
Improving the Antitumor Effect of Chemotherapy with Ocoxin as a Novel Adjuvant Agent to Treat Prostate Cancer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Ocoxin
2.3. Cell-Viability Assay
2.4. Cell-Cycle Analysis
2.5. mRNA Sequencing for LNCaP Cell Gene-Expression Analysis
2.6. Transcriptomic Analysis of LNCaP Cells Treated with Ocoxin
2.7. Quantification of the Differential Expression of Genes Altered by Ocoxin through RT-qPCR
2.8. Obtention of Cancer-Associated Fibroblast- and Osteoblast-Derived Secretomes
2.9. Chemoresistance Analysis in Prostate-Cancer Cells in the Presence of CAF and Osteoblast Secretomes
2.10. Cell Migration Assay
2.11. Animals
2.12. In Vivo Prostate-Cancer Tumor Development
2.13. Immunohistochemical and Immunofluorescence Analyses
2.14. Statistical Analysis
3. Results
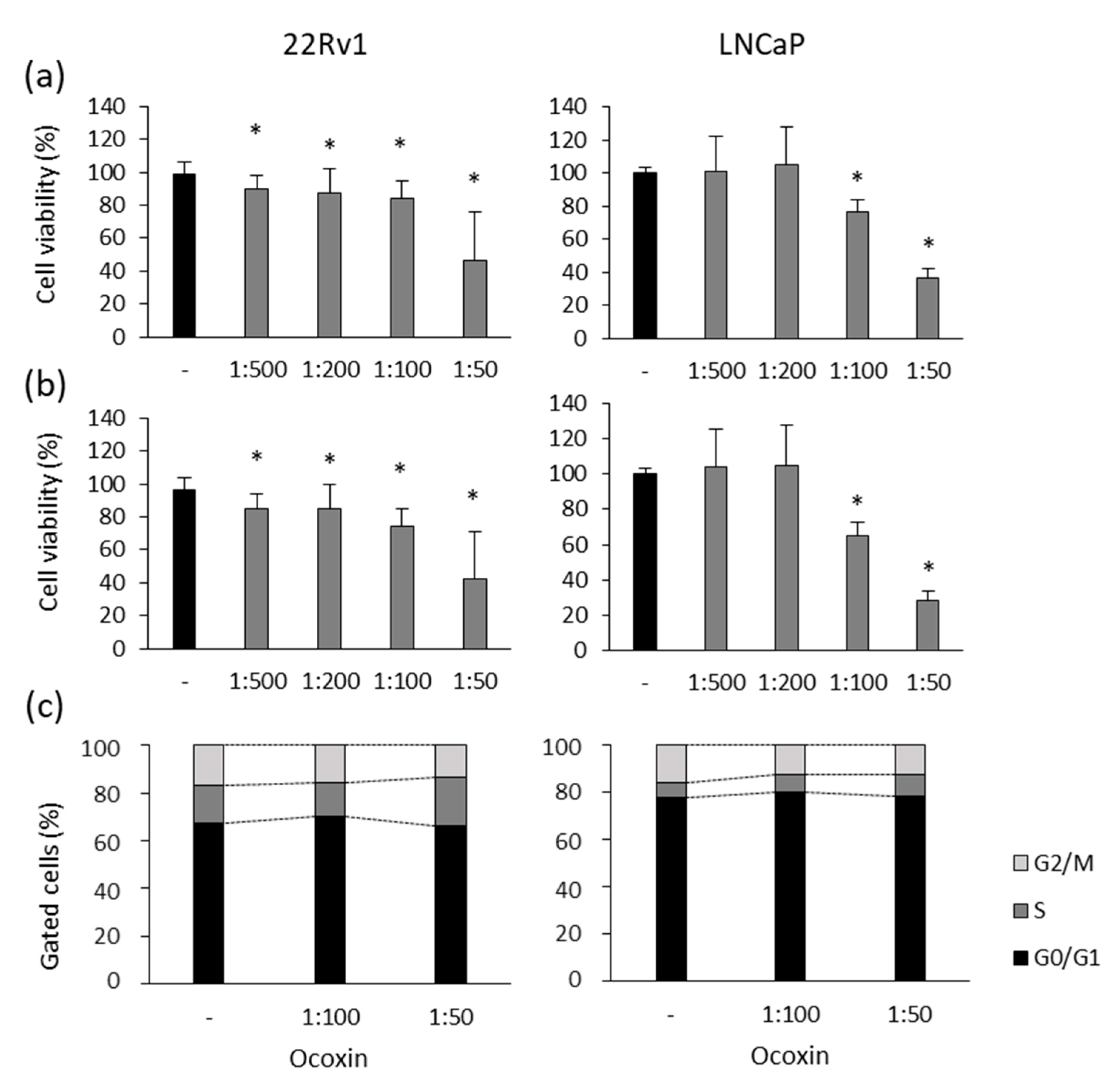
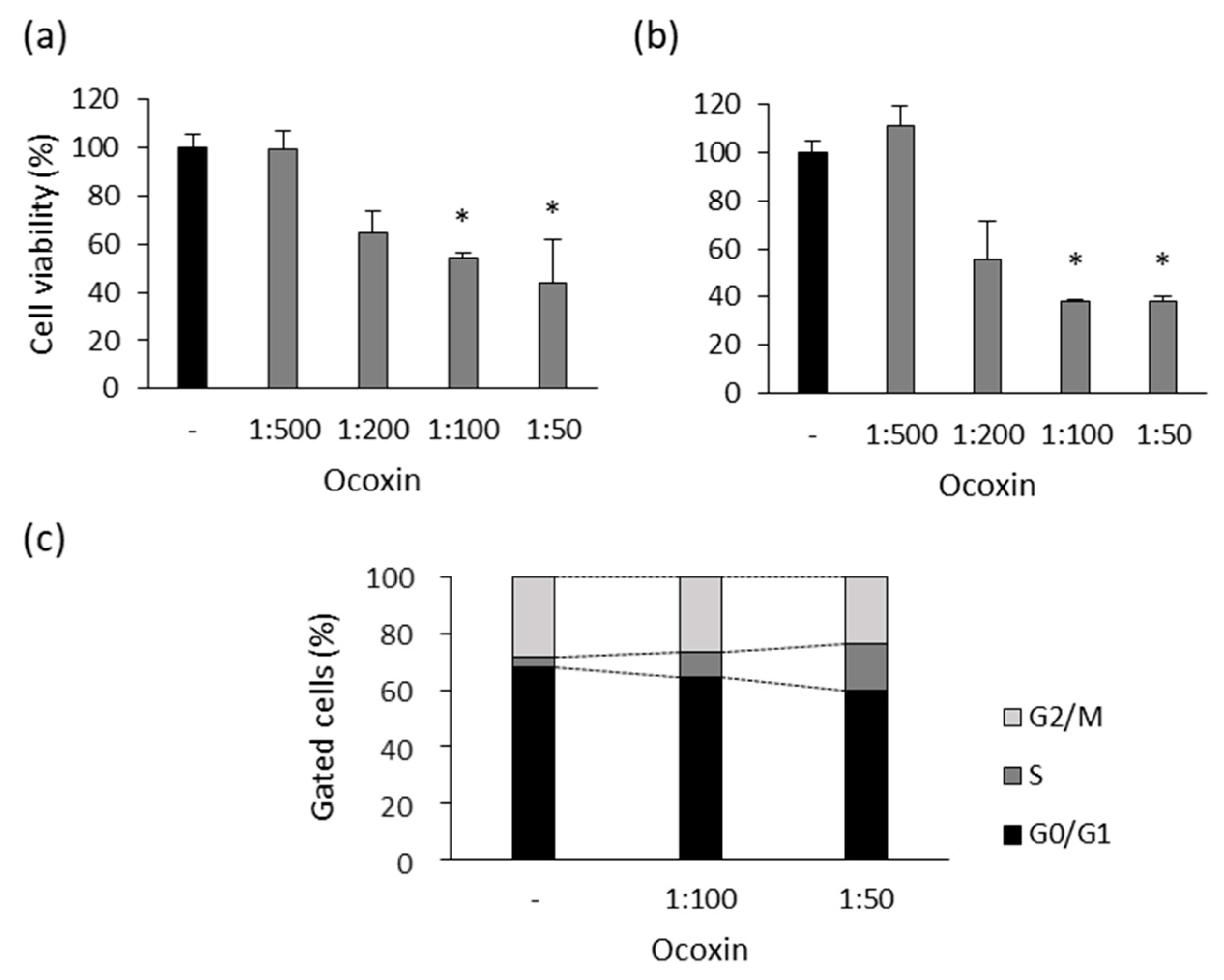
3.1. Ocoxin Altered the Viability and Cell Cycle of Human Prostate-Cancer Cell Lines
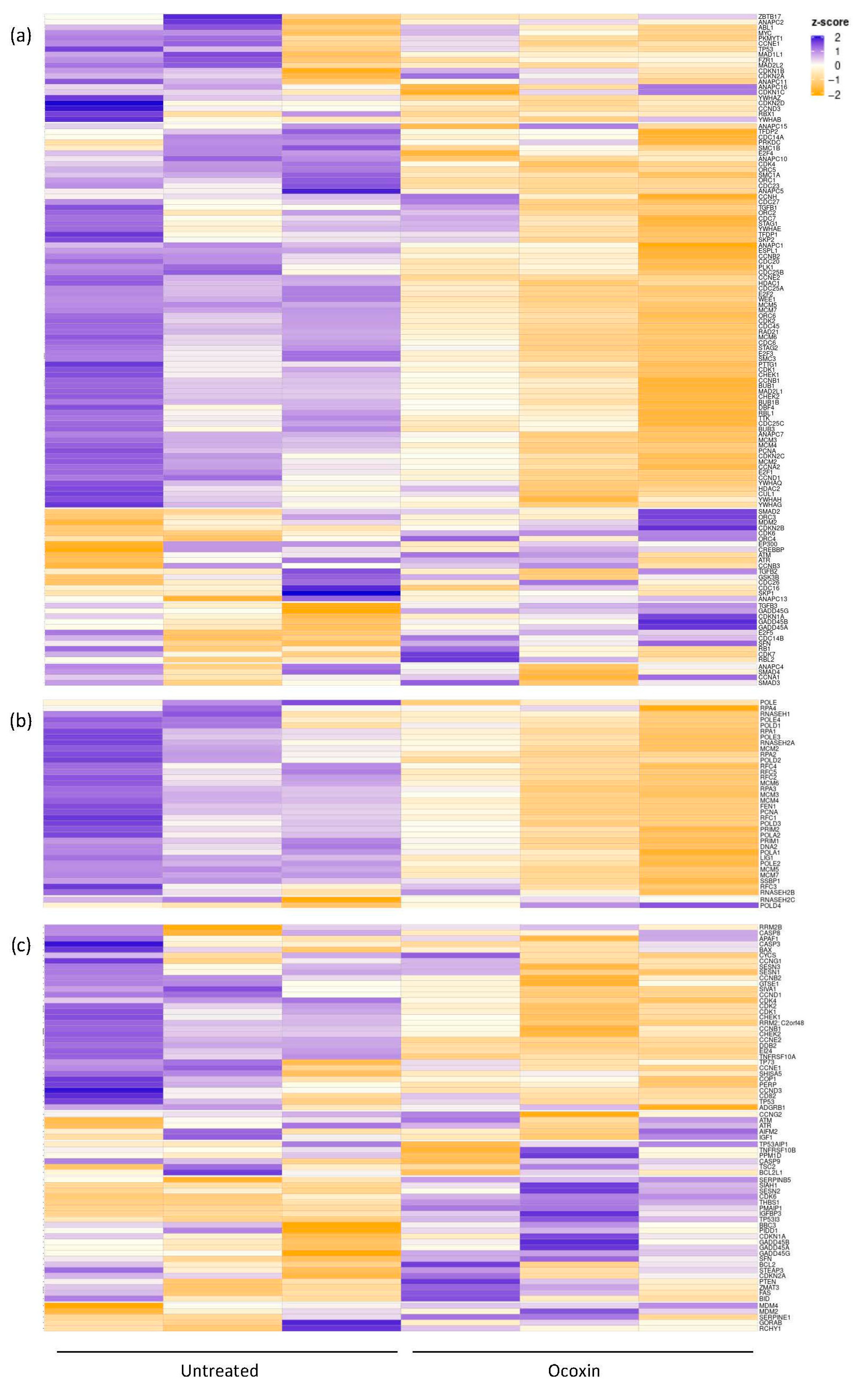
3.2. mRNAseq for the Analysis of Gene Expression of LNCaP Cells Treated with Ocoxin
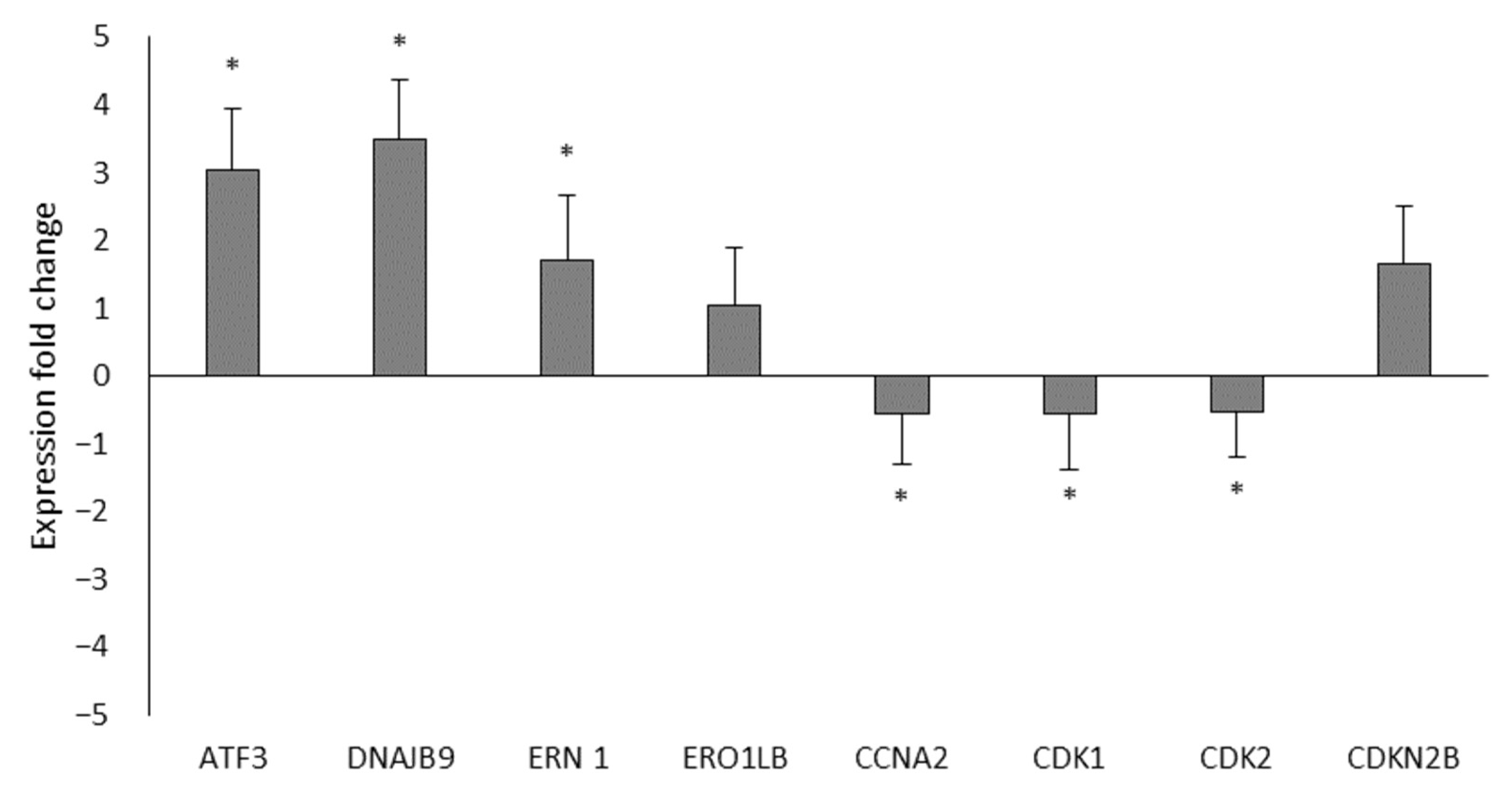
3.3. Analysis of the Differential Expression of LNCaP Cell Genes Treated with Ocoxin by RT-qPCR
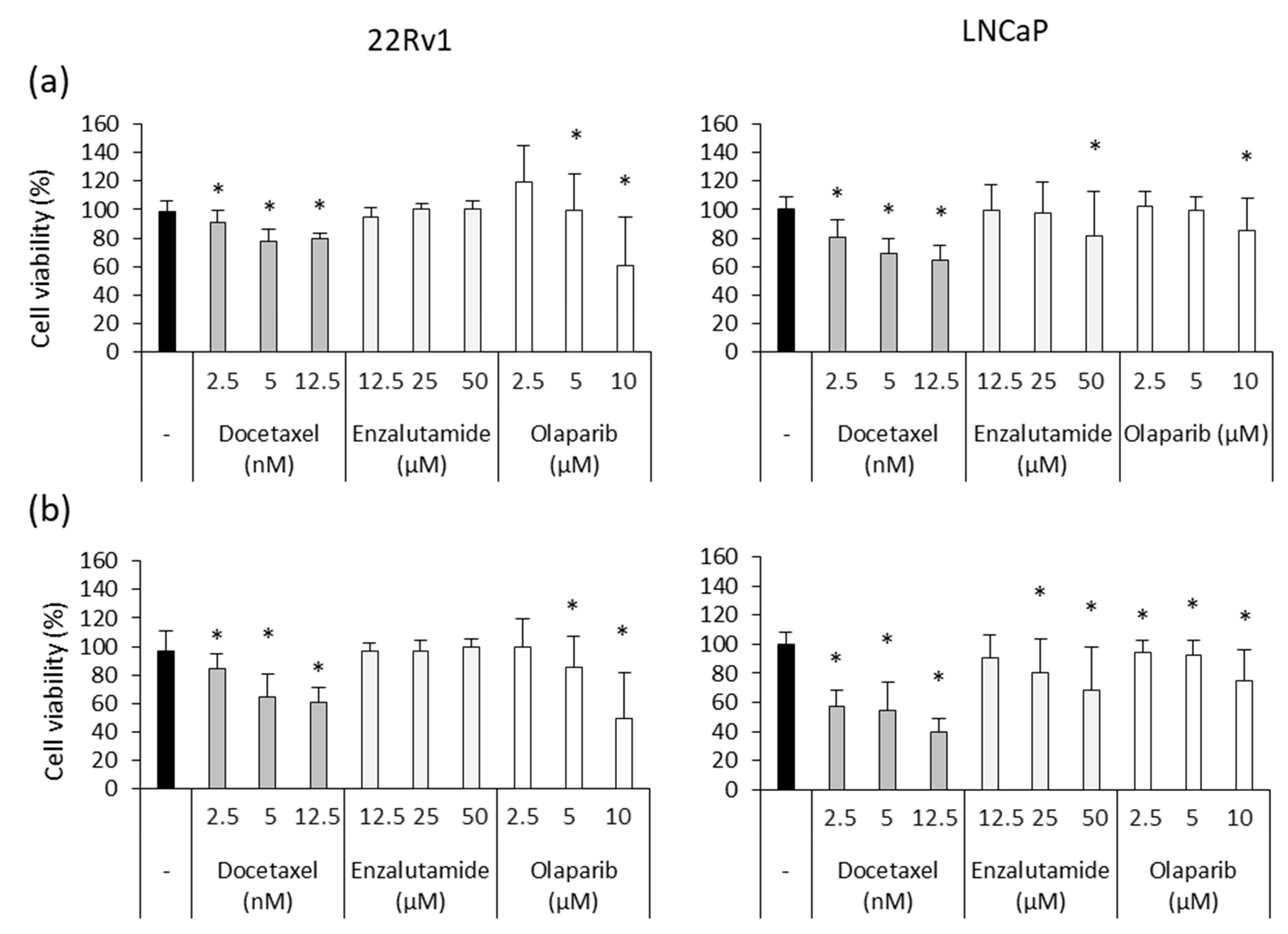
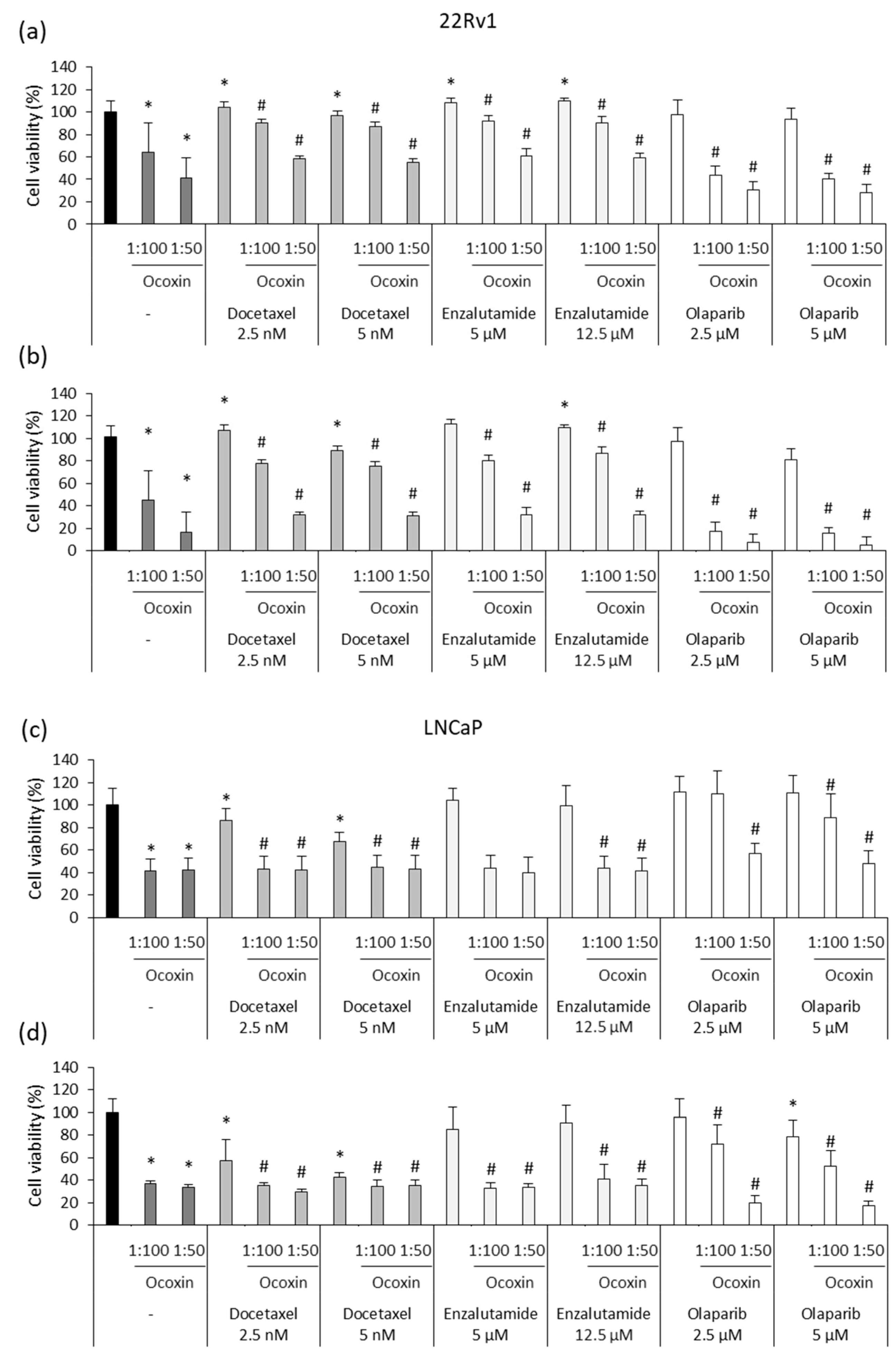
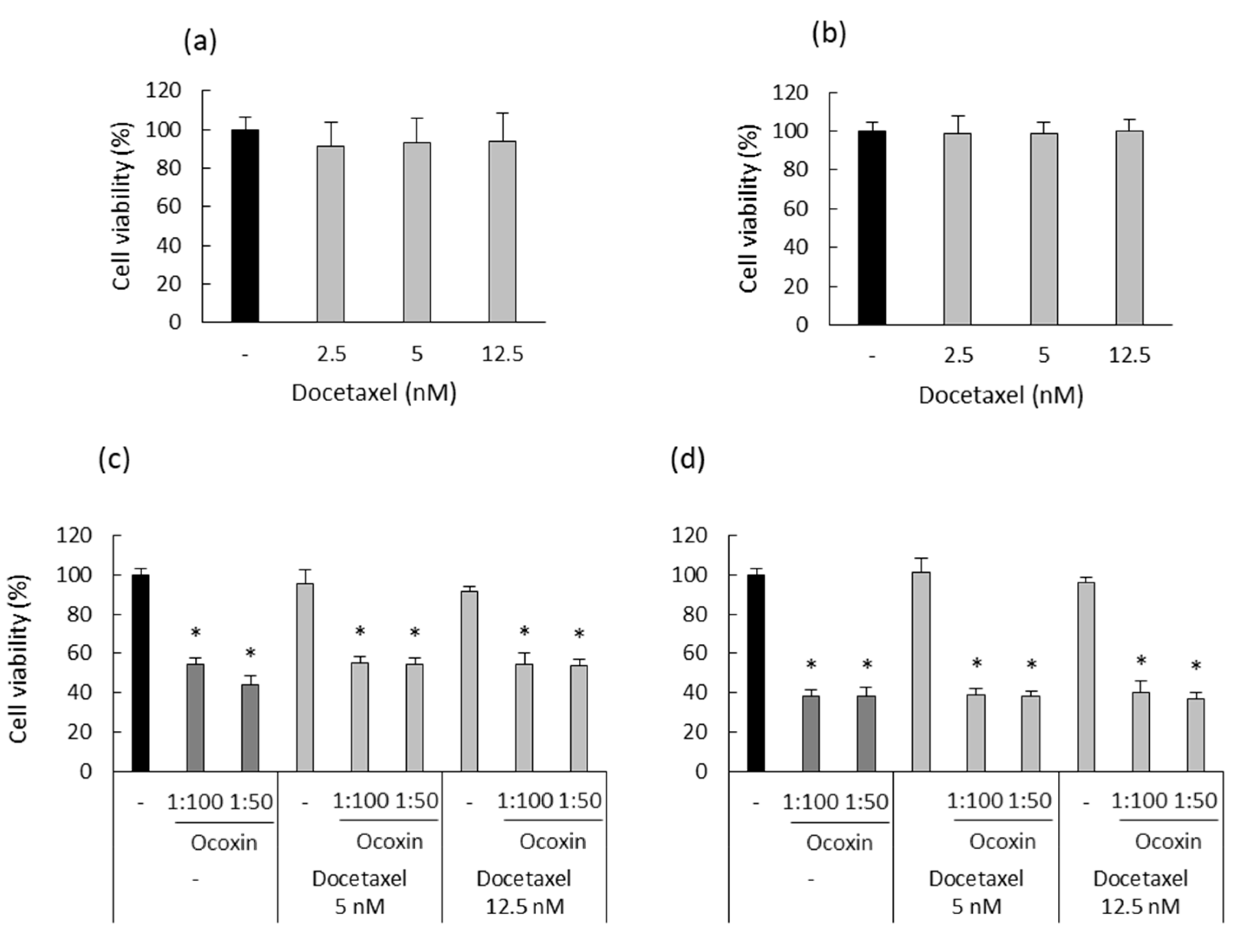
3.4. Ocoxin as an Adjuvant Agent of Docetaxel, Enzalutamide and Olaparib Increased the Cytotoxic Effect in Human Prostate-Cancer Cell Lines
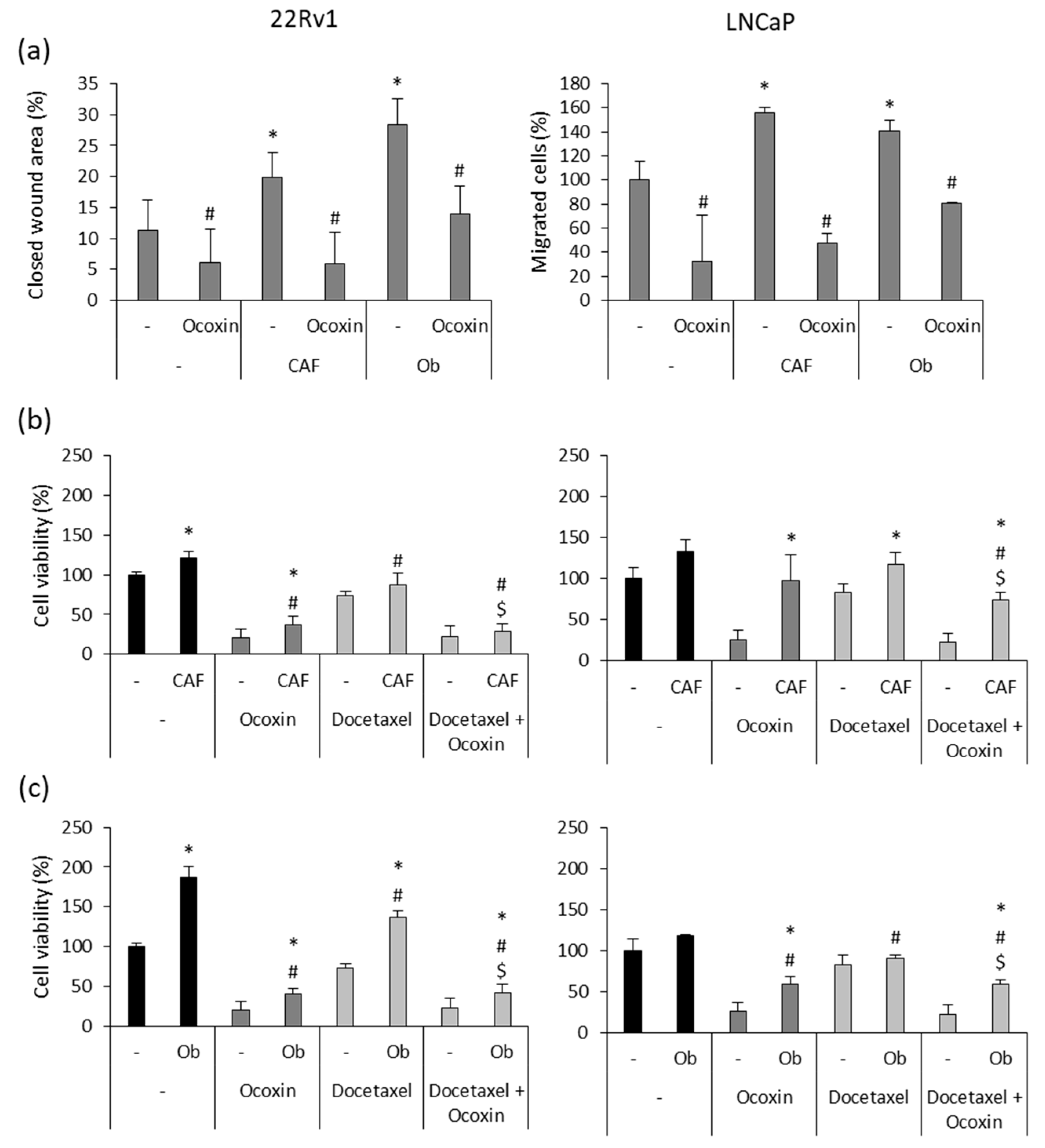
3.5. Ocoxin Reduced the Pro-Migratory Effect and Chemoresistance Produced by Soluble Factors Derived from Osteoblasts and CAFs Secreted on Human Prostate-Cancer Cells
3.6. Ocoxin Altered the Viability and Cell Cycle of a Murine Prostate-Cancer Cell Line
3.7. Ocoxin as an Adjuvant Agent of Docetaxel Increased the Cytotoxic Effect in a Murine Prostate-Cancer Cell Line
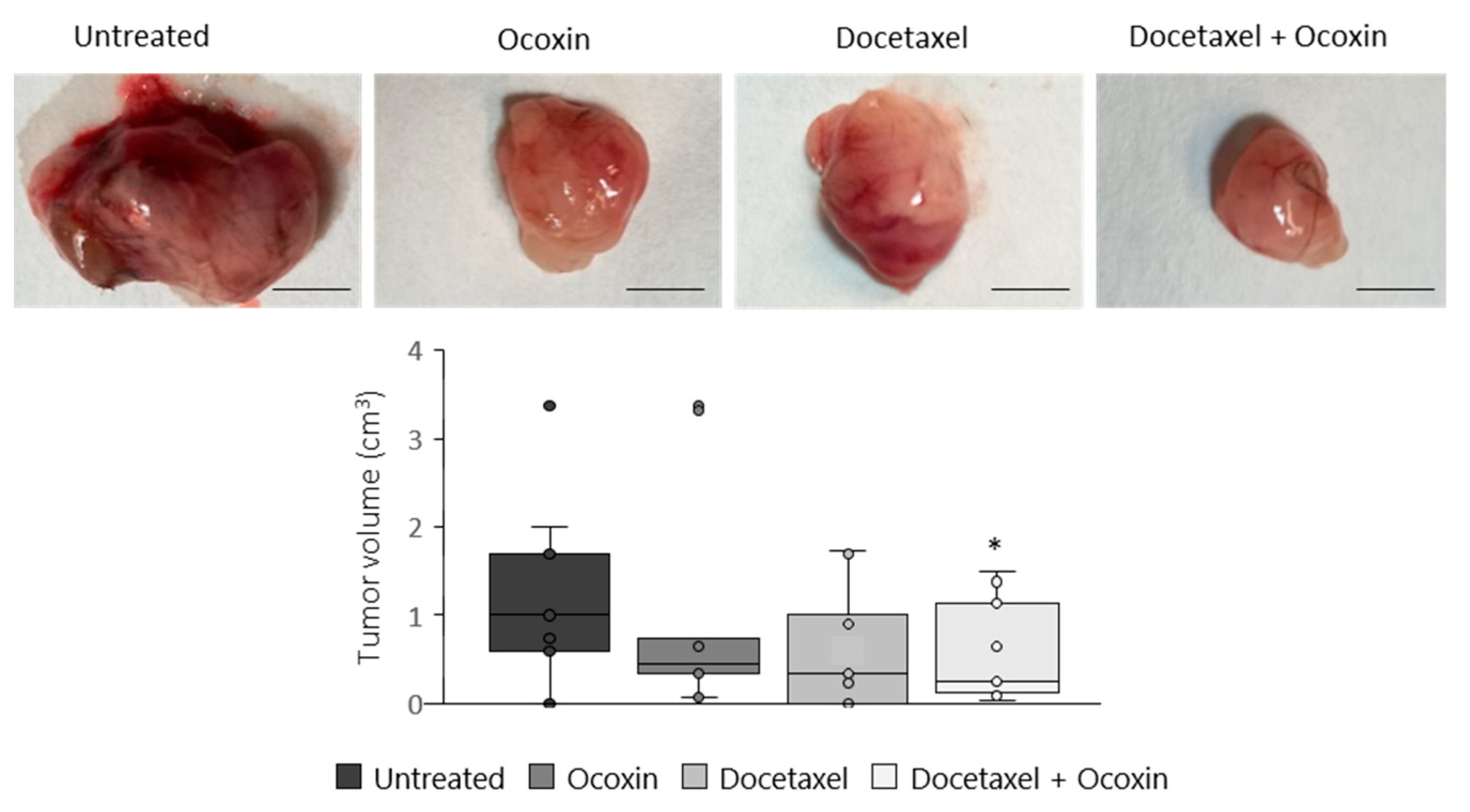
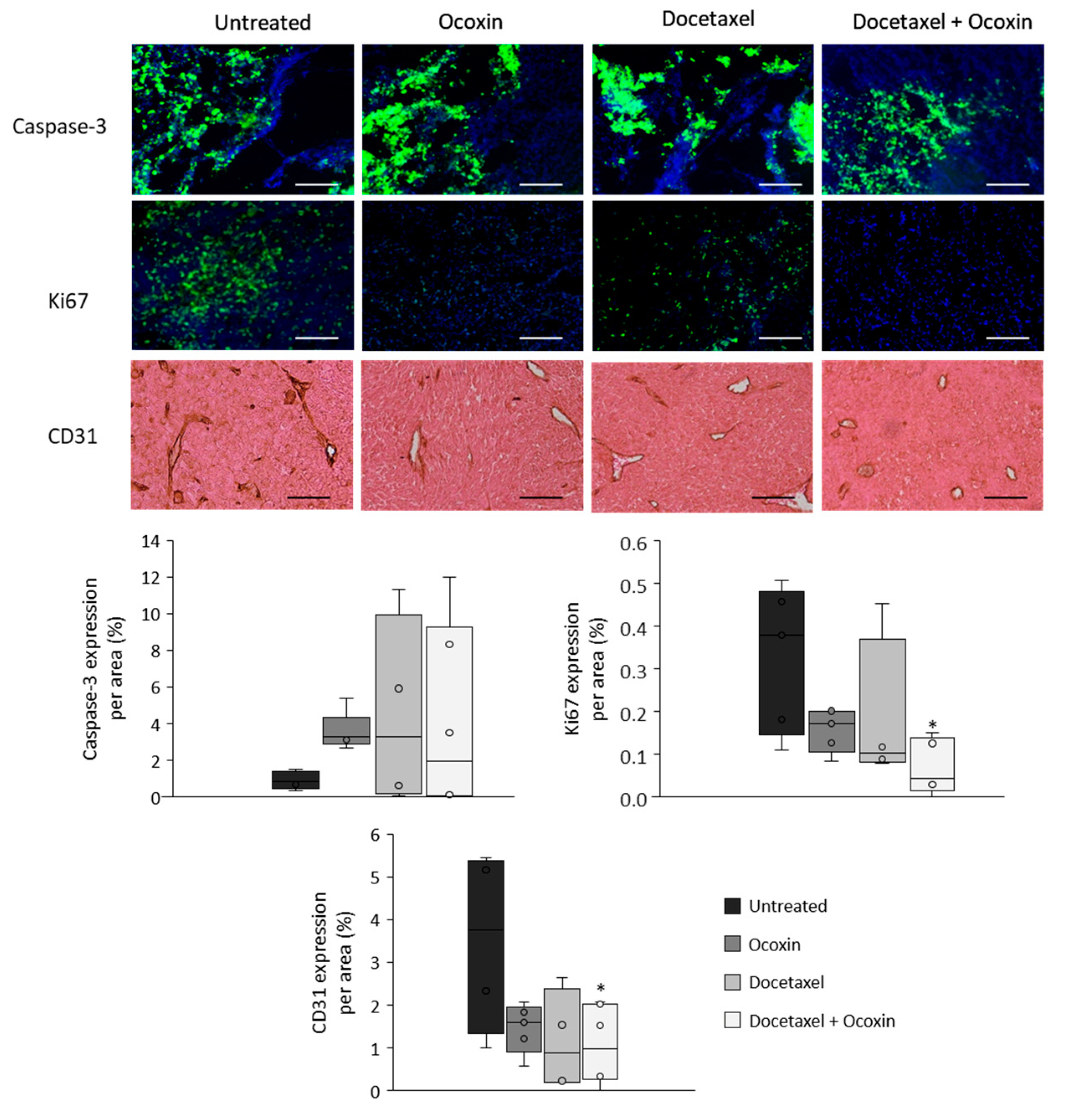
3.8. The Administration of Ocoxin as an Adjuvant of Docetaxel Reduced Prostate Tumor Volume In Vivo by Increasing Apoptosis and by Decreasing Proliferation and Angiogenesis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Litwin, M.S.; Tan, H. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef]
- Hoang, D.T.; Iczkowski, K.A.; Kilari, D.; See, W.; Nevalainen, M.T. Androgen receptor-dependent and -independent mechanisms driving prostate cancer progression: Opportunities for therapeutic targeting from multiple angles. Oncotarget 2017, 8, 3724–3745. [Google Scholar] [CrossRef]
- Merseburger, A.S.; Hupe, M.C. An Update on Triptorelin: Current Thinking on Androgen Deprivation Therapy for Prostate Cancer. Adv. Ther. 2016, 33, 1072–1093. [Google Scholar] [CrossRef]
- Vellky, J.E.; Ricke, W.A. Development and prevalence of castration-resistant prostate cancer subtypes. Neoplasia 2020, 22, 566–575. [Google Scholar] [CrossRef]
- Schatten, H. Brief Overview of Prostate Cancer Statistics, Grading, Diagnosis and Treatment Strategies. Adv. Exp. Med. Biol. 2018, 1095, 1–14. [Google Scholar] [PubMed]
- De Bono, J.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. Olaparib for Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 382, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Dror, C.M.; Wyatt, A.W.; Chi, K.N. Olaparib for the treatment of metastatic prostate cancer. Future Oncol. 2021, 17, 2413–2429. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.; Carreira, S.; Sandhu, S.; Miranda, S.; Mossop, H.; Perez-Lopez, R.; Nava Rodrigues, D.; Robinson, D.; Omlin, A.; Tunariu, N.; et al. DNA-Repair Defects and Olaparib in Metastatic Prostate Cancer. N. Engl. J. Med. 2015, 373, 1697–1708. [Google Scholar] [CrossRef]
- Hussain, M.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. PROfound Trial Investigators Survival with Olaparib in Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 383, 2345–2357. [Google Scholar] [CrossRef]
- Shevach, J.; Weiner, A.; Morgans, A.K. Quality of Life-Focused Decision-Making for Prostate Cancer. Curr. Urol. Rep. 2019, 20, 57. [Google Scholar] [CrossRef]
- Nagireddy, S.; Qureshi, R.; Best, J.; Frech, F.S.; Shah, K.; Soni, Y.; Kuchakulla, M.; Narasimman, M.; Arora, H. Current Treatment Modalities Targeting Tumor Microenvironment in Castration-Resistant Prostate Cancer. Adv. Exp. Med. Biol. 2021, 1329, 295–323. [Google Scholar] [PubMed]
- Sounni, N.E.; Noel, A. Targeting the tumor microenvironment for cancer therapy. Clin. Chem. 2013, 59, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.K.; Rise, K.; Giskeødegård, G.F.; Richardsen, E.; Bertilsson, H.; Størkersen, Ø.; Bathen, T.F.; Rye, M.; Tessem, M. Integrative metabolic and transcriptomic profiling of prostate cancer tissue containing reactive stroma. Sci. Rep. 2018, 8, 14269. [Google Scholar] [CrossRef]
- Shiao, S.L.; Chu, G.C.; Chung, L.W.K. Regulation of prostate cancer progression by the tumor microenvironment. Cancer Lett. 2016, 380, 340–348. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer. 2020, 20, 174–186. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Schoppmann, S.F.; Birner, P.; Stöckl, J.; Kalt, R.; Ullrich, R.; Caucig, C.; Kriehuber, E.; Nagy, K.; Alitalo, K.; Kerjaschki, D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am. J. Pathol. 2002, 161, 947–956. [Google Scholar] [CrossRef]
- Martori, C.; Sanchez-Moral, L.; Paul, T.; Pardo, J.C.; Font, A.; Ruiz de Porras, V.; Sarrias, M. Macrophages as a Therapeutic Target in Metastatic Prostate Cancer: A Way to Overcome Immunotherapy Resistance? Cancers 2022, 14, 440. [Google Scholar] [CrossRef]
- Han, I.; Song, H.; Ryu, J. IL-6 produced by prostate epithelial cells stimulated with Trichomonas vaginalis promotes proliferation of prostate cancer cells by inducing M2 polarization of THP-1-derived macrophages. PLoS Negl. Trop. Dis. 2020, 14, e0008126. [Google Scholar] [CrossRef]
- Prenen, H.; Mazzone, M. Tumor-associated macrophages: A short compendium. Cell Mol. Life Sci. 2019, 76, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Karpisheh, V.; Mousavi, S.M.; Naghavi Sheykholeslami, P.; Fathi, M.; Mohammadpour Saray, M.; Aghebati-Maleki, L.; Jafari, R.; Majidi Zolbanin, N.; Jadidi-Niaragh, F. The role of regulatory T cells in the pathogenesis and treatment of prostate cancer. Life Sci. 2021, 284, 119132. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.T.W.; Bryant, R.J.; Parkes, E.E. The tumor microenvironment and immune responses in prostate cancer patients. Endocr. Relat. Cancer 2021, 28, T95–T107. [Google Scholar] [CrossRef]
- Al-Janabi, H.; Lewis, C.E. Macrophage Regulation of the Development of Castration-Resistant Prostate Cancer. Cancer Res. 2021, 81, 5399–5400. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.; Lee, J.H.; Ponnazhagan, S. Revisiting Immunotherapy: A Focus on Prostate Cancer. Cancer Res. 2020, 80, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, A.Z.; Dallos, M.C.; Zahurak, M.L.; Partin, A.W.; Schaeffer, E.M.; Ross, A.E.; Allaf, M.E.; Nirschl, T.R.; Liu, D.; Chapman, C.G.; et al. T-Cell Infiltration and Adaptive Treg Resistance in Response to Androgen Deprivation with or Without Vaccination in Localized Prostate Cancer. Clin. Cancer Res. 2020, 26, 3182–3192. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Y.; Gao, W. The evolving role of immune cells in prostate cancer. Cancer Lett. 2022, 525, 9–21. [Google Scholar] [CrossRef]
- Zhang, Z.; Karthaus, W.R.; Lee, Y.S.; Gao, V.R.; Wu, C.; Russo, J.W.; Liu, M.; Mota, J.M.; Abida, W.; Linton, E.; et al. Tumor Microenvironment-Derived NRG1 Promotes Antiandrogen Resistance in Prostate Cancer. Cancer Cell 2020, 38, 279–296.e9. [Google Scholar] [CrossRef]
- Haider, M.; Holen, I.; Dear, T.N.; Hunter, K.; Brown, H.K. Modifying the osteoblastic niche with zoledronic acid in vivo-potential implications for breast cancer bone metastasis. Bone 2014, 66, 240–250. [Google Scholar] [CrossRef]
- Hensel, J.; Thalmann, G.N. Biology of Bone Metastases in Prostate Cancer. Urology 2016, 92, 6–13. [Google Scholar] [CrossRef]
- Massagué, J.; Obenauf, A.C. Metastatic colonization by circulating tumour cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Chen, Q.; Jing, R.; He, X.; Wang, H.; Ban, Y.; Ye, Q.; Xu, W.; Zheng, C. Molecular Basis of Prostate Cancer and Natural Products as Potential Chemotherapeutic and Chemopreventive Agents. Front. Pharmacol. 2021, 12, 738235. [Google Scholar] [CrossRef] [PubMed]
- Crocetto, F.; di Zazzo, E.; Buonerba, C.; Aveta, A.; Pandolfo, S.D.; Barone, B.; Trama, F.; Caputo, V.F.; Scafuri, L.; Ferro, M.; et al. Kaempferol, Myricetin and Fisetin in Prostate and Bladder Cancer: A Systematic Review of the Literature. Nutrients 2021, 13, 3750. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Yamthe, L.R.T.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martorell, M.; Setzer, W.N.; Rodrigues, C.F.; et al. Phytochemicals in Prostate Cancer: From Bioactive Molecules to Upcoming Therapeutic Agents. Nutrients 2019, 11, 1483. [Google Scholar] [CrossRef]
- Sekhoacha, M.; Riet, K.; Motloung, P.; Gumenku, L.; Adegoke, A.; Mashele, S. Prostate Cancer Review: Genetics, Diagnosis, Treatment Options, and Alternative Approaches. Molecules 2022, 27, 5730. [Google Scholar] [CrossRef]
- Wang, X.; Fang, G.; Pang, Y. Chinese Medicines in the Treatment of Prostate Cancer: From Formulas to Extracts and Compounds. Nutrients 2018, 10, 283. [Google Scholar] [CrossRef]
- Silk, N.; Reich, J.; Sinha, R.; Chawla, S.; Geary, K.; Zhang, D. The Effects of Resveratrol on Prostate Cancer through Targeting the Tumor Microenvironment. J. Xenobiot. 2021, 11, 16–32. [Google Scholar] [CrossRef]
- Pietrovito, L.; Iozzo, M.; Bacci, M.; Giannoni, E.; Chiarugi, P. Treatment with Cannabinoids as a Promising Approach for Impairing Fibroblast Activation and Prostate Cancer Progression. Int. J. Mol. Sci. 2020, 21, 787. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, X.; Chen, Y.; Tao, N.; Qin, Z. Ligustilide Inhibits Tumor Angiogenesis by Downregulating VEGFA Secretion from Cancer-Associated Fibroblasts in Prostate Cancer via TLR4. Cancers 2022, 14, 2406. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sethi, G.; Baladandayuthapani, V.; Krishnan, S.; Shishodia, S. Targeting cell signaling pathways for drug discovery: An old lock needs a new key. J. Cell. Biochem. 2007, 102, 580–592. [Google Scholar] [CrossRef]
- Garodia, P.; Ichikawa, H.; Malani, N.; Sethi, G.; Aggarwal, B.B. From ancient medicine to modern medicine: Ayurvedic concepts of health and their role in inflammation and cancer. J. Soc. Integr. Oncol. 2007, 5, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Kannaiyan, R.; Sethi, G. Targeting cell signaling and apoptotic pathways by dietary agents: Role in the prevention and treatment of cancer. Nutr. Cancer 2011, 63, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Lecumberri, E.; Dupertuis, Y.M.; Miralbell, R.; Pichard, C. Green tea polyphenol epigallocatechin-3-gallate (EGCG) as adjuvant in cancer therapy. Clin. Nutr. 2013, 32, 894–903. [Google Scholar] [PubMed]
- Nisar, S.; Masoodi, T.; Prabhu, K.S.; Kuttikrishnan, S.; Zarif, L.; Khatoon, S.; Ali, S.; Uddin, S.; Akil, A.A.; Singh, M.; et al. Natural products as chemo-radiation therapy sensitizers in cancers. Biomed. Pharmacother. 2022, 154, 113610. [Google Scholar] [CrossRef]
- Singla, R.K.; Sai, C.S.; Chopra, H.; Behzad, S.; Bansal, H.; Goyal, R.; Gautam, R.K.; Tsagkaris, C.; Joon, S.; Singla, S.; et al. Natural Products for the Management of Castration-Resistant Prostate Cancer: Special Focus on Nanoparticles Based Studies. Front. Cell Dev. Biol. 2021, 9, 745177. [Google Scholar]
- Wang, K.; Yu, Y.; Chen, H.; Chiang, Y.; Ali, M.; Shieh, T.; Hsia, S. Recent Advances in Glycyrrhiza glabra (Licorice)-Containing Herbs Alleviating Radiotherapy- and Chemotherapy-Induced Adverse Reactions in Cancer Treatment. Metabolites 2022, 12, 535. [Google Scholar] [CrossRef]
- Kallifatidis, G.; Hoy, J.J.; Lokeshwar, B.L. Bioactive natural products for chemoprevention and treatment of castration-resistant prostate cancer. Semin. Cancer Biol. 2016, 40–41, 160–169. [Google Scholar] [CrossRef]
- Benedicto, A.; Hernandez-Unzueta, I.; Sanz, E.; Márquez, J. Ocoxin Increases the Antitumor Effect of BRAF Inhibition and Reduces Cancer Associated Fibroblast-Mediated Chemoresistance and Protumoral Activity in Metastatic Melanoma. Nutrients 2021, 13, 686. [Google Scholar] [CrossRef]
- Hernandez-Unzueta, I.; Benedicto, A.; Olaso, E.; Sanz, E.; Viera, C.; Arteta, B.; Márquez, J. Ocoxin oral solution(®) as a complement to irinotecan chemotherapy in the metastatic progression of colorectal cancer to the liver. Oncol. Lett. 2017, 13, 4002–4012. [Google Scholar]
- Hernandez-Unzueta, I.; Benedicto, A.; Romayor, I.; Herrero, A.; Sanz, E.; Arteta, B.; Olaso, E.; Márquez, J. Ocoxin Oral Solution Exerts an Antitumoral Effect in Pancreatic Cancer and Reduces the Stromal-Mediated Chemoresistance. Pancreas 2019, 48, 555–567. [Google Scholar]
- Márquez, J.; Mena, J.; Hernandez-Unzueta, I.; Benedicto, A.; Sanz, E.; Arteta, B.; Olaso, E. Ocoxin® oral solution slows down tumor growth in an experimental model of colorectal cancer metastasis to the liver in Balb/c mice. Oncol. Rep. 2016, 35, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Thirugnanam, S.; Xu, L.; Ramaswamy, K.; Gnanasekar, M. Glycyrrhizin induces apoptosis in prostate cancer cell lines DU-145 and LNCaP. Oncol. Rep. 2008, 20, 1387–1392. [Google Scholar]
- Shirakami, Y.; Shimizu, M. Possible Mechanisms of Green Tea and Its Constituents against Cancer. Molecules 2018, 23, 2284. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Davoodvandi, A.; Pourhanifeh, M.H.; Sharifi, N.; ArefNezhad, R.; Sahebnasagh, R.; Moghadam, S.A.; Sahebkar, A.; Mirzaei, H. Anti-cancer effects of cinnamon: Insights into its apoptosis effects. Eur. J. Med. Chem. 2019, 178, 131–140. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2022, 51, D587–D592. [Google Scholar] [CrossRef]
- Al-Mahtab, M.; Akbar, S.M.F.; Khan, M.S.I.; Rahman, S. Increased survival of patients with end-stage hepatocellular carcinoma due to intake of ONCOXIN®, a dietary supplement. Indian J. Cancer 2015, 52, 443–446. [Google Scholar] [PubMed]
- Kaidarova, D.R.; Kopp, M.V.; Pokrovsky, V.S.; Dzhugashvili, M.; Akimzhanova, Z.M.; Abdrakhmanov, R.Z.; Babich, E.N.; Bilan, E.V.; Byakhov, A.V.; Gurov, S.N.; et al. Multicomponent nutritional supplement Oncoxin and its influence on quality of life and therapy toxicity in patients receiving adjuvant chemotherapy. Oncol. Lett. 2019, 18, 5644–5652. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Lorente, R.; Hernández Durán, D.; García Viamontes, J.; Lence Anta, J.; Ortiz Reyes, R.; Sanz Navares, E. Efficacy of Oncoxin-Viusid on the Reduction of Adverse Reactions to Chemotherapy and Radiotherapy in Patients Diagnosed with Cervical Cancer and Endometrial Adenocarcinoma. J. Cancer Ther. 2020, 11, 276–295. [Google Scholar] [CrossRef]
- Shumsky, A.; Bilan, E.; Sanz, E.; Petrovskiy, F. Oncoxin nutritional supplement in the management of chemotherapy- and/or radiotherapy-associated oral mucositis. Mol. Clin. Oncol. 2019, 10, 463–468. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]
- Portela Pereira, P.; Gómez-Veiga, F. Advanced prostate cancer and quality of life. Arch. Esp. Urol. 2018, 71, 306–314. [Google Scholar]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. Natural Compounds in Prostate Cancer Prevention and Treatment: Mechanisms of Action and Molecular Targets. Cells 2020, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Termini, D.; Den Hartogh, D.J.; Jaglanian, A.; Tsiani, E. Curcumin against Prostate Cancer: Current Evidence. Biomolecules 2020, 10, 1536. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M. Green tea: An effective synergist with anticancer drugs for tertiary cancer prevention. Cancer Lett. 2012, 324, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Emerging Adjuvant Therapy for Cancer: Propolis and its Constituents. J. Diet. Suppl. 2016, 13, 245–268. [Google Scholar] [CrossRef]
- Sánchez, B.G.; Bort, A.; Mateos-Gómez, P.A.; Rodríguez-Henche, N.; Díaz-Laviada, I. Combination of the natural product capsaicin and docetaxel synergistically kills human prostate cancer cells through the metabolic regulator AMP-activated kinase. Cancer. Cell. Int. 2019, 19, 54. [Google Scholar] [CrossRef]
- Tummala, R.; Lou, W.; Gao, A.C.; Nadiminty, N. Quercetin Targets hnRNPA1 to Overcome Enzalutamide Resistance in Prostate Cancer Cells. Mol. Cancer Ther. 2017, 16, 2770–2779. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Henning, S.M.; Heber, D.; Vadgama, J.V. Sensitization to docetaxel in prostate cancer cells by green tea and quercetin. J. Nutr. Biochem. 2015, 26, 408–415. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Bahramsoltani, R.; Rahimi, R. Phytochemicals as Adjunctive with Conventional Anticancer Therapies. Curr. Pharm. Des. 2016, 22, 4201–4218. [Google Scholar] [CrossRef]
- Qi, F.; Zhao, L.; Zhou, A.; Zhang, B.; Li, A.; Wang, Z.; Han, J. The advantages of using traditional Chinese medicine as an adjunctive therapy in the whole course of cancer treatment instead of only terminal stage of cancer. Biosci. Trends 2015, 9, 16–34. [Google Scholar] [CrossRef]
- Fundora Ramos, M.I.; Maden, L.B.; Casanova, F.O.; Cruz, F.H.; Reyes, C.S.; Gato, A.H.; Lyncon, I.B.; González, E.V.; Morales, K.P.; Lence, J.J.; et al. Oncoxin-Viusid(®) may improve quality of life and survival in patients with hormone-refractory prostate cancer undergoing onco-specific treatments. Mol. Clin. Oncol. 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Chakraborty, A. Cinnamon in Anticancer Armamentarium: A Molecular Approach. J. Toxicol. 2018, 2018, 8978731. [Google Scholar] [CrossRef] [PubMed]
- Pandiella-Alonso, A.; Díaz-Rodríguez, E.; Sanz, E. Antitumoral Properties of the Nutritional Supplement Ocoxin Oral Solution: A Comprehensive Review. Nutrients 2020, 12, 2661. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, J.; Qu, X.; Zhu, N.; Ji, Y. Downregulation of Cdk1 and cyclinB1 expression contributes to oridonin-induced cell cycle arrest at G2/M phase and growth inhibition in SGC-7901 gastric cancer cells. Asian Pac. J. Cancer Prev. 2014, 15, 6437–6441. [Google Scholar] [CrossRef]
- Wu, J.; Lv, Q.; He, J.; Zhang, H.; Mei, X.; Cui, K.; Huang, N.; Xie, W.; Xu, N.; Zhang, Y. MicroRNA-188 suppresses G1/S transition by targeting multiple cyclin/CDK complexes. Cell. Commun. Signal. 2014, 12, 66. [Google Scholar] [CrossRef]
- Casado-Vela, J.; Martínez-Torrecuadrada, J.L.; Casal, J.I. Differential phosphorylation patterns between the Cyclin-A2/CDK2 complex and their monomers. Protein Expr. Purif. 2009, 66, 15–21. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Zhou, Y.; Wang, F.; Yin, C.; Ding, L.; Zhang, S. Tanshinone IIA suppresses the progression of lung adenocarcinoma through regulating CCNA2-CDK2 complex and AURKA/PLK1 pathway. Sci. Rep. 2021, 11, 23681. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, Y.; Yang, C.; Simeone, D.M.; Sun, T.; DeGraff, D.J.; Tang, M.; Zhang, Y.; Wu, X. Dominant role of CDKN2B/p15INK4B of 9p21.3 tumor suppressor hub in inhibition of cell-cycle and glycolysis. Nat. Commun. 2021, 12, 2047. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, K.; Huang, J.; Wang, S.; Liu, Y.; Fan, G.; Liu, M.; Yang, G.; Wang, C.; Fan, P. CDKN2B is critical for verapamil-mediated reversal of doxorubicin resistance in hepatocellular carcinoma. Oncotarget 2017, 8, 110052–110063. [Google Scholar] [CrossRef]
- Stearns, M.E.; Wang, M. Synergistic Effects of the Green Tea Extract Epigallocatechin-3-gallate and Taxane in Eradication of Malignant Human Prostate Tumors. Transl. Oncol. 2011, 4, 147–156. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Fernández, A.; Ordóñez, R.; Reiter, R.J.; González-Gallego, J.; Mauriz, J.L. Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 2015, 59, 292–307. [Google Scholar] [CrossRef] [PubMed]
- Iurlaro, R.; Muñoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [PubMed]
- Mo, P.; Wang, H.; Lu, H.; Boyd, D.D.; Yan, C. MDM2 mediates ubiquitination and degradation of activating transcription factor 3. J. Biol. Chem. 2010, 285, 26908–26915. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Lu, D.; Hai, T.; Boyd, D.D. Activating transcription factor 3, a stress sensor, activates p53 by blocking its ubiquitination. EMBO J. 2005, 24, 2425–2435. [Google Scholar] [CrossRef]
- Muñoz-Guardiola, P.; Casas, J.; Megías-Roda, E.; Solé, S.; Perez-Montoyo, H.; Yeste-Velasco, M.; Erazo, T.; Diéguez-Martínez, N.; Espinosa-Gil, S.; Muñoz-Pinedo, C.; et al. The anti-cancer drug ABTL0812 induces ER stress-mediated cytotoxic autophagy by increasing dihydroceramide levels in cancer cells. Autophagy 2021, 17, 1349–1366. [Google Scholar] [CrossRef]
- Wang, X.; Gocek, E.; Novik, V.; Harrison, J.S.; Danilenko, M.; Studzinski, G.P. Inhibition of Cot1/Tlp2 oncogene in AML cells reduces ERK5 activation and up-regulates p27Kip1 concomitant with enhancement of differentiation and cell cycle arrest induced by silibinin and 1,25-dihydroxyvitamin D(3). Cell. Cycle 2010, 9, 4542–4551. [Google Scholar] [CrossRef]
- Li, T.; Su, L.; Zhong, N.; Hao, X.; Zhong, D.; Singhal, S.; Liu, X. Salinomycin induces cell death with autophagy through activation of endoplasmic reticulum stress in human cancer cells. Autophagy 2013, 9, 1057–1068. [Google Scholar] [CrossRef]
- Puissant, A.; Robert, G.; Fenouille, N.; Luciano, F.; Cassuto, J.; Raynaud, S.; Auberger, P. Resveratrol promotes autophagic cell death in chronic myelogenous leukemia cells via JNK-mediated p62/SQSTM1 expression and AMPK activation. Cancer Res. 2010, 70, 1042–1052. [Google Scholar] [CrossRef]
- Salazar, M.; Carracedo, A.; Salanueva, I.J.; Hernández-Tiedra, S.; Lorente, M.; Egia, A.; Vázquez, P.; Blázquez, C.; Torres, S.; García, S.; et al. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J. Clin. Investig. 2009, 119, 1359–1372. [Google Scholar] [CrossRef]
- Lin, S.; Yu-Lee, L.; Lin, S. Osteoblastic Factors in Prostate Cancer Bone Metastasis. Curr. Osteoporos Rep. 2018, 16, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Docherty, F.E.; Brown, H.K.; Reeves, K.J.; Fowles, A.C.M.; Ottewell, P.D.; Dear, T.N.; Holen, I.; Croucher, P.I.; Eaton, C.L. Prostate cancer cells preferentially home to osteoblast-rich areas in the early stages of bone metastasis: Evidence from in vivo models. J. Bone Miner. Res. 2014, 29, 2688–2696. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Mei, J.; Lu, J.; Wang, Y.; Hu, M.; Ma, F.; Long, H.; Qin, Z.; Tao, N. Ligustilide promotes apoptosis of cancer-associated fibroblasts via the TLR4 pathways. Food Chem. Toxicol. 2020, 135, 110991. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | |
| L-Glycine | 1200 mg |
| L-Arginine | 384 mg |
| L-Cysteine | 122.4 mg |
| Minerals | |
| Zinc sulfate | 48 mg |
| Glucosamine sulfate potassium chloride | 1200 mg |
| Manganese sulfate | 2.4 mg |
| Plant Extracts | |
| Licorice extract (Glycyrrhiza glabra) | 120 mg |
| Green tea extract (Canellia sinensis) | 15 mg |
| Cinnamon extract (Cinnamomum verum) | 1.8 mg |
| Vitamins | |
| Vitamin C | 72 mg |
| Vitamin B5 | 7.2 mg |
| Vitamin B6 | 2.4 mg |
| Vitamin B9 | 240 µg |
| Vitamin B12 | 1.2 µg |
| Gene | Sequence |
|---|---|
| ATF3 | Forward: AGAAAGAGTCGGAGAAGC |
| Reverse: TGAAGGTTGAGCATGTATATC | |
| DNAJB9 | Forward: TGCAGAAGCATATGAAACAC |
| Reverse: ACTAGTAAAAGCACTGTGTC | |
| ERO1LB | Forward: GGAGGAATTCCGATTACATTTC |
| Reverse: TTCCCCATAATCTGCATTTG | |
| CDK1 | Forward: ATGAGGTAGTAACACTCTGG |
| Reverse: CCTATACTCCAAATGCAACTG | |
| CDK2 | Forward: TGTTATCGCAAATGCTGC |
| Reverse: TCAAGAAGGCTATCAGAGTC | |
| CCNA2 | Forward: AGTATCATGGTGTTTCTCTCC |
| Reverse: AATTTGTACTTGGCCACAAC | |
| CDKN2B | Forward: GACTAGTGGAGAAGGTGC |
| Reverse: TCATCATGACCTGGATCG | |
| ERN1 | Forward: GAATAGAAAAGGAATCCCTGG |
| Reverse: TTCTTATTTCTCATGGCTCG |
| KEGG ID | Description |
|---|---|
| hsa04110 | Cell cycle |
| hsa03030 | DNA replication |
| hsa04115 | p53 signaling pathway |
| UPREGULATED | ||
| KEGG ID | Description | Genes |
| hsa01100 | Metabolic pathways | 50 |
| hsa05022 | Pathways of neurodegeneration | 15 |
| hsa04151 | PI3K-Akt signaling pathway | 13 |
| hsa05200 | Pathways in cancer | 13 |
| hsa05208 | Chemical carcinogenesis—reactive oxygen species | 12 |
| hsa05014 | Amyotrophic lateral sclerosis | 10 |
| hsa01240 | Biosynthesis of cofactors | 10 |
| hsa05010 | Alzheimer disease | 9 |
| hsa04141 | Protein processing in endoplasmic reticulum | 9 |
| hsa05168 | Herpes simplex virus 1 infection | 9 |
| hsa05165 | Human papillomavirus infection | 8 |
| hsa04510 | Focal adhesion | 8 |
| hsa04144 | Endocytosis | 8 |
| hsa05225 | Hepatocellular carcinoma | 8 |
| hsa04140 | Autophagy | 7 |
| hsa04210 | Apoptosis | 7 |
| hsa04150 | mTOR signaling pathway | 7 |
| hsa04115 | p53 signaling pathway | 7 |
| hsa04010 | MAPK signaling pathway | 7 |
| hsa05016 | Huntington disease | 7 |
| DOWNREGULATED | ||
| KEGG ID | Description | Genes |
| hsa04110 | Cell cycle | 18 |
| hsa03030 | DNA replication | 10 |
| hsa05200 | Pathways in cancer | 8 |
| hsa01100 | Metabolic pathways | 8 |
| hsa05166 | Human T-cell leukemia virus 1 infection | 6 |
| hsa04218 | Cellular senescence | 6 |
| hsa04914 | Progesterone-mediated oocyte maturation | 6 |
| hsa04114 | Oocyte meiosis | 5 |
| hsa05169 | Epstein–Barr virus infection | 5 |
| hsa04115 | p53 signaling pathway | 4 |
| hsa05207 | Chemical carcinogenesis—receptor activation | 4 |
| hsa05203 | Viral carcinogenesis | 4 |
| hsa04080 | Neuroactive ligand-receptor interaction | 4 |
| hsa03460 | Fanconi anemia pathway | 4 |
| hsa04611 | Platelet activation | 4 |
| hsa03430 | Mismatch repair | 4 |
| hsa05202 | Transcriptional misregulation in cancer | 4 |
| hsa05161 | Hepatitis B | 4 |
| hsa03440 | Homologous recombination | 4 |
| hsa05165 | Human papillomavirus infection | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez-Unzueta, I.; Benedicto, A.; Telleria, U.; Sanz, E.; Márquez, J. Improving the Antitumor Effect of Chemotherapy with Ocoxin as a Novel Adjuvant Agent to Treat Prostate Cancer. Nutrients 2023, 15, 2536. https://doi.org/10.3390/nu15112536
Hernandez-Unzueta I, Benedicto A, Telleria U, Sanz E, Márquez J. Improving the Antitumor Effect of Chemotherapy with Ocoxin as a Novel Adjuvant Agent to Treat Prostate Cancer. Nutrients. 2023; 15(11):2536. https://doi.org/10.3390/nu15112536
Chicago/Turabian StyleHernandez-Unzueta, Iera, Aitor Benedicto, Uxue Telleria, Eduardo Sanz, and Joana Márquez. 2023. "Improving the Antitumor Effect of Chemotherapy with Ocoxin as a Novel Adjuvant Agent to Treat Prostate Cancer" Nutrients 15, no. 11: 2536. https://doi.org/10.3390/nu15112536