healthy, free-living subjects aged 25-72 y. Am. J. Clin. Nutr. 2003, 77, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P.; DiNicolantonio, J.J. The importance of a balanced omega-6 to omega-3 ratio in the prevention and management of obesity. Open Heart 2016, 3, e000385. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Immunomodulation by omega-3 fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 327–335. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; O’Keefe, J.H. Importance of maintaining a low omega-6/omega-3 ratio for reducing inflammation. Open Heart 2018, 5, e000946. [Google Scholar] [CrossRef]
- Fujikawa, M.; Yamashita, N.; Yamazaki, K.; Sugiyama, E.; Suzuki, H.; Hamazaki, T. Eicosapentaenoic Acid Inhibits Antigen-Presenting Cell Function of Murine Splenocytes. Immunology 1992, 75, 330–335. [Google Scholar]
- Calder, P.C.; Bevan, S.J.; Newsholme, E.A. The Inhibition of T-lymphocyte Proliferation by Fatty Acids Is via an Eicosanoid-Independent Mechanism. Immunology 1992, 75, 108–115. [Google Scholar]
- Betz, M.; Fox, B.S. Prostaglandin E2 Inhibits Production of Th1 Lymphokines but Not of Th2 Lymphokines. J. Immunol. 1991, 146, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Snijdewint, F.G.M.; Kalinski, P.; Wierenga, E.A.; Bos, J.D.; Kapsenberg, M.L. Prostaglandin E2 Differentially Modulates Cytokine Secretion Profiles of Human T Helper Lymphocytes. J. Immunol. 1993, 150, 5321–5329. [Google Scholar] [CrossRef]
- Kunkel, S.L.; Chensue, S.W.; Phan, S.H. Prostaglandins as Endogenous Mediators of Interleukin-1 Production. J. Immunol. 1986, 136, 186–192. [Google Scholar] [CrossRef]
- Renz, H.; Gong, J.-H.; Schmidt, A.; Nain, M.; Gemsa, D. Release of Tumor Necrosis Factor-α from Macrophages: Enhancement and Suppression Are Dose-Dependently Regulated by Prostaglandin E2 and Cyclic Nucleotides. J. Immunol. 1988, 141, 2388–2393. [Google Scholar] [CrossRef]
- Rode, H.N.; Szamel, M.; Schneider, S.; Resch, K. Phospholipid Metabolism of Stimulated Lymphocytes. Preferential Incorporation of Polyunsaturated Fatty Acids into Plasma Membrane Phospholipid upon Stimulation with Concanavalin A. Biochim. Biophys. Acta 1982, 688, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Sreeramkumar, V.; Fresno, M.; Cuesta, N. Prostaglandin E2 and T cells: Friends or foes? Immunol. Cell Biol. 2012, 90, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Raman, P.; Kaplan, B.L.; Kaminski, N.E. 15-Deoxy-Delta(1)(2),(1)(4)- prostaglandin J(2)-glycerol, a putative metabolite of 2-arachidonoyl glycerol and a peroxisome proliferator-activated receptor gamma ligand, modulates nuclear factor of activated T cells. J. Pharmacol. Exp. Ther. 2013, 342, 816–826. [Google Scholar] [CrossRef]
- Kumar, G.S.; Das, U.N. Effect of prostaglandins and their precursors on the proliferation of human lymphocytes and their secretion of tumor necrosis factor and various interleukins. Prostaglandins Leukot Essent Fat. Acids 1994, 50, 331–334. [Google Scholar] [CrossRef]
- Xue, L.; Barrow, A.; Fleming, V.M.; Hunter, M.G.; Ogg, G.; Klenerman, P.; Pettipher, R. Leukotriene E4 activates human Th2 cells for exaggerated proinflammatory cytokine production in response to prostaglandin D2. J. Immunol. 2011, 188, 694–702. [Google Scholar] [CrossRef]
- Ariel, A.; Chiang, N.; Arita, M.; Petasis, N.A.; Serhan, C.N. Aspirin-Triggered Lipoxin A 4 and B 4 Analogs Block Extracellular Signal-Regulated Kinase-Dependent TNF-α Secretion from Human T Cells. J. Immunol. 2003, 170, 6266–6272. [Google Scholar] [CrossRef] [PubMed]
- Harbige, L.S. Fatty acids, the immune response, and autoimmunity: A question of n− 6 essentiality and the balance between n− 6 and n− 3. Lipids 2003, 38, 323–341. [Google Scholar] [CrossRef]
- Matono, R.; Miyano, K.; Kiyohara, T.; Sumimoto, H. Arachidonic acid induces direct interaction of the p67(phox)-Rac complex with the phagocyte oxidase Nox2, leading to superoxide production. J. Biol. Chem. 2014, 289, 24874–24884. [Google Scholar] [CrossRef]
- Pompeia, C.; Cury-Boaventura, M.F.; Curi, R. Arachidonic acid triggers an oxidative burst in leukocytes. Braz. J. Med. Biol. Res. 2003, 36, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.C.; Dennis, E.A. Omega-3 fatty acids cause dramatic changes in TLR4 and purinergic eicosanoid signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 8517–8522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, H.; Zhang, W.; Cai, Y.; Shan, P.; Wu, D.; Zhang, B.; Liu, H.; Khan, Z.A.; Liang, G. Arachidonic acid inhibits inflammatory responses by binding to myeloid differentiation factor-2 (MD2) and preventing MD2/toll-like receptor 4 signaling activation. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165683. [Google Scholar] [CrossRef] [PubMed]
- Marion-Letellier, R.; Butler, M.; Dechelotte, P.; Playford, R.J.; Ghosh, S. Comparison of cytokine modulation by natural peroxisome proliferator-activated receptor gamma ligands with synthetic ligands in intestinal-like Caco-2 cells and human dendritic cells—Potential for dietary modulation of peroxisome proliferator-activated receptor gamma in intestinal inflammation. Am. J. Clin. Nutr. 2008, 87, 939–94828. [Google Scholar] [PubMed]
- Yaqoob, P.; Pala, H.S.; Cortina-Borja, M.; Newsholme, E.A.; Calder, P.C. Encapsulated fish oil enriched in -tocopherol alters plasma phospholipid and mononuclear cell fatty acid compositions but not mononuclear cell functions. Eur. J. Clin. Investig. 2000, 30, 260–274. [Google Scholar] [CrossRef]
- Healy, D.A.; Wallace, F.A.; Miles, E.A.; Calder, P.C.; Newsholme, P. The effect of low to moderate amounts of dietary fish oil on neutrophil lipid composition and function. Lipids 2000, 35, 763–768. [Google Scholar] [CrossRef]
- Yaqoob, P.; Calder, P.C. Effects of dietary lipid manipulation upon inflammatory mediator production by murine macrophages. Cell Immunol. 1995, 163, 120–128. [Google Scholar] [CrossRef]
- Lee, T.H.; Mencia-Huerta, J.M.; Shih, C.; Corey, E.J.; Lewis, R.A.; Austen, K.F. Characterization and biologic properties of 5,12-dihydroxy derivatives of eicosapentaenoic acid, including leukotriene-B5 and the double lipoxygenase product. J. Biol. Chem. 1984, 259, 2383–2389. [Google Scholar] [CrossRef]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from w-6 and w-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef]
- Wada, M.; DeLong, C.J.; Hong, Y.H.; Rieke, C.J.; Song, I.; Sidhu, R.S.; Yuan, C.; Warnock, M.; Schmaier, A.H.; Yokoyama, C.; et al. Enzymes and receptors of prostaglandin pathways with arachidonic acid-derived versus eicosapentaenoic acid-derived substrates and products. J. Biol. Chem. 2007, 282, 22254–22266. [Google Scholar] [CrossRef]
- Gorjao, R.; Verlengia, R.; de Lima, T.M.; Soriano, F.G.; Boaventura, M.F.C.; Kanunfre, C.C.; Peres, C.M.; Sampaio, S.C.; Otton, R.; Folador, A.; et al. Effect of docosahexaenoic acid-rich fish oil supplementation on human leukocyte function. Clin. Nutr. 2006, 25, 923–928. [Google Scholar] [CrossRef]
- Miles, E.A.; Banerjee, T.; Dooper, M.M.; M'Rabet, L.; Graus, Y.M.; Calder, P.C. The influence of different combinations of gamma-linolenic acid, stearidonic acid and EPA on immune function in healthy young male subjects. Br. J. Nutr. 2004, 91, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Siga, L.H. Basic science for the clinician 39: NF-kappaB-function, activation, control, and consequences. J. Clin. Rheumatol. 2006, 12, 207–211. [Google Scholar] [CrossRef]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef]
- Babcock, T.A.; Novak, T.; Ong, E.; Jho, D.H.; Helton, W.S.; Espat, N.J. Modulation of lipopolysaccharide-stimulated macrophage tumor necrosis factor-a production by w-3 fatty acid is associated with differential cyclooxygenase-2 protein expression and is independent of interleukin-10. J. Surg. Res. 2002, 107, 135–139. [Google Scholar] [PubMed]
- Novak, T.E.; Babcock, T.A.; Jho, D.H.; Helton, W.S.; Espat, N.J. NF-kappa B inhibition by omega -3 fatty acids modulates LPS-stimulated macrophage TNF-alpha transcription. Am. J. Physiol. 2003, 284, L84–L89. [Google Scholar]
- Carlsson, J.A.; Wold, A.E.; Sandberg, A.S.; Ostman, S.M. The Polyunsaturated Fatty Acids Arachidonic Acid and Docosahexaenoic Acid Induce Mouse Dendritic Cells Maturation but Reduce T-Cell Responses in Vitro. PLoS ONE 2015, 10, 0143741. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Fan, Y.Y.; Barhoumi, R.; Smith, R.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress the localization and activation of signaling proteins at the immunological synapse in murine CD4+ T cells by affecting lipid raft formation. J. Immunol. 2008, 181, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Y.; Barhoumi, R.; Fan, Y.Y.; Rivera, G.M.; Hannoush, R.N.; McMurray, D.N.; Chapkin, R.S. n-3 polyunsaturated fatty acids suppress CD4(+) T cell proliferation by altering phosphatidylinositol-(4,5) -bisphosphate [PI(4,5)P-2] organization. Biochim. Et Biophys. Acta-Biomembr. 2016, 1858, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Fuentes, N.R.; Hou, T.Y.; Barhoumi, R.; Li, X.C.; Deutz, N.E.P.; Engelen, M.P.K.J.; McMurray, D.N.; Chapkin, R.S. Remodelling of primary human CD4(+) T cell plasma membrane order by n-3 PUFA. Br. J. Nutr. 2018, 119, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Zech, T.; Ejsing, C.S.; Gaus, K.; de Wet, B.; Shevchenko, A.; Simons, K.; Harder, T. Accumulation of raft lipids in T-cell plasma membrane domains engaged in TCR signalling. EMBO J. 2009, 28, 466–476. [Google Scholar] [CrossRef]
- Cucchi, D.; Camacho-Munoz, D.; Certo, M.; Niven, J.; Smith, J.; Nicolaou, A.; Mauro, C. Omega-3 polyunsaturated fatty acids impinge on CD4+ T cell motility and adipose tissue distribution via direct and lipid mediator-dependent effects. Cardiovasc. Res. 2020, 116, 1006–1020. [Google Scholar] [CrossRef]
- Onodera, T.; Fukuhara, A.; Shin, J.; Hayakawa, T.; Otsuki, M.; Shimomura, I. Eicosapentaenoic acid and 5-HEPE enhance macrophage-mediated Treg induction in mice. Sci. Rep. 2017, 7, 4560. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Chiang, N. Protectins and maresins: New pro-resolving families of mediators in acute inflammation and resolution bioactive metabolome. Biochim. Biophys Acta Mol Cell Biol Lipids 2015, 1851, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Dalli, J.; Karamnov, S.; Choi, A.; Park, C.K.; Xu, Z.Z.; Ji, R.R.; Zhu, M.; Petasis, N.A. Macrophage proresolving mediator maresin 1 stimulates tissue regeneration and controls pain. FASEB J. 2012, 26, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Dalli, J.; Zhu, M.; Vlasenko, N.A.; Deng, B.; Haeggström, J.Z.; Petasis, N.A.; Serhan, C.N. The novel 13S,14S-epoxy-maresin is converted by human macrophages to maresin 1 (MaR1), inhibits leukotriene A4 hydrolase (LTA4H), and shifts macrophage phenotype. FASEB J. 2013, 27, 2573–2583. [Google Scholar] [CrossRef] [PubMed]
- Matte, A.; Recchiuti, A.; Federti, E.; Koehl, B.; Mintz, T.; El Nemer, W.; Tharaux, P.L.; Brousse, V.; Andolfo, I.; Lamolinara, A.; et al. Resolution of sickle cell disease associated inflammation and tissue damage with 17R-resolvin D1. Blood J. Am. Soc. Hematol. 2019, 133, 252–265. [Google Scholar] [CrossRef]
- Norling, L.V.; Spite, M.; Yang, R.; Flower, R.J.; Perretti, M.; Serhan, C.N. Cutting edge: Humanized nano-proresolving medicines mimic inflammation-resolution and enhance wound healing. J. Immunol. 2011, 186, 5543–5547. [Google Scholar] [CrossRef] [PubMed]
- Eke, P.I.; Dye, B.A.; Wei, L.; Thornton-Evans, G.O.; Genco, R.J. Prevalence of periodontitis in adults in the United States: 2009 and 2010. J. Dent. Res. 2012, 91, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Kassebaum, N.J.; Smith, A.G.; Bernabé, E.; Fleming, T.D.; Reynolds, A.E.; Vos, T.; Murray, C.J.; Marcenes, W.; GBD 2015 Oral Health Collaborators. Global, regional, and national prevalence, incidence, and disability-adjusted life years for oral conditions for 195 countries, 1990–2015: A systematic analysis for the global burden of diseases, injuries, and risk factors. J. Dent. Res. 2017, 96, 380–387. [Google Scholar] [CrossRef]
- Kosaka, T.; Ono, T.; Yoshimuta, Y.; Kida, M.; Kikui, M.; Nokubi, T.; Maeda, Y.; Kokubo, Y.; Watanabe, M.; Miyamoto, Y. The effect of periodontal status and occlusal support on masticatory performance: The Suita study. J. Clin. Periodontol. 2014, 41, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Genco, R.J.; Borgnakke, W.S. Risk factors for periodontal disease. Periodontol 2000 2013, 62, 59–94. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. The inflammophilic character of the periodontitis-associated microbiota. Mol. Oral. Microbiol. 2014, 29, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Tonetti, M.S.; Greenwell, H.; Kornman, K.S. Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J. Clin. Periodontol. 2018, 45, S149–S161. [Google Scholar] [CrossRef]
- Loos, B.G.; Van Dyke, T.E. The role of inflammation and genetics in periodontal disease. Periodontol. 2000 2020, 83, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Cantore, S.; Dedola, A.; Santacroce, L.; Laino, L.; Cicciù, M.; Mastrangelo, F. IL-1 haplotype analysis in periodontal disease. J. Biol. Regul. Homeost. Agents 2018, 32, 433–437. [Google Scholar]
- Scarel-Caminaga, R.M.; Kim, Y.J.; Viana, A.C.; Curtis, K.M.; Corbi, S.C.; Sogumo, P.M.; Orrico, S.R.; Cirelli, J.A. Haplotypes in the interleukin 8 gene and their association with chronic periodontitis susceptibility. Biochem. Genet. 2011, 49, 292–302. [Google Scholar] [CrossRef]
- Nibali, L.; Pelekos, G.; D’Aiuto, F.; Chaudhary, N.; Habeeb, R.; Ready, D.; Parkar, M.; Donos, N. Influence of IL-6 haplotypes on clinical and inflammatory response in aggressive periodontitis. Clin. Oral Investig. 2013, 17, 1235–1242. [Google Scholar] [CrossRef]
- Anovazzi, G.; Medeiros, M.C.; Pigossi, S.C.; Finoti, L.S.; Mayer, M.P.; Rossa, C.; Scarel-Caminaga, R.M. Functional haplotypes in interleukin 4 gene associated with periodontitis. PLoS ONE 2017, 12, e0169870. [Google Scholar] [CrossRef]
- Magnusson, I.; Lindhe, J.; Yoneyama, T.; Liljenberg, B. Recolonization of a subgingival microbiota following scaling in deep pockets. J. Clin. Periodontol. 1984, 11, 193–207. [Google Scholar] [CrossRef]
- Cortellini, P.; Buti, J.; Pini Prato, G.; Tonetti, M.S. Periodontal regeneration compared with access flap surgery in human intra-bony defects 20-year follow-up of a randomized clinical trial: Tooth retention, periodontitis recurrence and costs. J. Clin. Periodontol. 2017, 44, 58–66. [Google Scholar] [CrossRef]
- Preshaw, P.M. Host response modulation in periodontics. Periodontol. 2000 2008, 48, 92–110. [Google Scholar] [CrossRef] [PubMed]
- Kruse, A.B.; Kowalski, C.D.; Leuthold, S.; Vach, K.; Ratka-Krüger, P.; Woelber, J.P. What is the impact of the adjunctive use of omega-3 fatty acids in the treatment of periodontitis? A systematic review and meta-analysis. Lipids Health Dis. 2020, 19, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Araghizadeh, N.; Paknejad, M.; Alaeddini, M.; Minaii, B.; Abdollahi, M.; Khorasanie, R. The efficacy and prophylactic characteristics of omega-3 fatty acids in experimental gingivitis in rats. Iran J. Basic Med. Sci. 2014, 17, 87–92. [Google Scholar] [PubMed]
- Hankenson, K.D.; Watkins, B.A.; Schoenlein, I.A.; Allen, K.G.; Turek, J.J. Omega-3 fatty acids enhance ligament fibroblast collagen formation in association with changes in interleukin-6 production. Proc. Soc. Exp. Biol. Med. 2000, 223, 88–95. [Google Scholar] [CrossRef]
- Caughey, G.E.; Mantzioris, E.; Gibson, R.A.; Cleland, L.G.; James, M.J. The effect on human tumor necrosis factor α and interleukin 1β production of diets enriched in n-3 fatty acids from vegetable oil or fish oil. Am. J. Clin. Nutr. 1996, 63, 116–122. [Google Scholar] [CrossRef]
- Alam, S.Q.; Bergens, B.M.; Alam, B.S. Arachidonic acid, prostaglandin E2 and leukotriene C4 levels in gingiva and submandibular salivary glands of rats fed diets containing n-3 fatty acids. Lipids 1991, 26, 895–900. [Google Scholar] [CrossRef]
- Naqvi, A.Z.; Buettner, C.; Phillips, R.S.; Davis, R.B.; Mukamal, K.J. n-3 fatty acids and periodontitis in US adults. J. Am. Diet. Assoc. 2010, 110, 1669–1675. [Google Scholar] [CrossRef]
- Sierra, S.; Lara-Villoslada, F.; Comalada, M.; Olivares, M.; Xaus, J. Dietary eicosapentaenoic acid and docosahexaenoic acid equally incorporate as decosahexaenoic acid but differ in inflammatory effects. Nutrition 2008, 24, 245–254. [Google Scholar] [CrossRef]
- Collie-Duguid, E.; Wahle, K. Inhibitory effect of fish oil n−3 polyunsaturated fatty acids on the expression of endothelial cell adhesion molecules. Biochem. Biophys. Res. Commun. 1996, 220, 969–974. [Google Scholar] [CrossRef]
- Hughes, D.; Southon, S.; Pinder, A. (n−3) Polyunsaturated fatty acids modulate the expression of functionally associated molecules on human monocytes in vitro. J. Nutr. 1996, 126, 603–610. [Google Scholar] [CrossRef]
- Albelda, S.; Smith, C.; Ward, P. Adhesion molecules and inflammatory injury. FASEB J. 1994, 8, 504–512. [Google Scholar] [CrossRef]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef]
- Levy, B.D.; Kohli, P.; Gotlinger, K.; Haworth, O.; Hong, S.; Kazani, S.; Israel, E.; Haley, K.J.; Serhan, C.N. Protectin D1 is generated in asthma and dampens airway inflammation and hyperresponsiveness. J. Immunol. 2007, 178, 496–50229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, B.; Bokka, N.R.; Maddipati, K.R.; Ayilavarapu, S.; Weltman, R.; Zhu, L.; Chen, W.; Zheng, W.J.; Angelov, N.; Van Dyke, T.E.; et al. Distinct profiles of specialized pro-resolving lipid mediators and corresponding receptor gene expression in periodontal inflammation. Front. Immunol. 2020, 11, 1307. [Google Scholar] [CrossRef]
- Pouliot, M.; Clish, C.B.; Petasis, N.A.; Van Dyke, T.E.; Serhan, C.N. Lipoxin A(4) analogues inhibit leukocyte recruitment to Porphyromonas gingivalis: A role for cyclooxygenase-2 and lipoxins in periodontal disease. Biochemistry 2000, 39, 4761–4768. [Google Scholar] [CrossRef] [PubMed]
- Elabdeen, H.R.; Mustafa, M.; Szklenar, M.; Ruhl, R.; Ali, R.; Bolstad, A.I. Ratio of pro-resolving and pro-inflammatory lipid mediator precursors as potential markers for aggressive periodontitis. PLoS ONE 2013, 8, e70838. [Google Scholar] [CrossRef]
- Hong, S.; Porter, T.F.; Lu, Y.; Oh, S.F.; Pillai, P.S.; Serhan, C.N. Resolvin E1 metabolome in local inactivation during inflammation-resolution. J. Immunol. 2008, 180, 3512–3519. [Google Scholar] [CrossRef]
- Dalli, J.; Serhan, C.N. Specific lipid mediator signatures of human phagocytes: Microparticles stimulate macrophage efferocytosis and pro-resolving mediators. Blood J. Am. Soc. Hematol. 2012, 120, e60–e72. [Google Scholar] [CrossRef] [PubMed]
- Zarrough, A.E.; Hasturk, H.; Stephens, D.N.; Van Dyke, T.E.; Kantarci, A. Resolvin D1 modulates periodontal ligament fibroblast function. J. Periodontol. 2022. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, L.C.; Heufelder, A.E. Role of receptor activator of nuclear factor-kappaB ligand and osteoprotegerin in bone cell biology. J. Mol. Med. 2001, 79, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Kobayashi, Y.; Yamasaki, S.; Kawakami, A.; Eguchi, K.; Sasaki, H.; Sakai, H. Protein expression and functional difference of membrane-bound and soluble receptor activator of NFkappaB ligand: Modulation of the expression by osteotropic factors and cytokines. Biochem. Biophys. Res. Commun. 2000, 275, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Candelario, J.; Tavakoli, H.; Chachisvilis, M. PTH1 receptor is involved in mediating cellular response to long-chain polyunsaturated fatty acids. PLoS ONE 2012, 7, e52583. [Google Scholar] [CrossRef] [Green Version]
- Yeh, L.C.C.; Ford, J.J.; Lee, J.C.; Adamo, M.L. Palmitate attenuates osteoblast differentiation of fetal rat calvarial cells. Biochem. Biophys. Res Commun. 2014, 450, 777–781. [Google Scholar] [CrossRef]
- Kruger, M.C.; Coetzee, M.; Haag, M.; Weiler, H. Long-chain polyunsaturated fatty acids: Selected mechanisms of action on bone. Prog. Lipid Res. 2010, 49, 438–449. [Google Scholar] [CrossRef]
- Kruger, M.C.; Coetzee, M.; Haag, M.; Weiler, H. Long-chain polyunsaturated fatty acids may mutually benefit both obesity and osteoporosis. Nutr. Res. 2013, 33, 521–533. [Google Scholar]
- Azuma, M.M.; Gomes-Filho, J.E.; Cardoso, C.D.B.M.; Pipa, C.B.; Narciso, L.G.; Bomfim, S.R.M.; Jacinto, R.D.C.; Cintra, L.T.A. Omega 3 fatty acids reduce the triglyceride levels in rats with apical periodontitis. Braz. Dent. J. 2018, 29, 173–178. [Google Scholar] [CrossRef]
- Bendyk, A.; Marino, V.; Zilm, P.S.; Howe, P.; Bartold, P.M. Effect of dietary omega-3 polyunsaturated fatty acids on experimental periodontitis in the mouse. J. Periodontal. Res. 2009, 44, 211–216. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Z.; Zhang, X.; Yu, H.; Kirkwood, K.L.; Lopes-Virella, M.F.; Huang, Y. Metabolic syndrome exacerbates inflammation and bone loss in periodontitis. J. Dent. Res. 2015, 94, 362–370. [Google Scholar] [CrossRef]
- Choi, J.S.; Park, N.H.; Hwang, S.Y.; Sohn, J.H.; Kwak, I.; Cho, K.K.; Choi, I.S. The antibacterial activity of various saturated and unsaturated fatty acids against several oral pathogens. J. Environ. Biol. 2013, 34, 673–676. [Google Scholar] [PubMed]
- Kasonga, A.E.; Deepak, V.; Kruger, M.C.; Coetzee, M. Arachidonic acid and do- cosahexaenoic acid suppress osteoclast formation and activity in human CD14+ monocytes, in vitro. PLoS ONE 2015, 10, e0125145. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Bhattacharya, A.; Fernandes, G. Docosahexaenoic acid is more potent inhibitor of osteoclast differentiation in RAW 264.7 cells than eicosapentaenoic acid. J. Cell Physiol. 2008, 214, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Boeyens, J.C.; Deepak, V.; Chua, W.H.; Kruger, M.C.; Joubert, A.M.; Coetzee, M. Effects of omega 3-and omega 6-polyunsaturated fatty acids on RANKL-induced osteoclast differentiation of RAW264.7 cells: A comparative in vitro study. Nutrients 2014, 6, 2584–2601. [Google Scholar] [CrossRef]
- Nakamura, I.; Duong, L.T.; Rodan, S.B.; Rodan, G.A. Involvement of alpha(v) beta3 integrins in osteoclast function. J. Bone Miner. Metab. 2007, 25, 337–344. [Google Scholar] [CrossRef]
- Kim, H.J.; Ohk, B.; Yoon, H.J.; Kang, W.Y.; Seong, S.J.; Kim, S.Y.; Yoon, Y.R. Docosahexaenoic acid signaling attenuates the proliferation and differentiation of bone marrow-de- rived osteoclast precursors and promotes apoptosis in mature osteoclasts. Cell Signal 2017, 29, 226–232. [Google Scholar] [CrossRef]
- Hasturk, H.; Kantarci, A.; Ohira, T.; Arita, M.; Ebrahimi, N.; Chiang, N.; Petasis, N.A.; Levy, B.D.; Serhan, C.N.; Van Dyke, T.E. RvE1 protects from local inflammation and osteoclast- mediated bone destruction in periodontitis. FASEB J. 2006, 20, 401–403. [Google Scholar] [CrossRef]
- Herrera, B.S.; Ohira, T.; Gao, L.; Omori, K.; Yang, R.; Zhu, M.; Muscara, M.N.; Serhan, C.N.; Van Dyke, T.E.; Gyurko, R. An endogenous regulator of inflammation, resolvin E1, modulates osteoclast differentiation and bone resorption. Br. J. Pharmacol. 2008, 155, 1214–1223. [Google Scholar] [CrossRef]
- Jovanovic, D.V.; Di Battista, J.A.; Martel-Pelletier, J.; Jolicoeur, F.C.; He, Y.; Zhang, M.; Mineau, F.; Pelletier, J.P. IL-17 stimulates the production and expression of proinflammatory cytokines, IL-beta and TNF-alpha, by human macrophages. J. Immunol. 1998, 160, 3513–3521. [Google Scholar] [CrossRef]
- Pacifici, R. The role of IL-17 and TH17 cells in the bone catabolic activity of PTH. Front. Immunol. 2016, 7, 57. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Leuti, A.; Dalli, J.; Jacobsson, A.; Battistini, L.; Maccarrone, M.; Serhan, C.N. Pro-resolving lipid mediators Resolvin D1, Resolvin D2 and Maresin 1 are critical in modulating T cell responses. Sci. Transl. Med. 2016, 8, 353ra111. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Ebersole, J.L. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral Microbiol. 2010, 25, 75–80. [Google Scholar] [CrossRef]
- Ribeiro-Vidal, H.; Sánchez, M.C.; Alonso-Español, A.; Figuero, E.; Ciudad, M.J.; Collado, L.; Herrera, D.; Sanz, M. Antimicrobial activity of EPA and DHA against oral pathogenic bacteria using an in vitro multi-species subgingival biofilm model. Nutrients 2020, 12, 2812. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P. Potential applications of antimicrobial fatty acids in medicine, agriculture and other industries. Recent Pat. Antiinfect Drug Discov. 2012, 7, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.; Michel, V.; Matos, A.A.; Carvalho, P.; Oliveira, M.J.; Ferreira, R.M.; Dillies, M.A.; Huerre, M.; Seruca, R.; Figueiredo, C.; et al. Docosahexaenoic acid inhibits Helicobacter pylori growth in vitro and mice gastric mucosa colonization. PLoS ONE 2012, 7, e35072. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panezai, J.; van Dyke, T. Polyunsaturated Fatty Acids and Their Immunomodulatory Actions in Periodontal Disease. Nutrients 2023, 15, 821. https://doi.org/10.3390/nu15040821
Panezai J, van Dyke T. Polyunsaturated Fatty Acids and Their Immunomodulatory Actions in Periodontal Disease. Nutrients. 2023; 15(4):821. https://doi.org/10.3390/nu15040821
Chicago/Turabian StylePanezai, Jeneen, and Thomas van Dyke. 2023. "Polyunsaturated Fatty Acids and Their Immunomodulatory Actions in Periodontal Disease" Nutrients 15, no. 4: 821. https://doi.org/10.3390/nu15040821