

The Effects of Iron Deficiency on the Gut Microbiota in Women of Childbearing Age
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects Recruitment and Sample Collection
2.2. DNA Extraction from Stool Samples
2.3. 16S rRNA Gene-Based Metagenomics
2.4. Bioinformatics Analysis of 16S rRNA Gene Amplicons
2.5. qRT-PCR Analysis
3. Results
3.1. Patient Profiles with Baseline Characteristics
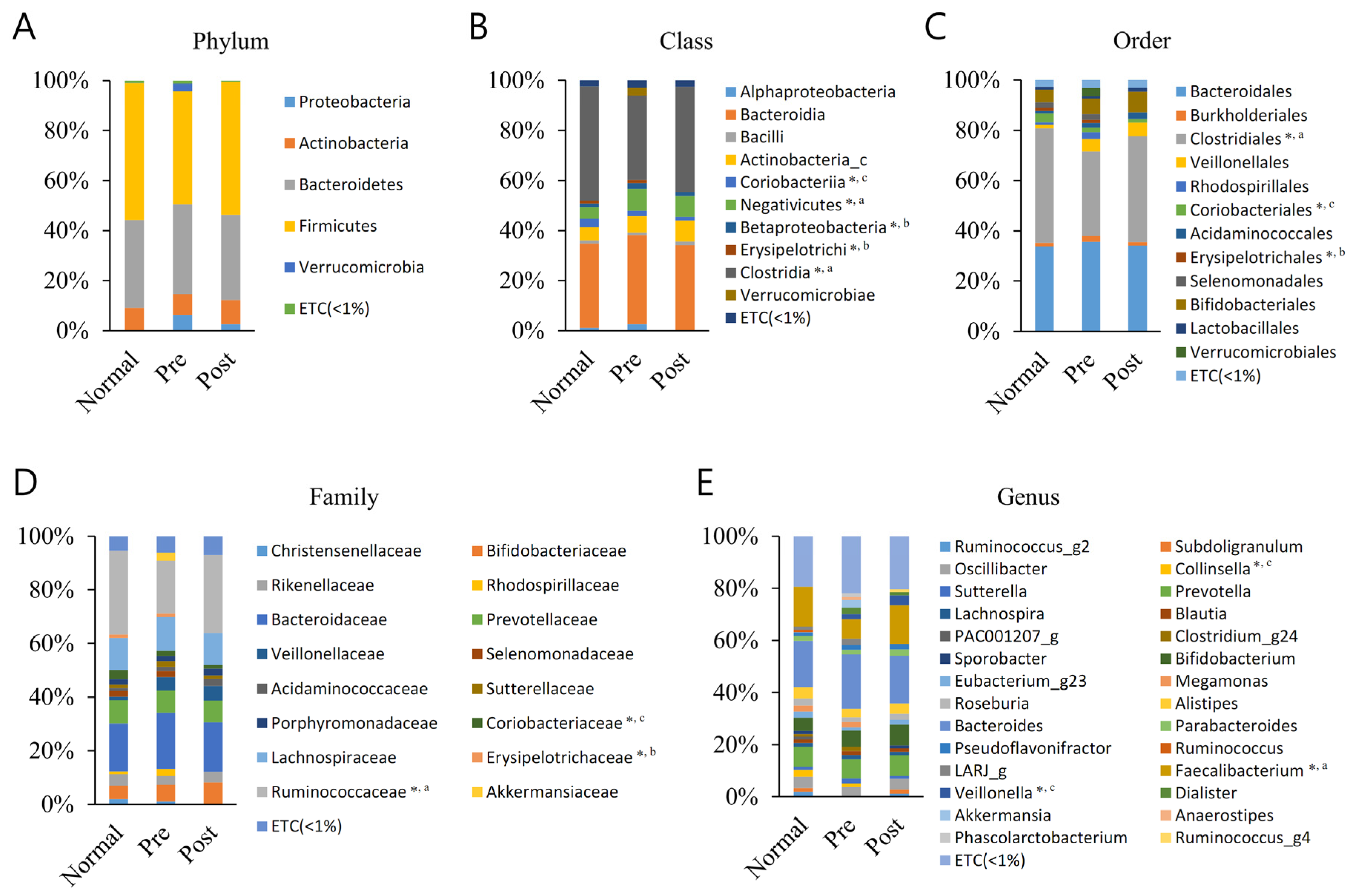
3.2. Averaged Taxonomic Composition
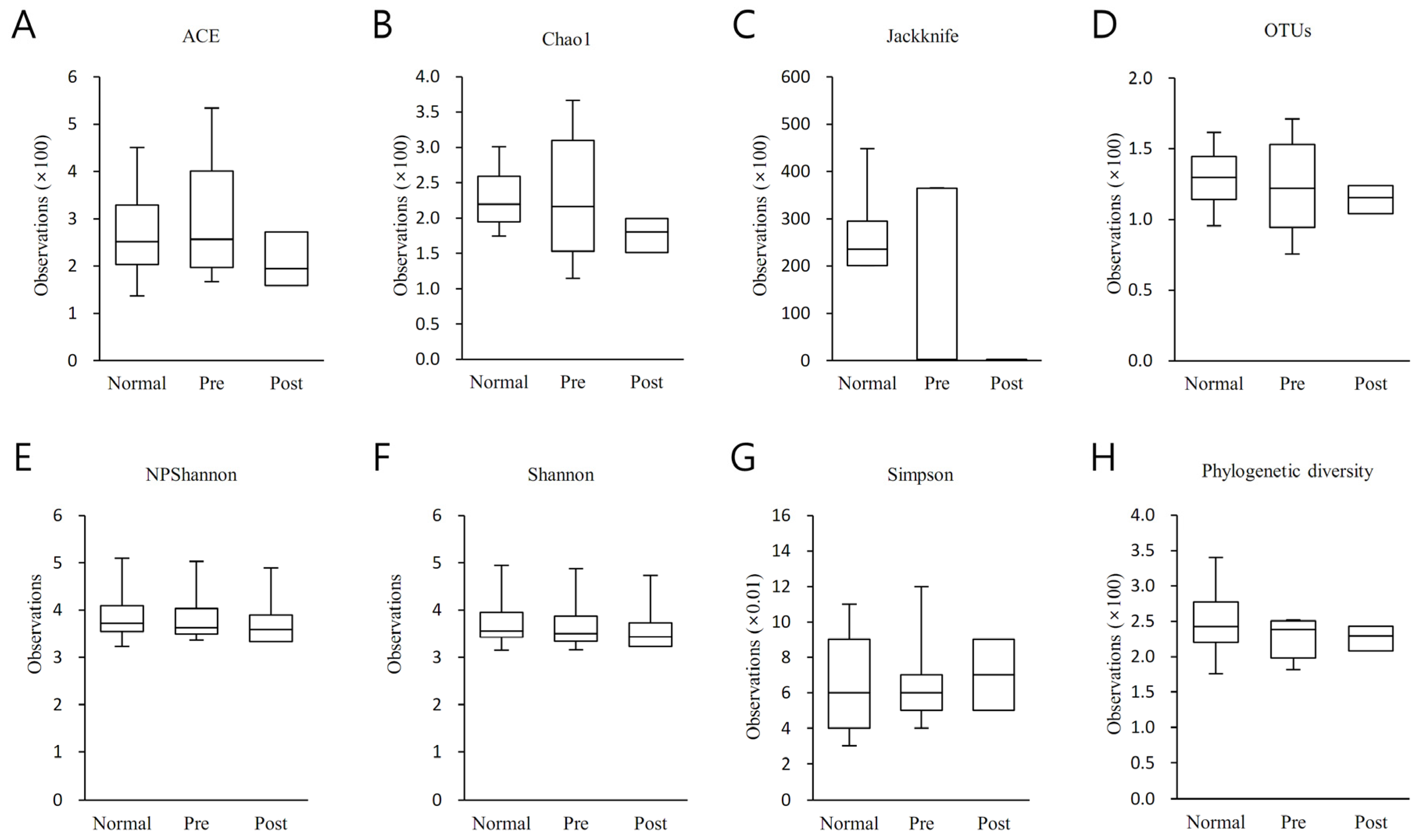
3.3. Alpha Diversity Analysis
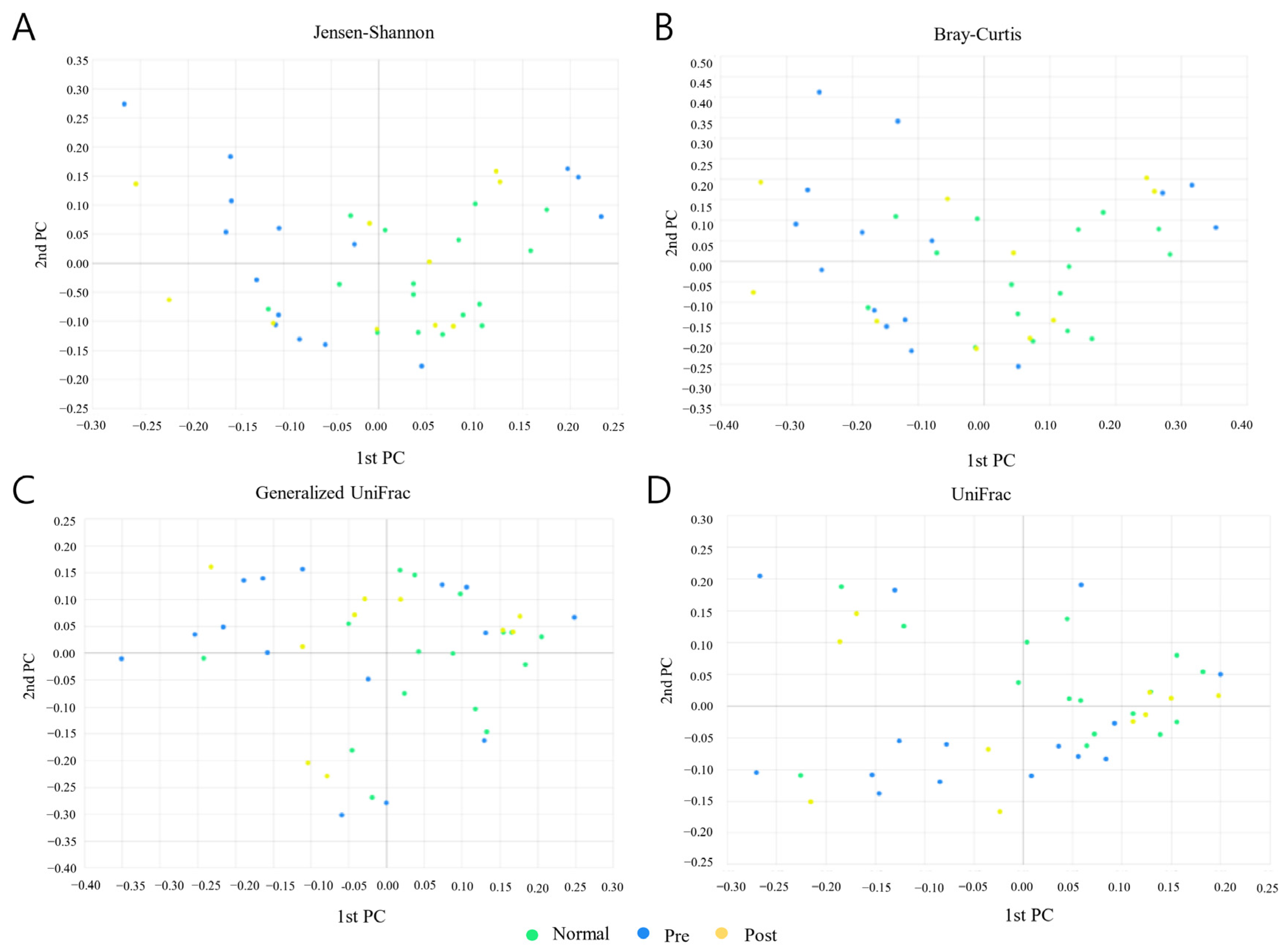
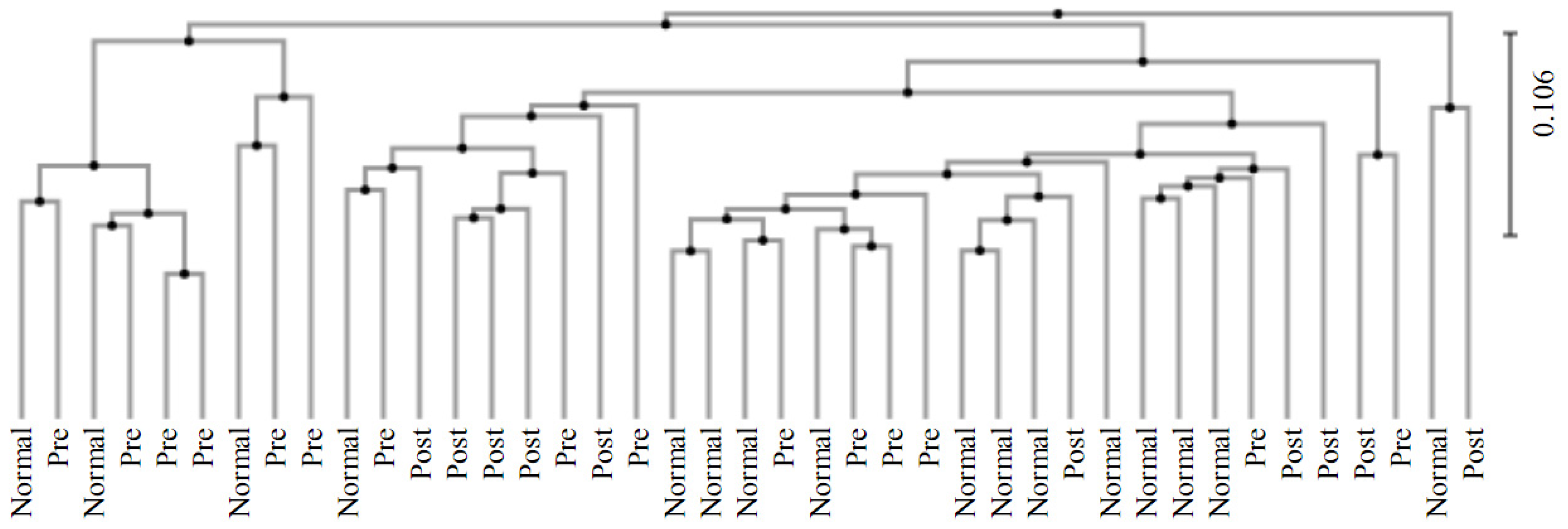
3.4. Beta Diversity Analysis
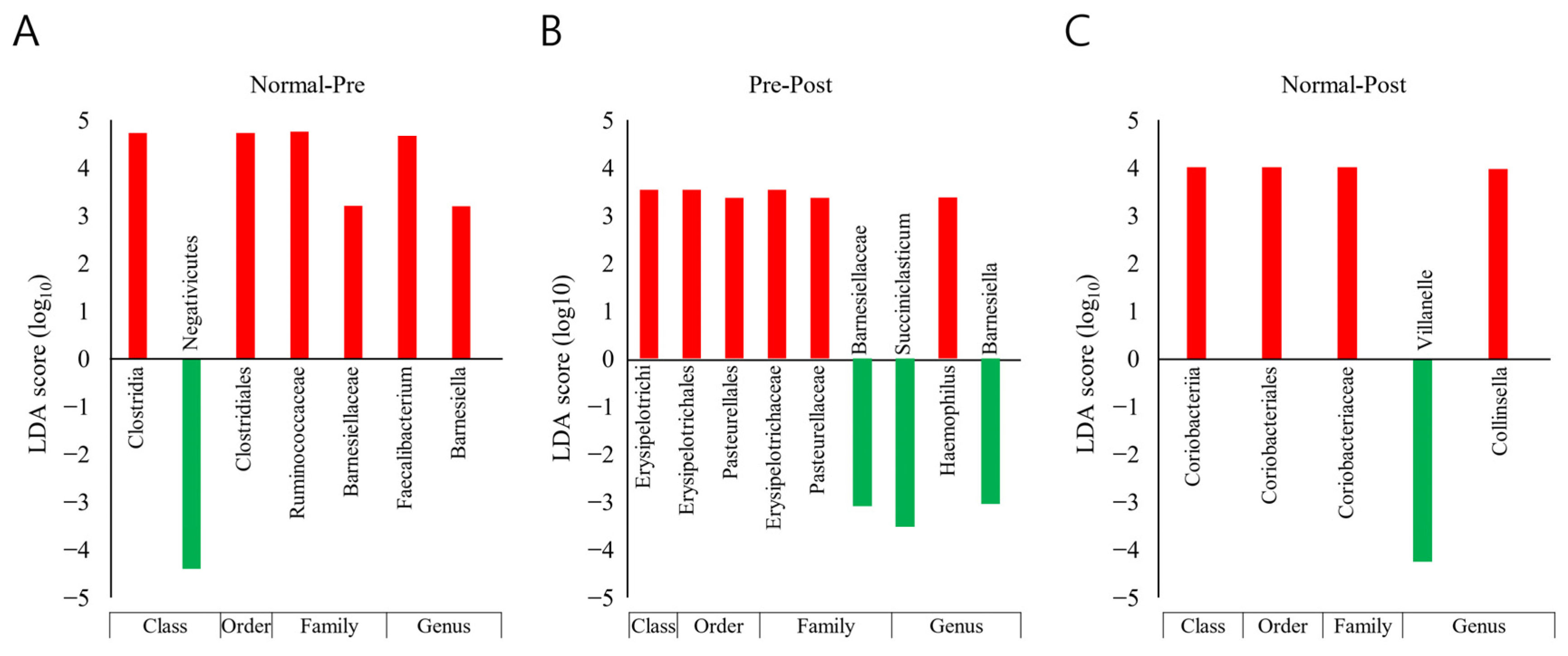
3.5. Taxonomic Biomarker Discovery
3.6. Prediction of Functional Biomarkers
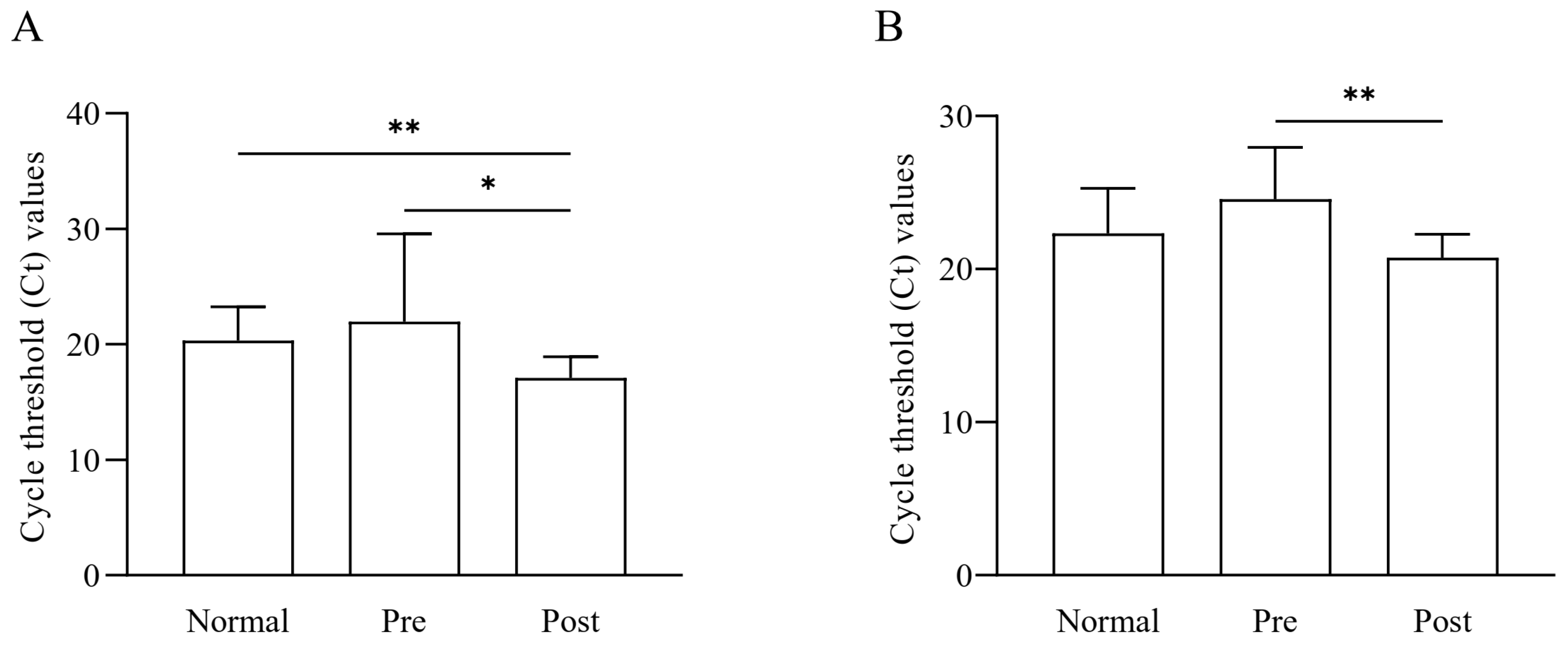
3.7. Quantification of Faecalibacterium Based on qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, S.F. Iron Deficiency Anemia. Nutr. Clin. Pract. 2008, 23, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Zurawski, D.V.; McLendon, M.K. Monoclonal Antibodies as an Antibacterial Approach Against Bacterial Pathogens. Antibiotics 2020, 9, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, M.; Simonetti, M.; Marconi, E.; Brignoli, O.; Cancian, M.; Masotti, A.; Pegoraro, V.; Heiman, F.; Cricelli, C.; Lapi, F. Gender differences in determinants of iron-deficiency anemia: A population-based study conducted in four European countries. Ann. Hematol. 2019, 98, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.B.; Arnipalli, S.R.; Mehta, P.; Carrau, S.; Ziouzenkova, O. Iron Deficiency Anemia: Efficacy and Limitations of Nutritional and Comprehensive Mitigation Strategies. Nutrients 2022, 14, 2976. [Google Scholar] [CrossRef]
- Chen, H.; Wu, W.; Tang, S.; Fu, R.; Gong, X.; Hou, H.; Xu, J. Altered fecal microbial and metabolic profile reveals potential mechanisms underlying iron deficiency anemia in pregnant women in China. Bosn. J. Basic Med. Sci. 2022, 22, 923–933. [Google Scholar] [CrossRef]
- DeLoughery, T.G. Iron Deficiency Anemia. Med. Clin. N. Am. 2017, 101, 319–332. [Google Scholar] [CrossRef]
- McArdle, H.J.; Gambling, L.; Kennedy, C. Iron deficiency during pregnancy: The consequences for placental function and fetal outcome. Proc. Nutr. Soc. 2014, 73, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Cassat, J.E.; Skaar, E.P. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Goodnough, L.T.; Nemeth, E.; Ganz, T. Detection, evaluation, and management of iron-restricted erythropoiesis. Blood 2010, 116, 4754–4761. [Google Scholar] [CrossRef]
- Stevens, G.A.; Paciorek, C.J.; Flores-Urrutia, M.C.; Borghi, E.; Namaste, S.; Wirth, J.P.; Suchdev, P.S.; Ezzati, M.; Rohner, F.; Flaxman, S.R.; et al. National, regional, and global estimates of anaemia by severity in women and children for 2000–19: A pooled analysis of population-representative data. Lancet Glob. Health 2022, 10, e627–e639. [Google Scholar] [CrossRef] [PubMed]
- Pavord, S.; Myers, B.; Robinson, S.; Allard, S.; Strong, J.; Oppenheimer, C.; British Committee for Standards in Haematology. UK guidelines on the management of iron deficiency in pregnancy. Br. J. Haematol. 2012, 156, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.L.; O’Dwyer, N.J.; Parker, H.M.; Donges, C.E.; Cheng, H.L.; Steinbeck, K.S.; Cox, E.P.; Franklin, J.L.; Garg, M.L.; Rooney, K.B.; et al. Iron Deficiency Anemia, Not Iron Deficiency, Is Associated with Reduced Attention in Healthy Young Women. Nutrients 2017, 9, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chueh, H.W.; Jung, H.L.; Shim, Y.J.; Choi, H.S.; Han, J.Y.; on the behalf of the Red Blood Cell Disorder Working Party of The Korean Society of Hematology. High anemia prevalence in Korean older adults, an advent healthcare problem: 2007–2016 KNHANES. BMC Geriatr. 2020, 20, 509. [Google Scholar] [CrossRef]
- Adamu, A.L.; Crampin, A.; Kayuni, N.; Amberbir, A.; Koole, O.; Phiri, A.; Nyirenda, M.; Fine, P. Prevalence and risk factors for anemia severity and type in Malawian men and women: Urban and rural differences. Popul. Health Metr. 2017, 15, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO World Health Organization. Anaemia in Women and Children; WHO: Geneva, Switzerland, 2021. [Google Scholar]
- Rahman, M.M.; Abe, S.K.; Rahman, M.S.; Kanda, M.; Narita, S.; Bilano, V.; Ota, E.; Gilmour, S.; Shibuya, K. Maternal anemia and risk of adverse birth and health outcomes in low- and middle-income countries: Systematic review and meta-analysis1,2. Am. J. Clin. Nutr. 2016, 103, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Milman, N.; Taylor, C.L.; Merkel, J.; Brannon, P.M. Iron status in pregnant women and women of reproductive age in Europe. Am. J. Clin. Nutr. 2017, 106, 1655S–1662S. [Google Scholar] [CrossRef] [Green Version]
- Levi, M.; Rosselli, M.; Simonetti, M.; Brignoli, O.; Cancian, M.; Masotti, A.; Pegoraro, V.; Cataldo, N.; Heiman, F.; Chelo, M.; et al. Epidemiology of iron deficiency anaemia in four European countries: A population-based study in primary care. Eur. J. Haematol. 2016, 97, 583–593. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, E.; Marley, A.; Samaan, M.A.; Brookes, M.J. Iron deficiency anaemia: Pathophysiology, assessment, practical management. BMJ Open Gastroenterol. 2022, 9, e000759. [Google Scholar] [CrossRef]
- Zakrzewski, M.; Wilkins, S.J.; Helman, S.L.; Brilli, E.; Tarantino, G.; Anderson, G.J.; Frazer, D.M. Supplementation with Sucrosomial (R) iron leads to favourable changes in the intestinal microbiome when compared to ferrous sulfate in mice. Biometals 2022, 35, 27–38. [Google Scholar] [CrossRef]
- Soriano-Lerma, A.; Garcia-Burgos, M.; Alferez, M.J.M.; Perez-Carrasco, V.; Sanchez-Martin, V.; Linde-Rodriguez, A.; Ortiz-Gonzalez, M.; Soriano, M.; Garcia-Salcedo, J.A.; Lopez-Aliaga, I. Gut microbiome-short-chain fatty acids interplay in the context of iron deficiency anaemia. Eur. J. Nutr. 2022, 61, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, W.; Luo, Y. Oral and intravenous iron treatment alter the gut microbiome differentially in dialysis patients. Int. Urol. Nephrol. 2022, 61, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Wimbley, T.D.; Graham, D.Y. Diagnosis and management of iron deficiency anemia in the 21st century. Ther. Adv. Gastroenterol. 2011, 4, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Human Microbiome Project—Core Microbiome Sampling Protocol A; HMP Initiative; National Institutes of Health (NIH): Bethesda, MD, USA, 2010.
- Rickham, P.P. Human experimentation. Code of ethics of the world medical association. Declaration of Helsinki. Br. Med. J. 1964, 2, 177. [Google Scholar] [PubMed] [Green Version]
- Kim, S.; Seo, H.; Rahim, M.A.; Tajdozian, H.; Kim, Y.-S.; Song, H.-Y. Characteristics of Vaginal Microbiome in Women with Pelvic Inflammatory Disease in Korea. Pol. J. Microbiol. 2021, 70, 345–357. [Google Scholar] [CrossRef]
- Myers, E.W.; Miller, W. Optimal alignments in linear space. Bioinformatics 1988, 4, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, T.J.; Eddy, S.R. nhmmer: DNA homology search with profile HMMs. Bioinformatics 2013, 29, 2487–2489. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Shokralla, S.; Porter, T.M.; Gibson, J.F.; Dobosz, R.; Janzen, D.H.; Hallwachs, W.; Golding, G.B.; Hajibabaei, M.J.S.R. Massively parallel multiplex DNA sequencing for specimen identification using an Illumina MiSeq platform. Sci. Rep. 2015, 5, 9687. [Google Scholar] [CrossRef]
- Chao, A.; Lee, S.M. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Chao, A.J.B. Estimating the population size for capture-recapture data with unequal catchability. Biometrics 1987, 43, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Overton, W.S.J.E. Robust estimation of population size when capture probabilities vary among animals. Ecology 1979, 60, 927–936. [Google Scholar] [CrossRef]
- Chao, A.; Shen, T.-J. Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample. Environ. Ecol. Stat. 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring biological diversity. Curr. Biol. 2021, 31, R1174–R1177. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Lin, J. Divergence measures based on the Shannon entropy. IEEE Trans. Inform. Theory 1991, 37, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Beals, E.W. Bray-Curtis ordination: An effective strategy for analysis of multivariate ecological data. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1984; Volume 14, pp. 1–55. [Google Scholar]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H.J.B. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef] [Green Version]
- Hamady, M.; Lozupone, C.; Knight, R. Fast UniFrac: Facilitating high-throughput phylogenetic analyses of microbial communities including analysis of pyrosequencing and PhyloChip data. ISME J. 2010, 4, 17–27. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Ye, Y.; Doak, T.G. A parsimony approach to biological pathway reconstruction/inference for genomes and metagenomes. PLoS Comput. Biol. 2009, 5, e1000465. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.J. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Fitzgerald, C.B.; Shkoporov, A.N.; Sutton, T.D.S.; Chaplin, A.V.; Velayudhan, V.; Ross, R.P.; Hill, C. Comparative analysis of Faecalibacterium prausnitzii genomes shows a high level of genome plasticity and warrants separation into new species-level taxa. BMC Genom. 2018, 19, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Yampara-Iquise, H.; Jones, J.E.; Andrew Carson, C. Development of Faecalibacterium 16S rRNA gene marker for identification of human faeces. J. Appl. Microbiol. 2009, 106, 634–641. [Google Scholar] [CrossRef]
- Safiri, S.; Kolahi, A.A.; Noori, M.; Nejadghaderi, S.A.; Karamzad, N.; Bragazzi, N.L.; Sullman, M.J.M.; Abdollahi, M.; Collins, G.S.; Kaufman, J.S.; et al. Burden of anemia and its underlying causes in 204 countries and territories, 1990–2019: Results from the Global Burden of Disease Study 2019. J. Hematol. Oncol. 2021, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Muleviciene, A.; D’Amico, F.; Turroni, S.; Candela, M.; Jankauskiene, A. Iron deficiency anemia-related gut microbiota dysbiosis in infants and young children: A pilot study. Acta Microbiol. Immunol. Hung. 2018, 65, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.B.; Chassard, C.; Rohner, F.; Goran, E.K.N.; Nindjin, C.; Dostal, A.; Utzinger, J.; Ghattas, H.; Lacroix, C.; Hurrell, R.F. The effects of iron fortification on the gut microbiota in African children a randomized controlled trial in Cote d’Ivoire. Am. J. Clin. Nutr. 2010, 92, 1406–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquel, S.; Martin, R.; Rossi, O.; Bermudez-Humaran, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef]
- Ferreira-Halder, C.V.; Faria, A.V.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pr. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Liang, F.; Guo, R.; Zhu, C.; Zhao, X.; Wang, X.; Liu, F.; Jiang, M.; Liang, Q.; Zeng, S.; et al. Gut Microbiota Signatures in Gestational Anemia. Front. Cell Infect. Microbiol. 2021, 11, 549678. [Google Scholar] [CrossRef] [PubMed]
- Deschemin, J.C.; Noordine, M.L.; Remot, A.; Willemetz, A.; Afif, C.; Canonne-Hergaux, F.; Langella, P.; Karim, Z.; Vaulont, S.; Thomas, M.; et al. The microbiota shifts the iron sensing of intestinal cells. FASEB J. 2016, 30, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut-heart axis in atherosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef]
- Wessling-Resnick, M. Iron homeostasis and the inflammatory response. Annu. Rev. Nutr. 2010, 30, 105–122. [Google Scholar] [CrossRef] [Green Version]
- Layer, G. Heme biosynthesis in prokaryotes. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118861. [Google Scholar] [CrossRef]
- Dailey, H.A.; Dailey, T.A.; Gerdes, S.; Jahn, D.; Jahn, M.; O’Brian, M.R.; Warren, M.J. Prokaryotic Heme Biosynthesis: Multiple Pathways to a Common Essential Product. Microbiol. Mol. Biol. Rev. 2017, 81, e00048-16. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal (n = 16) | IDA-pre. Tx. (n = 15) | IDA post. Tx. (n = 10) | Normal Reference Value | |
|---|---|---|---|---|
| Age (years) | 29.25 ± 7.1 | 37.13 ± 7.92 | ||
| Height (cm) | 161.59 ± 3.85 | 161.31 ± 3.48 | ||
| WBC (/µL) | 6793.75 ± 1614.3 | 5046.67 ± 1374.7 | 5440 ± 1734.74 | 4000–10,000 |
| Hemoglobin (g/dl) | 13.43 ± 0.77 | 8.57 ± 1.26 * | 13.31 ± 0.75 | 12–16 |
| Hematocrit (%) | 40.11 ± 1.93 | 28.11 ± 3.33 * | 39.85 ± 2.54 | 36–48 |
| MCV (fl) | 92.52 ± 3.82 | 67.93 ± 7.01 * | 89.19 ± 5.72 | 86–102 |
| RDW (%) | 14.59 ± 4.61 | 19.44 ± 3.45 * | 13.91 ± 1.14 | 11.5–14.5 |
| Platelet (103/µL) | 247.94 ± 45.86 | 344.93 ± 95.67 | 297.5 ± 83.64 | 130–450 |
| Ferritin (ng/mL) | 49.55 (32.42, 75.75) | 2.31 ± 1.4 * | 60.87 ± 77.28 | 5–204 |
| Class | Norl (%) | PRE (%) | Post (%) | Order | Nor (%) | PRE (%) | Post (%) | Family | Nor (%) | PRE (%) | Post (%) | Genus | Nor (%) | PRE (%) | Post (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clostridia *, a | 45.6 | 33.8 | 42.2 | Clostridiales *, a | 45.6 | 33.8 | 42.2 | Ruminococcaceae **, a | 31.2 | 19.6 | 29.1 | Faecalibacterium **, a | 15.3 | 7.5 | 14.9 |
| Coriobacteriia *, c | 3.5 | 2.0 | 1.5 | Coriobacteriales *, c | 3.5 | 2.0 | 1.5 | Coriobacteriaceae *, c | 3.5 | 2.0 | 1.5 | Collinsella *, c | 2.6 | 1.2 | 0.0 |
| Negativicutes *, a | 4.5 | 8.8 | 8.4 | Veillonella *, c | 0.4 | 2.0 | 3.9 | ||||||||
| Erysipelotrichi *, b | 1.2 | 1.4 | 0.0 | Erysipelotrichales *, b | 1.2 | 1.4 | 0.0 | Erysipelotrichaceae *, b | 1.2 | 1.4 | 0.0 | ||||
| Betaproteobacteria *, b | 1.5 | 2.2 | 1.5 |
| Index | Normal–Pre | Normal–Post | Pre–Post |
|---|---|---|---|
| Jenson–Shannon | * (p = 0.037) | N.S. (p = 0.672) | N.S. (p = 1.000) |
| Bray–Curtis | * (p = 0.017) | N.S. (p = 0.282) | N.S. (p = 0.929) |
| Generalized UniFrac | * (p = 0.045) | N.S. (p = 0.430) | N.S. (p = 0.853) |
| UniFrac | N.S. (p = 0.062) | N.S. (p = 0.329) | N.S. (p = 0.594) |
| Ortholog | Definition | LDA Effect Size | p-Value | Normal (%) | Pre (%) | Post (%) | |
|---|---|---|---|---|---|---|---|
| Orthology | K03205 | Type IV secretion system protein VirD4 | 2.44 | 0.0288 | 0.16 | 0.10 | 0.15 |
| K02315 | DNA replication protein DnaC | 2.36 | 0.0209 | 0.14 | 0.09 | 0.14 | |
| K03169 | DNA topoisomerase III | 2.34 | 0.0358 | 0.16 | 0.12 | 0.15 | |
| K03496 | Chromosome partitioning protein | 2.33 | 0.0098 | 0.21 | 0.17 | 0.21 | |
| K06919 | Putative DNA primase/helicase | 2.32 | 0.0419 | 0.15 | 0.12 | 0.16 | |
| K02316 | DNA primase | 2.02 | 0.0493 | 0.11 | 0.10 | 0.12 | |
| Module (PICRUSt) | M00051 | Uridine monophosphate biosynthesis, glutamine (+PRPP) => UMP | 2.48 | 0.0155 | 0.89 | 0.86 | 0.92 |
| M00121 | Heme biosynthesis, glutamate => heme | 2.42 | 0.0144 | 0.33 | 0.39 | 0.35 | |
| Module (MinPath) | M00854 | Glycogen biosynthesis, glucose-1P => glycogen/starch | 2.59 | 0.0096 | 0.33 | 0.24 | 0.32 |
| M00086 | beta-Oxidation, acyl-CoA synthesis | 2.57 | 0.0132 | 0.34 | 0.30 | 0.37 | |
| Pathway (PICRUSt) | ko04142 | Lysosome | 2.45 | 0.0124 | 0.29 | 0.35 | 0.29 |
| Pathway (MinPath) | ko04910 | Insulin signaling pathway | 2.85 | 0.0110 | 0.34 | 0.49 | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, H.; Yoon, S.Y.; ul-Haq, A.; Jo, S.; Kim, S.; Rahim, M.A.; Park, H.-A.; Ghorbanian, F.; Kim, M.J.; Lee, M.-Y.; et al. The Effects of Iron Deficiency on the Gut Microbiota in Women of Childbearing Age. Nutrients 2023, 15, 691. https://doi.org/10.3390/nu15030691
Seo H, Yoon SY, ul-Haq A, Jo S, Kim S, Rahim MA, Park H-A, Ghorbanian F, Kim MJ, Lee M-Y, et al. The Effects of Iron Deficiency on the Gut Microbiota in Women of Childbearing Age. Nutrients. 2023; 15(3):691. https://doi.org/10.3390/nu15030691
Chicago/Turabian StyleSeo, Hoonhee, Seug Yun Yoon, Asad ul-Haq, Sujin Jo, Sukyung Kim, Md Abdur Rahim, Hyun-A Park, Fatemeh Ghorbanian, Min Jung Kim, Min-Young Lee, and et al. 2023. "The Effects of Iron Deficiency on the Gut Microbiota in Women of Childbearing Age" Nutrients 15, no. 3: 691. https://doi.org/10.3390/nu15030691