
Investigation of Behavior and Plasma Levels of Corticosterone in Restrictive- and Ad Libitum-Fed Diet-Induced Obese Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets and Feeding Regimens
2.3. Behavioral Tests
2.3.1. Open Field (OF)
2.3.2. Elevated Plus Maze (EPM)
2.3.3. Social Interaction (SI)
2.3.4. Hotplate (HP)
2.4. Measurement of Plasma Corticosterone Concentration
2.5. Statistics
3. Results
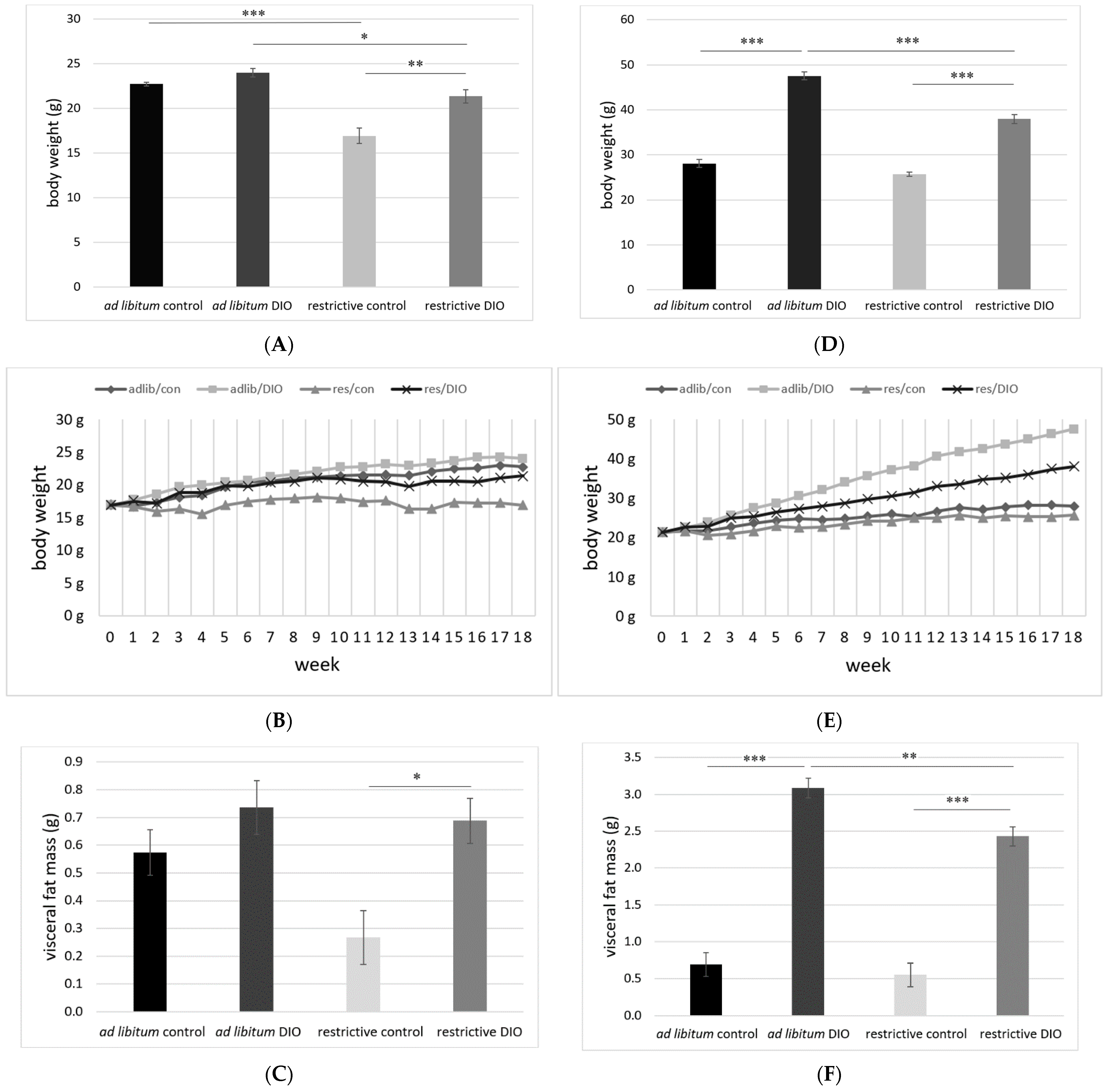
3.1. Body Weight
3.2. Open Field (OF)
3.3. Elevated Plus Maze (EPM)
3.4. Social Interaction (SI)
3.5. Hotplate (HP)
3.6. Plasma Corticosterone Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. The Top 10 Causes of Death. Available online: http://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 5 December 2018).
- World Health Organization. Overweight and Obesity. Available online: http://www.who.int/gho/ncd/risk_factors/overweight_text/en/ (accessed on 5 December 2018).
- Lutz, T.A.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharmacol. 2012, 58, 5–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, W.K. An overview of mongenic and syndromic obesities in humans. Pediatr. Blood Cancer 2012, 58, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Ichihara, S.; Yamada, Y. Genetic factors for human obesity. Cell Mol. Life Sci. 2008, 65, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Fellmann, L.; Nascimento, A.R.; Tibiriça, E.; Bousquet, P. Murine models for pharmacological studies of the metabolic syndrome. Pharmacol. Ther. 2013, 137, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Volynets, V. Nutritional influences of overfeeding on experimental outcomes in laboratory mice: Consequences for gut microbiota and other functional studies. Int. J. Med. Microbiol. 2016, 306, 328–333. [Google Scholar] [CrossRef]
- Hariri, N.; Thibault, L. High-fat diet-induced obesity in animal models. Nutr. Res. Rev. 2010, 23, 270–299. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Longo, V.D.; Harvie, M. Impact of intermittent fasting on health and disease processes. Ageing Res. Rev. 2017, 39, 46–58. [Google Scholar] [CrossRef]
- Ahmet, I.; Wan, R.; Mattson, M.P.; Lakatta, E.G.; Talan, M. Cardioprotection by intermittent fasting in rats. Circulation 2005, 112, 3115–3121. [Google Scholar] [CrossRef] [Green Version]
- Dinel, A.-L.; André, C.; Aubert, A.; Ferreira, G.; Layé, S.; Castanon, N. Cognitive and Emotional Alterations Are Related to Hippocampal Inflammation in a Mouse Model of Metabolic Syndrome. PLoS ONE 2011, 6, e24325. [Google Scholar] [CrossRef]
- André, C.; Dinel, A.-L.; Ferreira, G.; Layé, S.; Castanon, N. Diet-induced obesity progressively alters cognition, anxiety-like behavior and lipopolysaccharide-induced depressive-like behavior: Focus on brain indoleamine 2,3-dioxygenase activation. Brain Behav. Immun. 2014, 41, 10–21. [Google Scholar] [CrossRef]
- Balsevich, G.; Uribe, A.; Wagner, K.V.; Hartmann, J.; Santarelli, S.; Labermaier, C.; Schmidt, M.V. Interplay between diet-induced obesity and chronic stress in mice: Potential role of FKBP51. J. Endocrinol. 2014, 222, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, L.E.; Kitlinska, J.B.; Tilan, J.U.; Li, L.; Baker, S.B.; Johnson, M.D.; Lee, E.W.; Burnett, M.S.; Fricke, S.T.; Kvetnansky, R.; et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nat. Med. 2007, 13, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Krishnaveni, G.V.; Jones, A.; Veena, S.R.; Somashekara, R.; Karat, S.C.; Fall, C.H.D. Adiposity and Cortisol Response to Stress in Indian Adolescents. Indian Pediatr. 2018, 55, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Surkan, P.J.; Sakyi, K.S.; Hu, A.; Olinto, M.T.; Gonçalves, H.; Horta, B.L.; Gigante, D.P. Impact of stressful life events on central adiposity in the Pelotas Birth Cohort. Rev. Saude Publica 2018, 52, 61. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Jiménez, B.; Sámano, R.; Chinchilla-Ochoa, D.; Morales-Hernández, R.; Rodríguez-Ventura, A. Demographics and Psychological Factors Associated with Adiposity in Nurses. Int. J. Environ. Res. Public Health 2018, 15, 634. [Google Scholar] [CrossRef] [Green Version]
- Van der Valk, E.S.; Savas, M.; van Rossum, E.F.C. Stress and Obesity: Are There More Susceptible Individuals? Curr. Obes. Rep. 2018, 7, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Browne, C.A.; Hanke, J.; Rose, C.; Walsh, I.; Foley, T.; Clarke, G.; Schwegler, H.; Cryan, J.F.; Yilmazer-Hanke, D. Effect of acute swim stress on plasma corticosterone and brain monoamine levels in bidirectionally selected DxH recombinant inbred mouse strains differing in fear recall and extinction. Stress 2014, 17, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Shoji, H.; Miyakawa, T. Effects of test experience, closed-arm wall color, and illumination level on behavior and plasma corticosterone response in an elevated plus maze in male C57BL/6J mice: A challenge against conventional interpretation of the test. Mol. Brain 2021, 14, 149. [Google Scholar] [CrossRef]
- Boulangé, C.L.; Claus, S.P.; Chou, C.J.; Collino, S.; Montoliu, I.; Kochhar, S.; Holmes, E.; Rezzi, S.; Nicholson, J.K.; Dumas, M.E.; et al. Early metabolic adaptation in C57BL/6 mice resistant to high fat diet induced weight gain involves an activation of mitochondrial oxidative pathways. J. Proteome Res. 2013, 12, 1956–1968. [Google Scholar] [CrossRef]
- Peirson, S.N.; Brown, L.A.; Pothecary, C.A.; Benson, L.A.; Fisk, A.S. Light and the laboratory mouse. J. Neurosci. Methods 2018, 300, 26–36. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-like Behavior in Mice. JoVE 2015, 96, e52434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, R.J.; Dalvi, A. Anxiety, defence and the elevated plus-maze. Neurosci. Biobehav. Rev. 1997, 21, 801–810. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Cao, B.-J.; Dalvi, A.; Holmes, A. Animal models of anxiety: An ethological perspective. Braz. J. Med. Biol. Res. 1997, 30, 289–304. [Google Scholar] [CrossRef] [Green Version]
- Espejo, E.F. Structure of the mouse behaviour on the elevated plus-maze test of anxiety. Behav. Brain Res. 1997, 86, 105–112. [Google Scholar] [CrossRef]
- Mulder, G.B.; Pritchett-Corning, K. Rodent Analgesiometry: The Hot Plate, Tail Flick and von Frey Hairs. 2018. Available online: https://www.researchgate.net/publication/8531665_Rodent_analgesiometry_The_hot_plate_tail_flick_and_Von_Frey_hairs (accessed on 21 September 2019).
- Rat and Mice Weights Animal Resources Centre. 2021. Available online: https://www.arc.wa.gov.au/?page_id=125 (accessed on 21 May 2021).
- BALB/c Mouse Charles River. 2020. Available online: https://www.criver.com/products-services/find-model/balbc-mouse?region=23 (accessed on 20 January 2020).
- Holmes, M.M.; Mistlberger, R.E. Food anticipatory activity and photic entrainment in food-restricted BALB/c mice. Physiol. Behav. 2000, 68, 655–666. [Google Scholar] [CrossRef]
- Gelegen, C.; Collier, D.A.; Campbell, I.C.; Oppelaar, H.; Kas, M.J.H. Behavioral, physiological, and molecular differences in response to dietary restriction in three inbred mouse strains. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E574–E581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelegen, C.; Collier, D.A.; Campbell, I.C.; Oppelaar, H.; van den Heuvel, J.; Adan, R.A.H.; Kas, M.J. Difference in susceptibility to activity-based anorexia in two inbred strains of mice. Eur. Neuropsychopharmacol. 2007, 17, 199–205. [Google Scholar] [CrossRef]
- Dos Santos Guilherme, M.; Tsoutsouli, T.; Chongtham, M.C.; Winter, J.; Gerber, S.; Müller, M.B.; Endres, K. Selective targeting of chronic social stress-induced activated neurons identifies neurogenesis-related genes to be associated with resilience in female mice. Psychoneuroendocrinology 2022, 139, 105700. [Google Scholar] [CrossRef]
- Moraes, R.C.; Blondet, A.; Birkenkamp-Demtroeder, K.; Tirard, J.; Orntoft, T.F.; Gertler, A.; Durand, P.; Naville, D.; Bégeot, M. Study of the alteration of gene expression in adipose tissue of diet-induced obese mice by microarray and reverse transcription-polymerase chain reaction analyses. Endocrinology 2003, 144, 4773–4782. [Google Scholar] [CrossRef] [Green Version]
- Buchenauer, T.; Behrendt, P.; Bode, F.J.; Horn, R.; Brabant, G.; Stephan, M.; Nave, H. Diet-induced obesity alters behavior as well as serum levels of corticosterone in F344 rats. Physiol. Behav. 2009, 98, 563–569. [Google Scholar] [CrossRef]
- Olson, L.K.; Tan, Y.; Zhao, Y.; Aupperlee, M.D.; Haslam, S.Z. Pubertal exposure to high fat diet causes mouse strain-dependent alterations in mammary gland development and estrogen responsiveness. Int. J. Obes. 2010, 34, 1415–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boi, S.K.; Buchta, C.M.; Pearson, N.A.; Francis, M.B.; Meyerholz, D.K.; Grobe, J.L.; Norian, L.A. Obesity alters immune and metabolic profiles: New insight from obese-resistant mice on high-fat diet. Obesity 2016, 24, 2140–2149. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Hallahan, N.L.; Brown, S.H.; Liu, M.; Mitchell, T.W.; Cooney, G.J.; Turner, N. Mouse strain-dependent variation in obesity and glucose homeostasis in response to high-fat feeding. Diabetologia 2013, 56, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Nunez, I.; Caluag, T.; Kirby, K.; Rudick, C.N.; Dziarski, R.; Gupta, D. Nod2 and Nod2-regulated microbiota protect BALB/c mice from diet-induced obesity and metabolic dysfunction. Sci. Rep. 2017, 7, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, J.; Ericsson, M.; Joibari, M.M.; Anderson, F.; Carlsson, L.; Nilsson, S.K.; Sjödin, A.; Burén, J. A low-carbohydrate high-fat diet decreases lean mass and impairs cardiac function in pair-fed female C57BL/6J mice. Nutr. Metab. 2016, 13, 557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, A. Animal models of anxiety: Do I need multiple tests? Trends Pharmacol. Sci. 2008, 29, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Di Biase, S.; Longo, V.D. Dietary Interventions, Cardiovascular Aging, and Disease: Animal Models and Human Studies. Circ. Res. 2016, 118, 1612–1625. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; He, C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018, 44, 38–50. [Google Scholar] [CrossRef]
- Barrios-Correa, A.A.; Estrada, J.A.; Contreras, I. Leptin Signaling in the Control of Metabolism and Appetite: Lessons from Animal Models. J. Mol. Neurosci. 2018, 66, 390–402. [Google Scholar] [CrossRef]
- Jang, M.; Mistry, A.; Swick, A.G.; Romsos, D.R. Leptin rapidly inhibits hypothalamic neuropeptide Y secretion and stimulates corticotropin-releasing hormone secretion in adrenalectomized mice. J. Nutr. 2000, 130, 2813–2820. [Google Scholar] [CrossRef] [Green Version]
- Karl, T.; Duffy, L.; Herzog, H. Behavioural profile of a new mouse model for NPY deficiency. Eur. J. Neurosci. 2008, 28, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Brownlow, B.S.; Petro, A.; Feinglos, M.N.; Surwit, R.S. The role of motor activity in diet-induced obesity in C57BL/6J mice. Physiol. Behav. 1996, 60, 37–41. [Google Scholar] [CrossRef]
- Blancas-Velazquez, A.; Mendoza, J.; Garcia, A.N.; La Fleur, S.E. Diet-Induced Obesity and Circadian Disruption of Feeding Behavior. Front. Neurosci. 2017, 11, 350. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Torvinen, S.; Silvennoinen, M.; Rinnankoski-Tuikka, R.; Kainulainen, H.; Morko, J.; Peng, Z.; Kujala, U.M.; Rahkila, P.; Suominen, H. Effects of diet-induced obesity and voluntary wheel running on bone properties in young male C57BL/6J mice. Calcif. Tissue Int. 2010, 86, 411–419. [Google Scholar] [CrossRef]
- Miczek, K.A.; Maxson, S.C.; Fish, E.W.; Faccidomo, S. Aggressive behavioral phenotypes in mice. Behav. Brain Res. 2001, 125, 167–181. [Google Scholar] [CrossRef]
- Matsumoto, K.; Pinna, G.; Puia, G.; Guidotti, A.; Costa, E. Social isolation stress-induced aggression in mice: A model to study the pharmacology of neurosteroidogenesis. Stress 2005, 8, 85–93. [Google Scholar] [CrossRef]
- Lister, R.G.; Hilakivi, L.A. The effects of novelty, isolation, light and ethanol on the social behavior of mice. Psychopharmacology 1988, 96, 181–187. [Google Scholar] [CrossRef]
- Võikar, V.; Polus, A.; Vasar, E.; Rauvala, H. Long-term individual housing in C57BL/6J and DBA/2 mice: Assessment of behavioral consequences. Genes Brain Behav. 2005, 4, 240–252. [Google Scholar] [CrossRef]
- Surwit, R.S.; Kuhn, C.M.; Cochrane, C.; McCubbin, J.A.; Feinglos, M.N. Diet-induced type II diabetes in C57BL/6J mice. Diabetes 1988, 37, 1163–1167. [Google Scholar] [CrossRef]
- Rossi, H.L.; Luu, A.K.S.; Kothari, S.D.; Kuburas, A.; Neubert, J.K.; Caudle, R.M.; Recober, A. Effects of diet-induced obesity on motivation and pain behavior in an operant assay. Neuroscience 2013, 235, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.-J.; Han, W.; Cao, C.-Q.; Mao-Ying, Q.-L.; Mi, W.-L.; Wang, Y.-Q. Peripheral Leptin Signaling Mediates Formalin-Induced Nociception. Neurosci. Bull. 2018, 34, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Fulton, S. Diet-induced obesity promotes depressive-like behaviour that is associated with neural adaptations in brain reward circuitry. Int. J. Obes. 2013, 37, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, S.; Miao, Y.-L.; Jiao, G.-Z.; Sun, M.-J.; Li, H.; Lin, J.; Luo, M.-J.; Tan, J.-H. Dynamics and Correlation of Serum Cortisol and Corticosterone under Different Physiological or Stressful Conditions in Mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef] [PubMed]
- Kamakura, R.; Kovalainen, M.; Leppäluoto, J.; Herzig, K.-H.; Mäkelä, K.A. The effects of group and single housing and automated animal monitoring on urinary corticosterone levels in male C57BL/6 mice. Physiol. Rep. 2016, 4, e12703. [Google Scholar] [CrossRef] [PubMed]
- Herane Vives, A.; Angel, V.; de Papadopoulos, A.; Strawbridge, R.; Wise, T.; Young, A.H.; Arnone, D.; Cleare, A.J. The relationship between cortisol, stress and psychiatric illness: New insights using hair analysis. J. Psychiatr. Res. 2015, 70, 38–49. [Google Scholar] [CrossRef]
- Dlamini, S.N.; Lombard, Z.; Micklesfield, L.K.; Crowther, N.; Norris, S.A.; Snyman, T.; Crawford, A.A.; Walker, B.R.; Goedecke, J.H. Glucocorticoids associate with cardiometabolic risk factors in black South Africans. Endocr. Connect. 2021, 10, 873–884. [Google Scholar] [CrossRef]
- Spielmann, J.; Mattheis, L.; Jung, J.-S.; Rauße, H.; Glaß, M.; Bähr, I.; Quandt, D.; Oswald, J.; Kielstein, H. Effects of obesity on NK cells in a mouse model of postmenopausal breast cancer. Sci. Rep. 2020, 10, 20606. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allweyer, M.; Emde, M.; Bähr, I.; Spielmann, J.; Bieramperl, P.; Naujoks, W.; Kielstein, H. Investigation of Behavior and Plasma Levels of Corticosterone in Restrictive- and Ad Libitum-Fed Diet-Induced Obese Mice. Nutrients 2022, 14, 1746. https://doi.org/10.3390/nu14091746
Allweyer M, Emde M, Bähr I, Spielmann J, Bieramperl P, Naujoks W, Kielstein H. Investigation of Behavior and Plasma Levels of Corticosterone in Restrictive- and Ad Libitum-Fed Diet-Induced Obese Mice. Nutrients. 2022; 14(9):1746. https://doi.org/10.3390/nu14091746
Chicago/Turabian StyleAllweyer, Martin, Matthias Emde, Ina Bähr, Julia Spielmann, Philipp Bieramperl, Wiebke Naujoks, and Heike Kielstein. 2022. "Investigation of Behavior and Plasma Levels of Corticosterone in Restrictive- and Ad Libitum-Fed Diet-Induced Obese Mice" Nutrients 14, no. 9: 1746. https://doi.org/10.3390/nu14091746