
Mechanistic Action of Cell Cycle Arrest and Intrinsic Apoptosis via Inhibiting Akt/mTOR and Activation of p38-MAPK Signaling Pathways in Hep3B Liver Cancer Cells by Prunetrin—A Flavonoid with Therapeutic Potential

 , , , , , and
, , , , , and 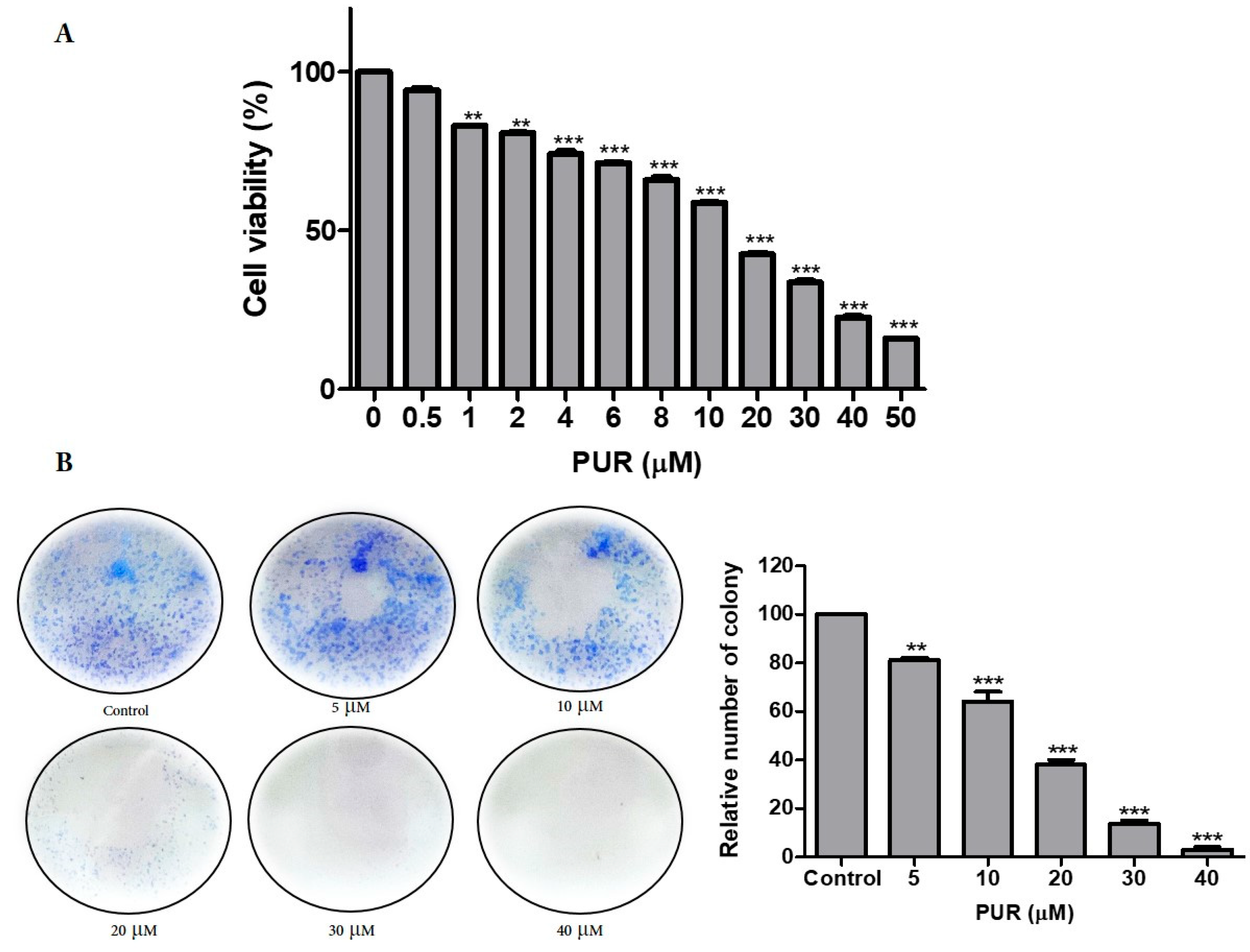{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line and Maintenance
2.2. Cytotoxicity Test
2.3. Colony Formation Assay
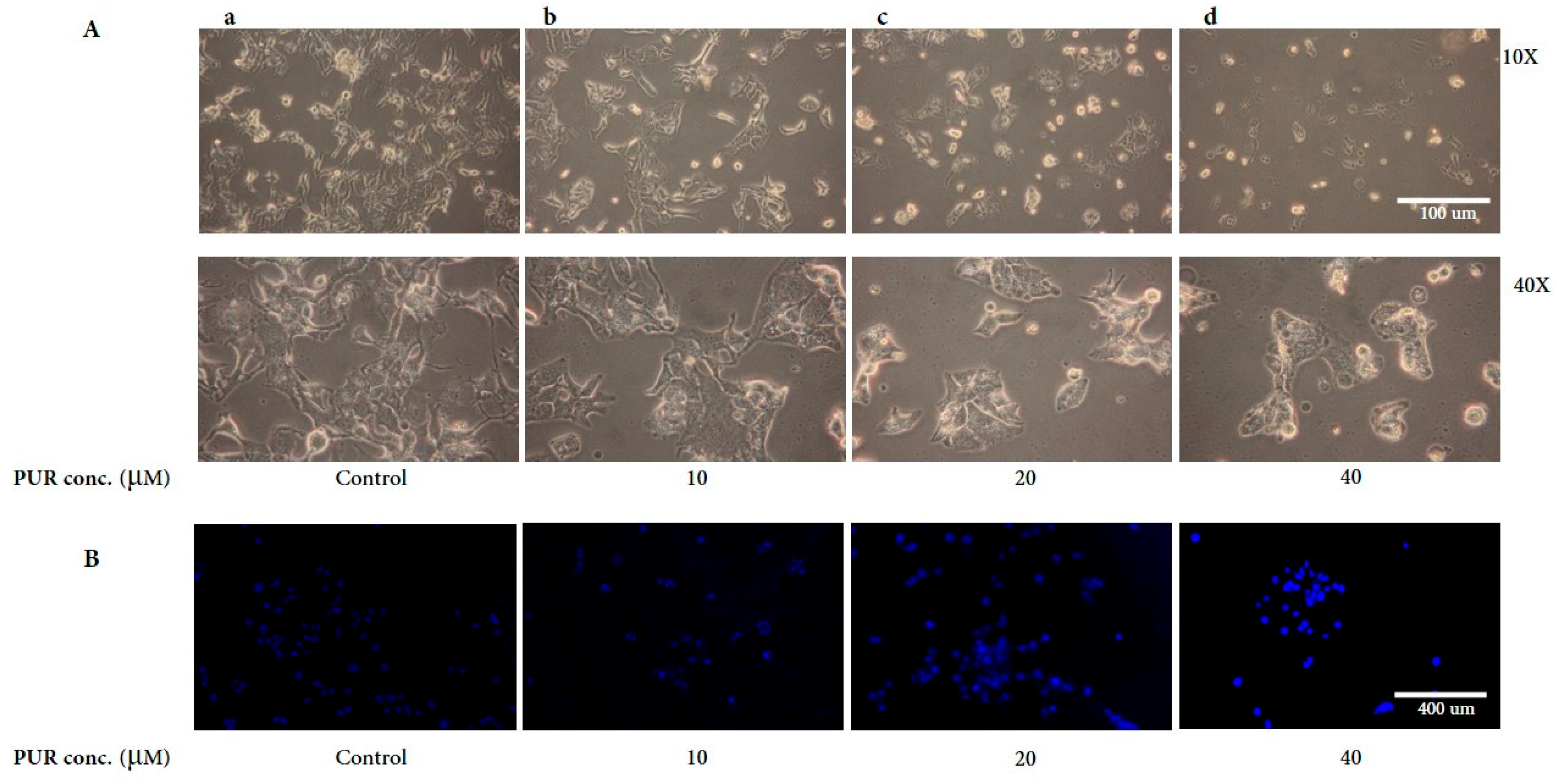
2.4. DAPI Staining
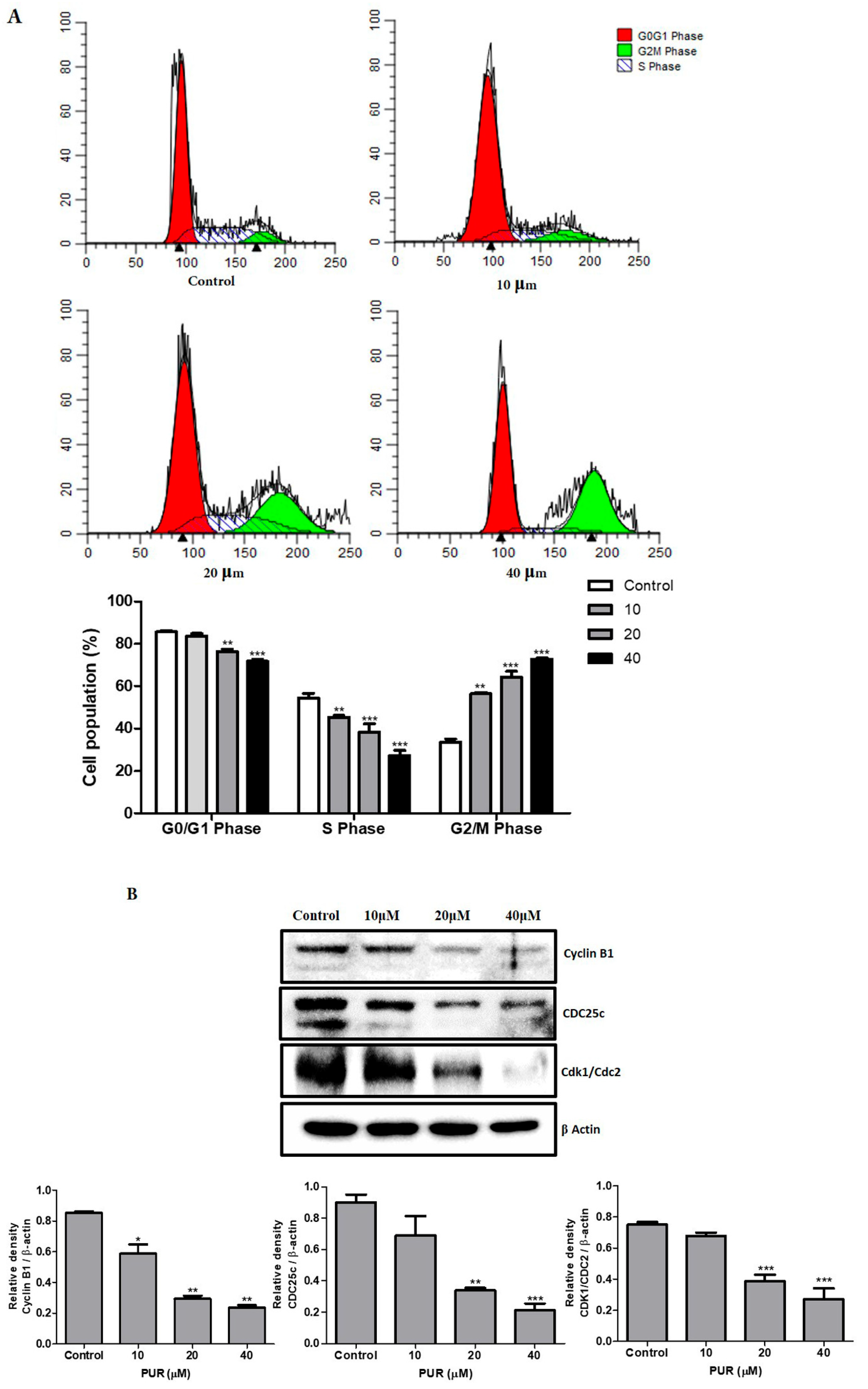
2.5. Cell Cycle Analysis
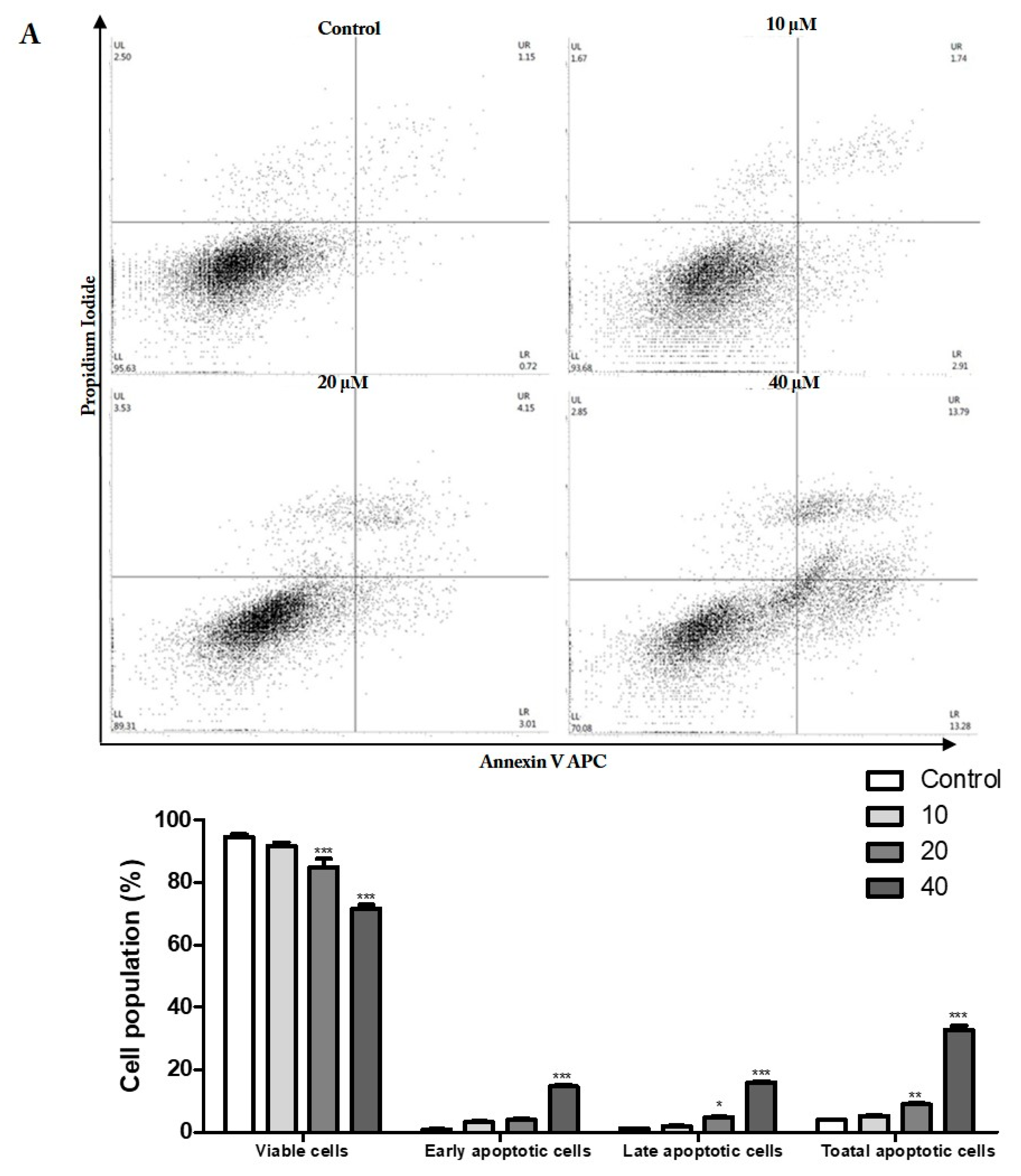
2.6. Double Staining
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Prunetrin Induces Cytotoxicity in Hep3B Cells
3.2. Morphological Assessment of Hep3B Cells Treated with PUR
3.3. Prunetrin Induces Cell Cycle Arrest in Hep3B Cells
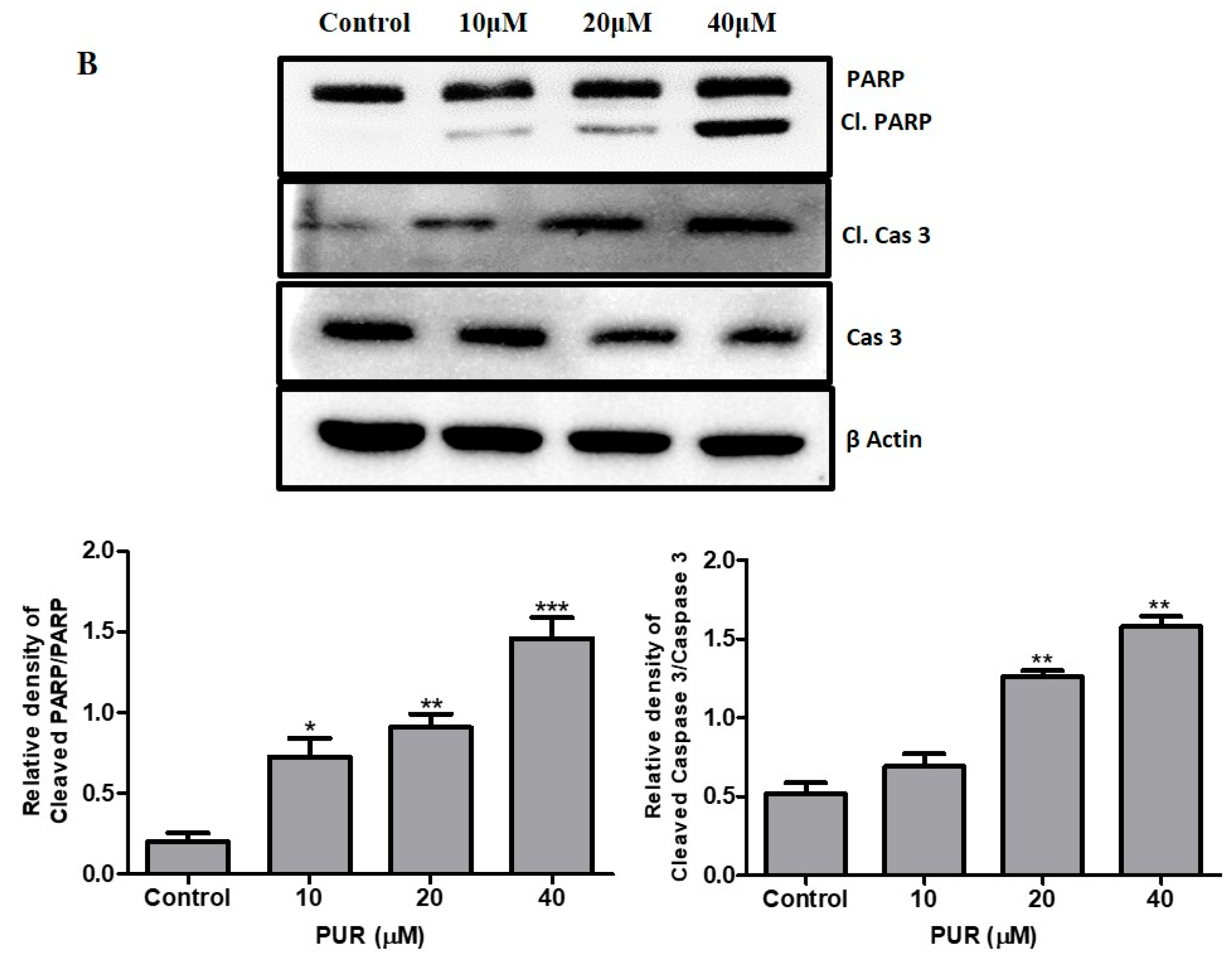
3.4. Prunetrin Induces Apoptosis and Increases the Expression of Apoptotic Hallmark Proteins
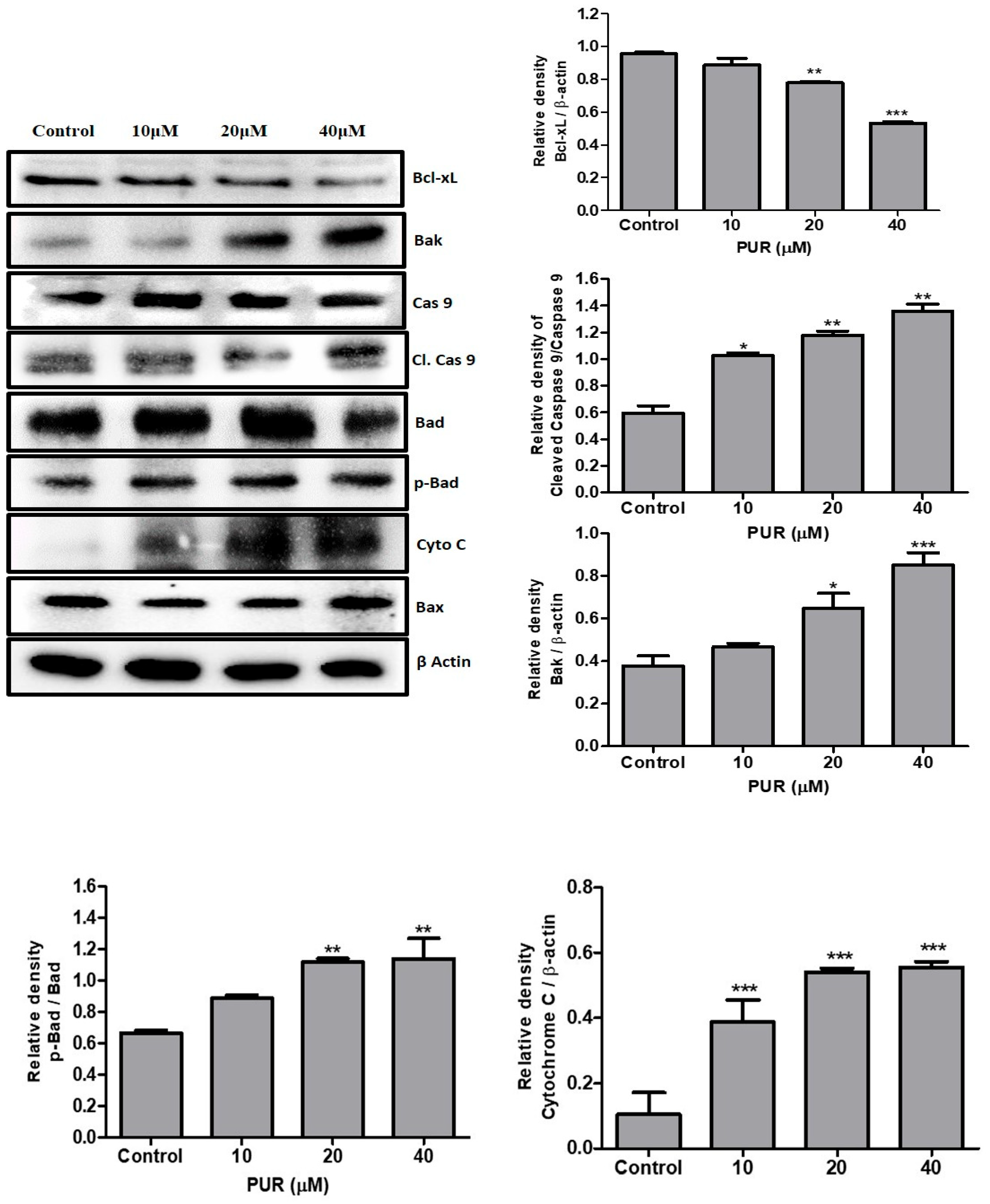
3.5. Prunetrin Activates an Intrinsic Apoptotic Pathway in Hep3B Cells
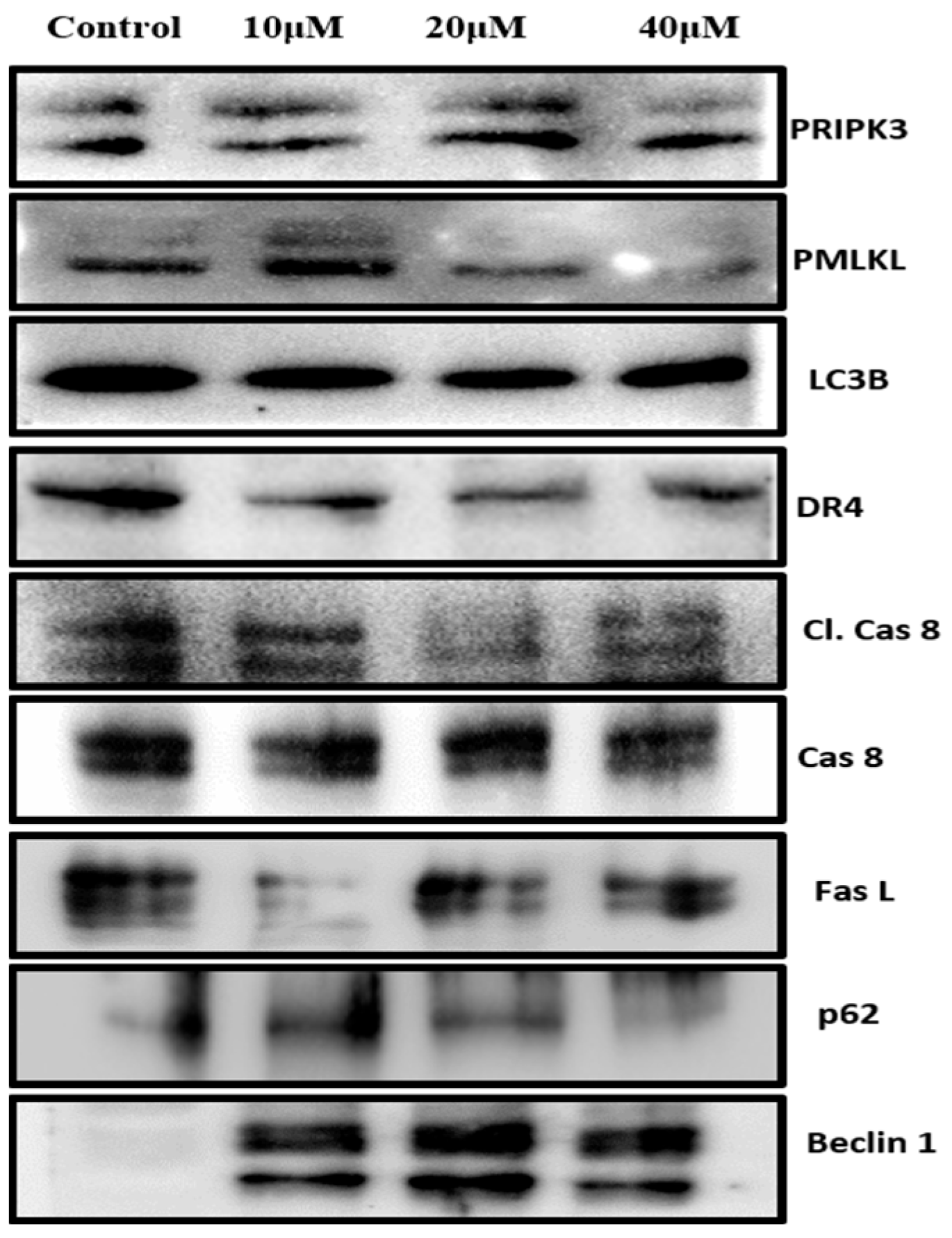
3.6. Prunetrin Does Not Induce Extrinsic Apoptosis and Other Cell Death Modes in Hep3B Cells
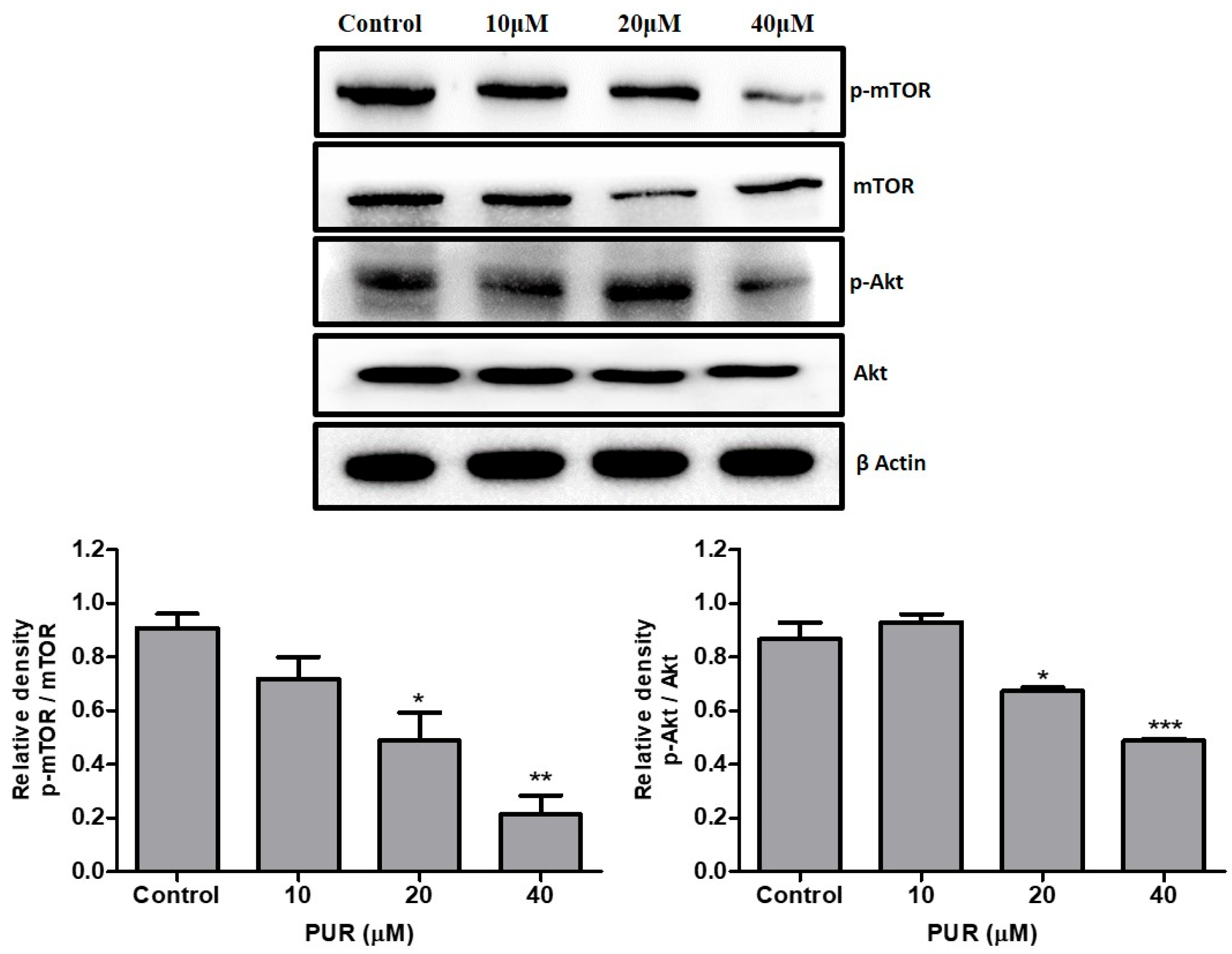
3.7. Prunetrin Inhibits AKT/mTOR Signaling Pathway
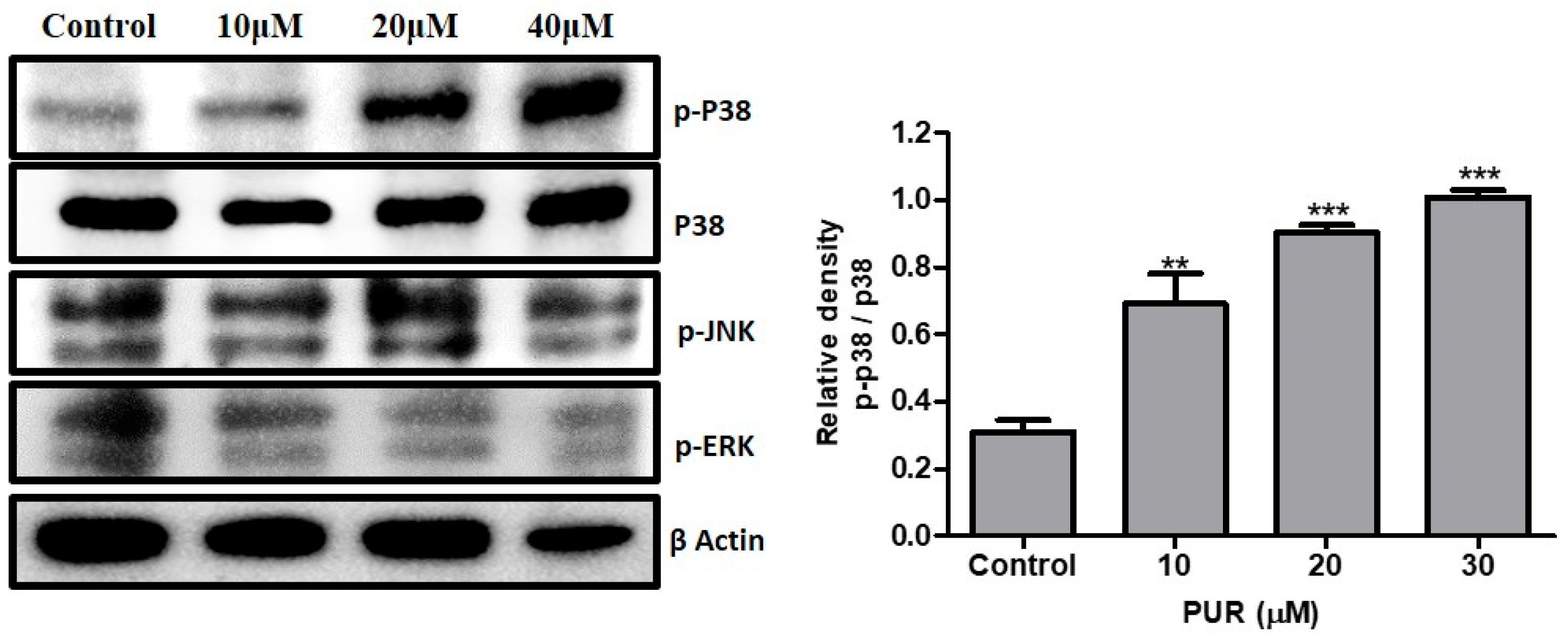
3.8. Effect of Prunetrin on MAPK Pathway
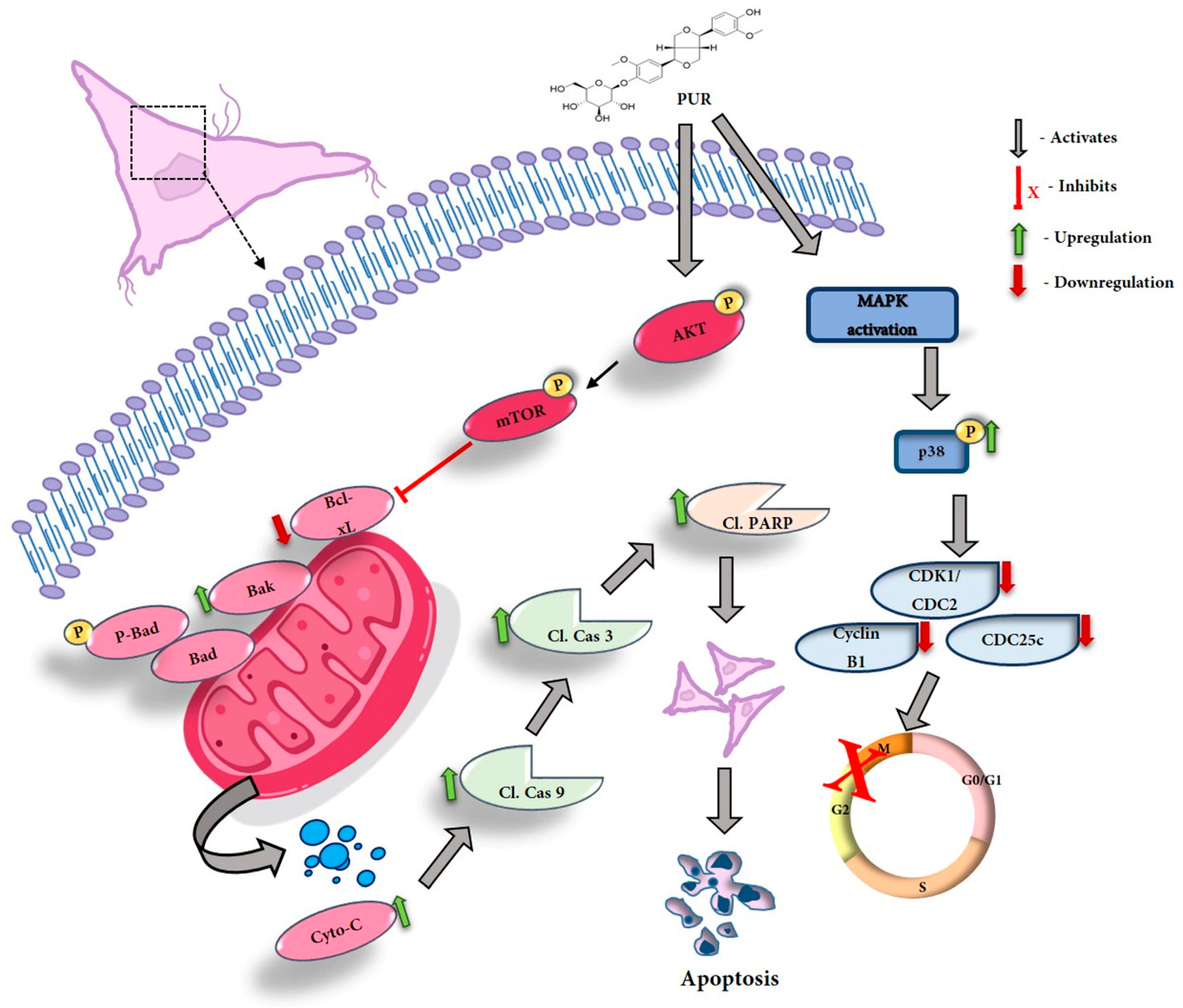
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Angeli, P.; Bernardi, M.; Villanueva, C.; Francoz, C.; Mookerjee, R.P.; Trebicka, J.; Krag, A.; Laleman, W.; Gines, P. EASL Clinical Practice Guidelines for the management of patients with decompensated cirrhosis. J. Hepatol. 2018, 69, 406–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deo, S.V.S.; Sharma, J.; Kumar, S. GLOBOCAN 2020 Report on Global Cancer Burden: Challenges and Opportunities for Surgical Oncologists. Ann. Surg. Oncol. 2022, 29, 6497–6500. [Google Scholar] [CrossRef]
- Sarveazad, A.; Agah, S.; Babahajian, A.; Amini, N.; Bahardoust, M. Predictors of 5 year survival rate in hepatocellular carcinoma patients. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2019, 24, 86. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Islami, F.; Guerra, C.E.; Minihan, A.; Yabroff, K.R.; Fedewa, S.A.; Sloan, K.; Wiedt, T.L.; Thomson, B.; Siegel, R.L.; Nargis, N. American Cancer Society’s report on the status of cancer disparities in the United States, 2021. CA Cancer J. Clin. 2022, 72, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Cassinotto, C.; Nogue, E.; Durand, Q.; Panaro, F.; Assenat, E.; Dohan, A.; Malafaye, N.; Guiu, B.; Molinari, N. Life expectancy of patients with hepatocellular carcinoma according to the upfront treatment: A nationwide analysis. Diagn. Interv. Imaging 2023, 104, 192–199. [Google Scholar] [CrossRef]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [Green Version]
- Abadi, A.J.; Mirzaei, S.; Mahabady, M.K.; Hashemi, F.; Zabolian, A.; Hashemi, F.; Raee, P.; Aghamiri, S.; Ashrafizadeh, M.; Aref, A.R.; et al. Curcumin and its derivatives in cancer therapy: Potentiating antitumor activity of cisplatin and reducing side effects. Phytother. Res. 2022, 36, 189–213. [Google Scholar] [CrossRef]
- Taheri, M.; Shoorei, H.; Anamag, F.T.; Ghafouri-Fard, S.; Dinger, M.E. LncRNAs and miRNAs participate in determination of sensitivity of cancer cells to cisplatin. Exp. Mol. Pathol. 2021, 123, 104602. [Google Scholar] [CrossRef]
- Wu, C.-C.; Bratton, S.B.; Pan, Z.; Niu, Y.; Liang, Y.; Zhang, X.; Dong, M.; Zhang, S.; Li, T.; Zhang, Y.; et al. Regulation of the Intrinsic Apoptosis Pathway by Reactive Oxygen Species. Antioxidants Redox Signal. 2013, 19, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Ghobrial, I.M.; Witzig, T.E.; Adjei, A.A. Targeting Apoptosis Pathways in Cancer Therapy. CA Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.M.; Flecha, A.R.; Keller, P.; Ram, A.; Karnak, D.; Galbán, S.; Galbán, C.J.; Ross, B.D.; Lawrence, T.S.; Rehemtulla, A.; et al. Cotargeting MAPK and PI3K Signaling with Concurrent Radiotherapy as a Strategy for the Treatment of Pancreatic Cancer. Mol. Cancer Ther. 2012, 11, 1193–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O. A com-prehensive review on MAPK: A promising therapeutic target in cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruman, D.A.; Rommel, C. PI3K and cancer: Lessons, challenges and opportunities. Nat. Rev. Drug Discov. 2014, 13, 140–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abusaliya, A.; Ha, S.E.; Bhosale, P.B.; Kim, H.H.; Park, M.Y.; Vetrivel, P.; Kim, G.S. Glycosidic flavonoids and their potential applications in cancer research: A review. Mol. Cell Toxicol. 2022, 18, 9–16. [Google Scholar] [CrossRef]
- Petrick, J.L.; Steck, S.E.; Bradshaw, P.T.; Trivers, K.F.; Abrahamson, P.E.; Engel, L.S.; He, K.; Chow, W.-H.; Mayne, S.T.; Risch, H.A.; et al. Dietary intake of flavonoids and oesophageal and gastric cancer: Incidence and survival in the United States of America (USA). Br. J. Cancer 2015, 112, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Goodarzi, S.; Tabatabaei, M.J.; Jafari, R.M.; Shemirani, F.; Tavakoli, S.; Mofasseri, M.; Tofighi, Z. Cuminum cyminum fruits as source of luteolin-7-O-glucoside, potent cytotoxic flavonoid against breast cancer cell lines. Nat. Prod. Res. 2018, 34, 1602–1606. [Google Scholar] [CrossRef]
- Naito, R.; Kano, H.; Shimada, T.; Makino, T.; Kadomoto, S.; Iwamoto, H.; Yaegashi, H.; Izumi, K.; Kadono, Y.; Nakata, H.; et al. A new flavonoid derivative exerts antitumor effects against androgen-sensitive to cabazitaxel-resistant prostate cancer cells. Prostate 2021, 81, 295–306. [Google Scholar] [CrossRef]
- Scagliarini, A.; Mathey, A.; Aires, V.; Delmas, D. Xanthohumol, a Prenylated Flavonoid from Hops, Induces DNA Damages in Colorectal Cancer Cells and Sensitizes SW480 Cells to the SN38 Chemotherapeutic Agent. Cells 2020, 9, 932. [Google Scholar] [CrossRef] [Green Version]
- Vetrivel, P.; Kim, S.M.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Senthil, K.; Kim, G.S. Compound Prunetin Induces Cell Death in Gastric Cancer Cell with Potent Anti-Proliferative Properties: In Vitro Assay, Molecular Docking, Dynamics, and ADMET Studies. Biomolecules 2020, 10, 1086. [Google Scholar] [CrossRef]
- Vetrivel, P.; Murugesan, R.; Bhosale, P.B.; Ha, S.E.; Kim, H.H.; Heo, J.D.; Kim, G.S. A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer. Cancers 2021, 13, 1918. [Google Scholar] [CrossRef] [PubMed]
- Abusaliya, A.; Bhosale, P.B.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Jeong, S.H.; Vetrivel, P.; Park, J.-S.; Kim, G.S. Prunetinoside Inhibits Lipopolysaccharide-Provoked Inflammatory Response via Suppressing NF-κB and Activating the JNK-Mediated Signaling Pathway in RAW264. 7 Macrophage Cells. Int. J. Mol. Sci. 2022, 23, 5442. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Wilson, C. Can dietary flavonoids be useful in the personalized treatment of colorectal cancer? World J. Gastrointest. Oncol. 2022, 14, 1115. [Google Scholar] [CrossRef]
- D’Arcy, M.S. A review of biologically active flavonoids as inducers of autophagy and apoptosis in neoplastic cells and as cytoprotective agents in non-neoplastic cells. Cell Biol. Int. 2022, 46, 1179–1195. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, X.; Zhu, X.; Pan, Z.; Hao, W.; Li, D.; Zheng, Q.; Tang, X. Luteolin potentiates low-dose oxaliplatin-induced inhibitory effects on cell proliferation in gastric cancer by inducing G2/M cell cycle arrest and apoptosis. Oncol. Lett. 2022, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Abushawish, K.Y.I.; Soliman, S.S.M.; Giddey, A.D.; Al-Hroub, H.M.; Mousa, M.; Alzoubi, K.H.; El-Huneidi, W.; Abu-Gharbieh, E.; Omar, H.A.; Elgendy, S.M.; et al. Multi-Omics Analysis Revealed a Significant Alteration of Critical Metabolic Pathways Due to Sorafenib-Resistance in Hep3B Cell Lines. Int. J. Mol. Sci. 2022, 23, 11975. [Google Scholar] [CrossRef]
- Ling, J.-Y.; Wang, Q.-L.; Liang, H.-N.; Liu, Q.-B.; Yin, D.-H.; Lin, L. Flavonoid-Rich Extract of Oldenlandia diffusa (Willd.) Roxb. Inhibits Gastric Cancer by Activation of Caspase-Dependent Mitochondrial Apoptosis. Chin. J. Integr. Med. 2023, 29, 213–223. [Google Scholar] [CrossRef]
- Matthews, H.K.; Bertoli, C.; de Bruin, R.A. Cell cycle control in cancer. Nat. Rev. Mol. Cell Biol. 2022, 23, 74–88. [Google Scholar] [CrossRef]
- Liu, J.; Peng, Y.; Wei, W. Cell cycle on the crossroad of tumorigenesis and cancer therapy. Trends Cell Biol. 2022, 32, 30–44. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. Cell cycle, CDKs and cancer: A changing paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef]
- Roskoski, R., Jr. Cyclin-dependent protein serine/threonine kinase inhibitors as anticancer drugs. Pharmacol. Res. 2019, 139, 471–488. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.Y.; Fang, R.C.; Yuan, W.S. Polyphyllin D induces G2/M Cell Cycle Arrest via Dysfunction of Cholesterol Biosynthesis in Liver Cancer Cells. Biomed. Environ. Sci. 2023, 36, 94–98. [Google Scholar]
- Bhosale, P.B.; Abusaliya, A.; Kim, H.H.; Ha, S.E.; Park, M.Y.; Jeong, S.H.; Vetrivel, P.; Heo, J.D.; Kim, J.-A.; Won, C.k. Apigetrin Promotes TNFα-Induced Apoptosis, Necroptosis, G2/M Phase Cell Cycle Arrest, and ROS Generation through Inhibition of NF-κB Pathway in Hep3B Liver Cancer Cells. Cells 2022, 11, 2734. [Google Scholar] [CrossRef] [PubMed]
- Pucci, B.; Kasten, M.; Giordano, A. Cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of poly (ADP-ribose) polymerase (PARP) cleavage in apoptosis: Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef] [Green Version]
- Jan, R.; Chaudhry, G.-E. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Basu, A. The interplay between apoptosis and cellular senescence: Bcl-2 family proteins as targets for cancer therapy. Pharmacol. Ther. 2021, 230, 107943. [Google Scholar] [CrossRef]
- Xia, W.; Gong, E.-s.; Lin, Y.; Zheng, B.; Yang, W.; Li, T.; Zhang, S.; Li, P.; Liu, R.-h. Wild pink bayberry free phenolic extract induces mitochondria-dependent apoptosis and G0/G1 cell cycle arrest through p38/MAPK and PI3K/Akt pathway in MDA-MB-231 cancer cells. Food Sci. Hum. Wellness 2023, 12, 1510–1518. [Google Scholar] [CrossRef]
- Ow, Y.-L.P.; Green, D.R.; Hao, Z.; Mak, T.W. Cytochrome c: Functions beyond respiration. Nat. Rev. Mol. Cell Biol. 2008, 9, 532–542. [Google Scholar] [CrossRef]
- Cai, J.; Yang, J.; Jones, D. Mitochondrial control of apoptosis: The role of cytochrome c. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 1998, 1366, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Li, W.; Miao, H.; Yao, J.; Li, Z.; Wei, L.; Zhao, L.; Guo, Q. LW-214, a newly synthesized flavonoid, induces intrinsic apoptosis pathway by down-regulating Trx-1 in MCF-7 human breast cells. Biochem. Pharmacol. 2014, 87, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.-P.; Yan, Y.; Xia, J.; Zhang, S.; Wang, M.; Chen, J.; Xu, Y. A phenylacetaldehyde-flavonoid adduct, 8-C-(E-phenylethenyl)-norartocarpetin, exhibits intrinsic apoptosis and MAPK pathways-related anticancer potential on HepG2, SMMC-7721 and QGY-7703. Food Chem. 2016, 197, 1085–1092. [Google Scholar] [CrossRef]
- Kim, J.-H.; Kang, J.W.; Kim, M.S.; Bak, Y.; Park, Y.S.; Jung, K.-Y.; Lim, Y.H.; Yoon, D.-Y. The apoptotic effects of the flavonoid N101-2 in human cervical cancer cells. Toxicol. Vitr. 2012, 26, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Han, J.; Hui, L. MAPK signaling in inflammation-associated cancer development. Protein Cell 2010, 1, 218–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santarpia, L.; Lippman, S.M.; El-Naggar, A.K. Targeting the MAPK-RAS-RAF signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 103–119. [Google Scholar] [CrossRef] [Green Version]
- Anjum, J.; Mitra, S.; Das, R.; Alam, R.; Mojumder, A.; Bin Emran, T.; Islam, F.; Rauf, A.; Hossain, J.; Aljohani, A.S.; et al. A renewed concept on the MAPK signaling pathway in cancers: Polyphenols as a choice of therapeutics. Pharmacol. Res. 2022, 184, 106398. [Google Scholar] [CrossRef]
- Kang, N.; Wang, M.-M.; Wang, Y.-H.; Zhang, Z.-N.; Cao, H.-R.; Lv, Y.-H.; Yang, Y.; Fan, P.-H.; Qiu, F.; Gao, X.-M. Tetrahydrocurcumin induces G2/M cell cycle arrest and apoptosis involving p38 MAPK activation in human breast cancer cells. Food Chem. Toxicol. 2014, 67, 193–200. [Google Scholar] [CrossRef]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Vara, J.Á.F.; Casado, E.; De Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Morgensztern, D.; McLeod, H.L. PI3K/Akt/mTOR pathway as a target for cancer therapy. Anti-Cancer Drugs 2005, 16, 797–803. [Google Scholar] [CrossRef]
- Tewari, D.; Patni, P.; Bishayee, A.; Sah, A.N.; Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: A novel therapeutic strategy. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022; pp. 1–17. [Google Scholar]
- Guan, X.; Pan, Z.; Xu, Z.; Zhang, S.; Tang, H.; Du, G.; Zhou, J. Natural flavonoid luteolin promotes the differentiation of porcine myoblasts through activation of PI3K/Akt/mTOR signaling. Food Biosci. 2022, 47, 101766. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, J.; Hou, J.; Hui, M.; Qi, H.; Lei, T.; Zhang, X.; Zhao, L.; Du, H. Induction of autophagy via the PI3K/Akt/mTOR signaling pathway by Pueraria flavonoids improves non-alcoholic fatty liver disease in obese mice. Biomed. Pharmacother. 2023, 157, 114005. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.-M.; Park, J.E.; Lee, J.Y.; Park, K.-R. Latifolin, a Natural Flavonoid, Isolated from the Heartwood of Dalbergia odorifera Induces Bioactivities through Apoptosis, Autophagy, and Necroptosis in Human Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 13629. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abusaliya, A.; Jeong, S.H.; Bhosale, P.B.; Kim, H.H.; Park, M.Y.; Kim, E.; Won, C.K.; Park, K.I.; Heo, J.D.; Kim, H.W.; et al. Mechanistic Action of Cell Cycle Arrest and Intrinsic Apoptosis via Inhibiting Akt/mTOR and Activation of p38-MAPK Signaling Pathways in Hep3B Liver Cancer Cells by Prunetrin—A Flavonoid with Therapeutic Potential. Nutrients 2023, 15, 3407. https://doi.org/10.3390/nu15153407
Abusaliya A, Jeong SH, Bhosale PB, Kim HH, Park MY, Kim E, Won CK, Park KI, Heo JD, Kim HW, et al. Mechanistic Action of Cell Cycle Arrest and Intrinsic Apoptosis via Inhibiting Akt/mTOR and Activation of p38-MAPK Signaling Pathways in Hep3B Liver Cancer Cells by Prunetrin—A Flavonoid with Therapeutic Potential. Nutrients. 2023; 15(15):3407. https://doi.org/10.3390/nu15153407
Chicago/Turabian StyleAbusaliya, Abuyaseer, Se Hyo Jeong, Pritam Bhagwan Bhosale, Hun Hwan Kim, Min Yeong Park, Eunhye Kim, Chung Kil Won, Kwang Il Park, Jeong Doo Heo, Hyun Wook Kim, and et al. 2023. "Mechanistic Action of Cell Cycle Arrest and Intrinsic Apoptosis via Inhibiting Akt/mTOR and Activation of p38-MAPK Signaling Pathways in Hep3B Liver Cancer Cells by Prunetrin—A Flavonoid with Therapeutic Potential" Nutrients 15, no. 15: 3407. https://doi.org/10.3390/nu15153407