
Molecular Iodine Supplement Prevents Streptozotocin-Induced Pancreatic Alterations in Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Pancreatitis and Insulitis Determination
2.4. Morphology, Vasculature and iNOS Presence
2.5. Gene Expression
2.6. Protein Extraction and Western Blotting
2.7. Lipid Peroxidation in Pancreas
2.8. Statistical Analyses
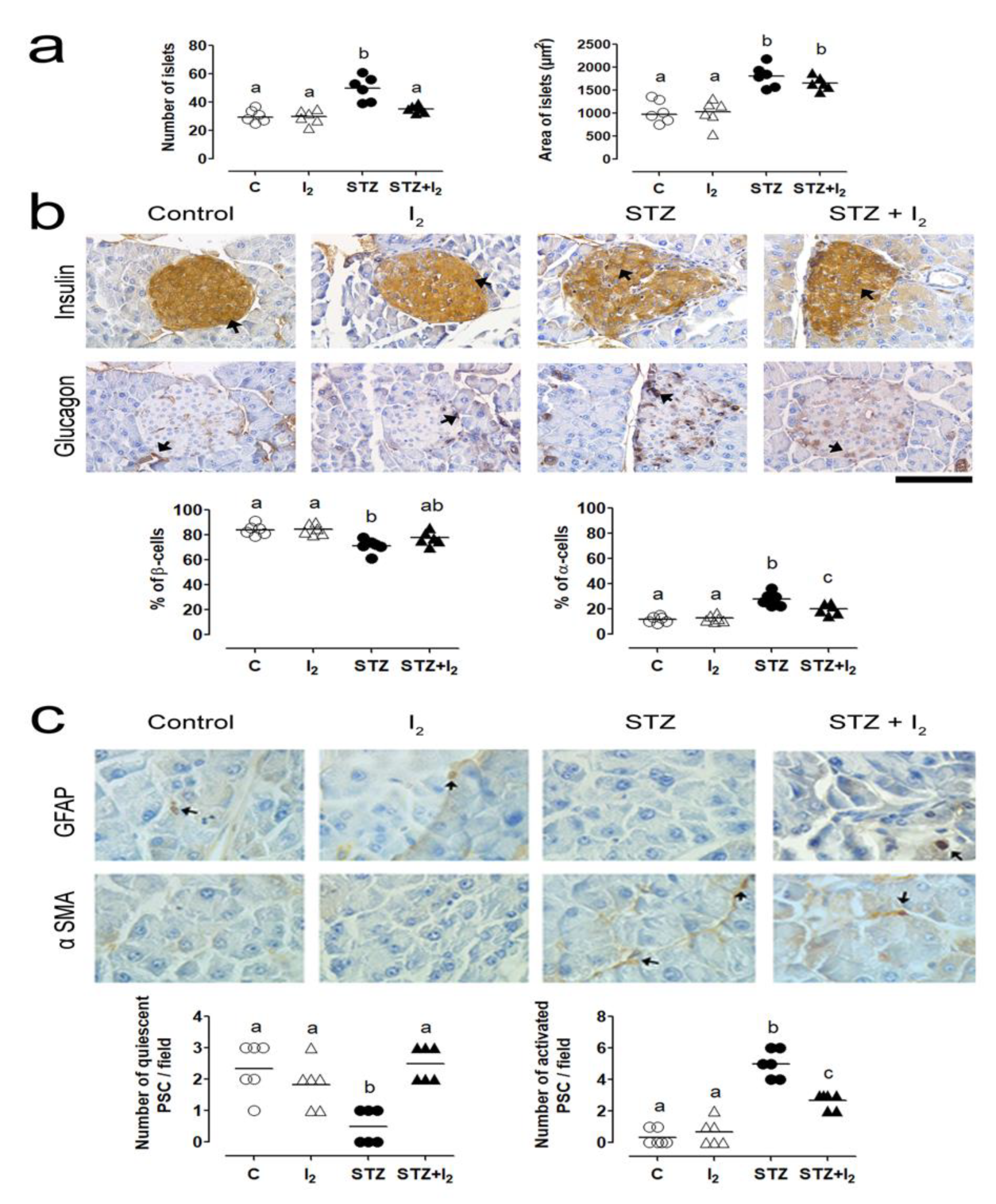
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Criddle, D.N. Reactive oxygen species Ca2+ stores and acute pancreatitis; a step closer to therapy? Cell Calcium 2016, 60, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, D.K.; Korc, M.; Petersen, G.M.; Eibl, G.; Li, D.; Rickels, M.R.; Suresh, T.C.; Abbruzzese, J.L. Diabetes, pancreatogenic diabetes, and pancreatic cancer. Diabetes 2017, 66, 1103–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, G.; Hong, W.; Guo, Y.; Bai, Y.; Chen, B. Molecular mechanism of pancreatic stellate cells activation in chronic pancreatitis and pancreatic cancer. J. Cancer 2020, 11, 1505–1515. [Google Scholar] [CrossRef] [Green Version]
- Marzoq, A.J.; Mustafa, S.A.; Heidrich, L.; Hoheisel, J.D.; Alhamdani, M.S.S. Impact of the secretome of activated pancreatic stellate cells on growth and differentiation of pancreatic tumour cells. Sci. Rep. 2019, 9, 5303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, R.; Jia, K.; Wang, J.; Yang, L.; Wang, Y.; Gao, L.; Hao, J.A. Rising star in pancreatic diseases: Pancreatic stellate cells. Front. Physiol. 2018, 9, 754–759. [Google Scholar] [CrossRef]
- Masamune, A.; Suzuki, N.; Kikuta, K.; Satoh, M.; Satoh, K.; Shimosegawa, T. Curcumin blocks activation of pancreatic stellate cells. J. Cell. Biochem. 2006, 97, 1080–1093. [Google Scholar] [CrossRef]
- Lin, Z.; Zheng, L.C.; Zhang, H.J.; Tsang, S.W.; Bian, Z.X. Anti-fibrotic effects of phenolic compounds on pancreatic stellate cells. BMC Complement. Altern. Med. 2015, 15, 259–562. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Cheng, L.; Jiang, Z.; Chen, K.; Zhou, C.; Sun, L.; Cao, J.; Quian, W.; Li, J.; Shan, T.; et al. Resveratrol inhibits ROS-promoted activation and glycolysis of pancreatic stellate cells via suppression of miR-21. Oxidative Med. Cell. Longev. 2018, 2018, 1346958. [Google Scholar] [CrossRef]
- Xia, Y.; Xiao, H.T.; Liu, K.; Zhang, H.J.; Tsang, S.W. Resveratrol ameliorates the severity of fibrogenesis in mice with experimental chronic pancreatitis. Mol. Nutr. Food Res. 2018, 62, 1700561–1700565. [Google Scholar] [CrossRef]
- Booth, D.M.; Mukherjee, R.; Sutton, R.; Criddle, D.N. Calcium and Reactive Oxygen Species in Acute Pancreatitis: Friend or Foe? Antioxid. Redox Signal. 2011, 15, 2683–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Antioxidant supplements for prevention of mortality in healthy participants and patients with various diseases. Cochrane Database Syst. Rev. 2008, 3, CD007176. [Google Scholar]
- Aceves, C.; Mendieta, I.; Anguiano, B.; Delgado-Gonzalez, E. Molecular iodine has extrathyroidal effects as an antioxidant, differentiator, and immunomodulator. Int. J. Mol. Sci. 2021, 22, 1228. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, Y.; Delgado, G.; Carabez, A.; Anguiano, B.; Aceves, C. Iodine and doxorubicin, a good combination for mammary cancer treatment: Antineoplastic adjuvancy, chemoresistance inhibition, and cardioprotection. Mol. Cancer 2013, 12, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, L.J.; Chen, G.Y.; Zhao, W.; Qiu, M.C. Effects of supplementation of different kinds of iodine on the antioxidative ability of retina in iodine deficient rats. Chin. J. Ophthalmol. 2003, 39, 495–498. [Google Scholar]
- Garcia-Solis, P.; Alfaro, Y.; Anguiano, B.; Delgado, G.; Guzman, R.C.; Nandi, S.; Aceves, C. Inhibition of N-methyl-N-nitrosourea-induced mammary carcinogenesis by molecular iodine (I2) but not by iodide (I−) treatment: Evidence that I2 prevents cancer promotion. Mol. Cell. Endocrinol. 2005, 236, 49–57. [Google Scholar] [CrossRef]
- Li, Q.; Mair, C.; Schedle, K.; Hellmayr, I.; Windisch, W. Effects of varying dietary iodine supplementation levels as iodide or iodate on thyroid status as well as mRNA expression and enzyme activity of antioxidative enzymes in tissues of grower/finisher pigs. Eur. J. Nutr. 2013, 52, 161–168. [Google Scholar] [CrossRef]
- Greenwald, B.Y.M.; Frusic-Zlotkin, M.; Soroka, Y.; Ben-Sasson, S.; Bianco-Peled, H.; Kohen, R. A novel role of topical iodine in skin: Activation of the Nrf2 pathway. Free Radic. Biol. Med. 2017, 104, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Nava-Villalba, M.; Nuñez-Anita, R.E.; Bontempo, A.; Aceves, C. Activation of peroxisome proliferator-activated receptor gamma is crucial for antitumoral effects of 6-iodolactone. Mol. Cancer 2015, 14, 168–172. [Google Scholar] [CrossRef] [Green Version]
- Mendieta, I.; Nuñez-Anita, R.E.; Nava-Villalba, M.; Zambrano-Estrada, X.; Delgado-Gonzalez, E.; Anguiano, B.; Aceves, C. Molecular iodine exerts antineoplastic effects by diminishing proliferation and invasive potential and activating the immune response in mammary cancer xenografts. BMC Cancer 2019, 19, 261–266. [Google Scholar] [CrossRef]
- Hartoft-Nielsen, M.L.; Rasmussen, A.K.; Bock, T.; Feldt-Rasmussen, U.; Kaas, A.; Buschard, K. Iodine and triiodothyronine reduce the incidence of type 1 diabetes mellitus in the autoimmune prone BB rats. Autoimmunity 2009, 42, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Castelan, J.; Delgado-Gonzalez, E.; Rodriguez-Benitez, E.; Castelan, F.; Aceves, C.; Cuevas-Romero, E. Protective effect of moderated dose of iodine in pancreatic alterations during hypothyroidism. In Proceedings of the Endocrine Meeting, New Orleans, LA, USA, 23–26 March 2019. [Google Scholar]
- Moreno, C.; Nicaise, C.; Gustot, T.; Quertinmont, E.; Nagy, N.; Parmentier, M.; Louis, H.; Deviere, J. Chemokine receptor CCR5 deficiency exacerbates cerulein-induced acute pancreatitis in mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 2006, 291, G1089–G1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibashe-Ahmed, M.; Brioudes, E.; Reith, W.; Bosco, D.; Berney, T. Toll-like receptor 4 inhibition prevents autoimmune diabetes in NOD mice. Sci. Rep. 2019, 9, 19350. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.H.; Chu, C.H.; Yu, J.C.; Chuang, W.C.; Lin, G.J.; Chen, P.L.; Chou, F.C.; Chau, L.Y.; Sytwu, H.K. Transgenic expression of haemoxygenase-1 in pancreatic beta cells protects non-obese mice used as a model of diabetes from autoimmune destruction and prolongs graft survival following islet transplantation. Diabetologia 2010, 53, 2389–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Yan, L.J. Streptozotocin-induced type 1 diabetes in rodents as a model for studying mitochondrial mechanisms of diabetic β cell glucotoxicity. Diabetes Metab. Syndr. Obes. Targets Ther. 2015, 8, 181–188. [Google Scholar]
- Furman, B.L. Streptozotocin induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5–47. [Google Scholar] [CrossRef] [PubMed]
- Le May, C.K.; Chu, M.; Hu, C.S.; Ortega, C.S.; Simpson, E.R.; Korach, K.S.; Tsai, M.J.; Mauvais-Jarvis, F. Estrogens protect pancreatic beta-cells from apoptosis and prevent insulin-deficient diabetes mellitus in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 9232–9237. [Google Scholar] [CrossRef] [Green Version]
- Maric-Bilkan, C. Sex differences in diabetic kidney disease. Mayo Clin. Proc. 2019, 95, 587–599. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Li, W.; Shotorbani, Y.P.; Dubansky, H.B.; Huang, L.; Chaudhari, S.; Wu, P.; Wang, L.A.; Ryou, M.G.; Zhou, Z.; et al. Comparison of diabetic nephropathy between male and female eNOS(−/−) db/db mice. Am. J. Physiol. Ren. Physiol. Mice 2019, 316, F889–F897. [Google Scholar] [CrossRef]
- Tian, L.; Nikolic-Paterson, D.J.; Tesch, G.H. Establishing equivalent diabetes in male and female Nos3-deficient mice results in a comparable onset of diabetic kidney injury. Physiol. Rep. 2019, 7, e14197. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, Q.; Zhang, M.; Yin, T.; Xu, R.; Xiao, W.; Wu, J.; Deng, B.; Gao, X.; Gong, W.; et al. Isoliquiritigenin ameliorates acute pancreatitis in mice via inhibition of oxidative stress and modulation of the Nrf2/HO-1 pathway. Oxidative Med. Cell. Longev. 2018, 17, 7161592. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Brunmeir, R.; Xu, F. Functional regulation of PPARs through post-translational modifications. Int. J. Mol. Sci. 2018, 19, 1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaster, R.; Lichte, P.; Fitzner, B.; Brock, P.; Glass, A.; Karopka, T.; Gierl, L.; Koczan, D.; Thiesen, H.J.; Sparmann, G.; et al. Peroxisome proliferator-activated receptor gamma overexpression inhibits pro-fibrogenic activities of immortalised rat pancreatic stellate cells. J. Cell. Mol. Med. 2005, 9, 670–682. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, H.; Jing, Q.; Yang, Y.; Xue, D.; Hao, C.; Zhang, W. Regulation of pancreatic fibrosis by acinar cell-derived exosomal mir-130a-3p via targeting of stellate cell PPARγ. J. Inflamm. Res. 2021, 14, 461–477. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Castelán, J.; Delgado-González, E.; Varela-Floriano, V.; Anguiano, B.; Aceves, C. Molecular Iodine Supplement Prevents Streptozotocin-Induced Pancreatic Alterations in Mice. Nutrients 2022, 14, 715. https://doi.org/10.3390/nu14030715
Rodríguez-Castelán J, Delgado-González E, Varela-Floriano V, Anguiano B, Aceves C. Molecular Iodine Supplement Prevents Streptozotocin-Induced Pancreatic Alterations in Mice. Nutrients. 2022; 14(3):715. https://doi.org/10.3390/nu14030715
Chicago/Turabian StyleRodríguez-Castelán, Julia, Evangelina Delgado-González, Valentin Varela-Floriano, Brenda Anguiano, and Carmen Aceves. 2022. "Molecular Iodine Supplement Prevents Streptozotocin-Induced Pancreatic Alterations in Mice" Nutrients 14, no. 3: 715. https://doi.org/10.3390/nu14030715