

Indole Acetic Acid Exerts Anti-Depressive Effects on an Animal Model of Chronic Mild Stress
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
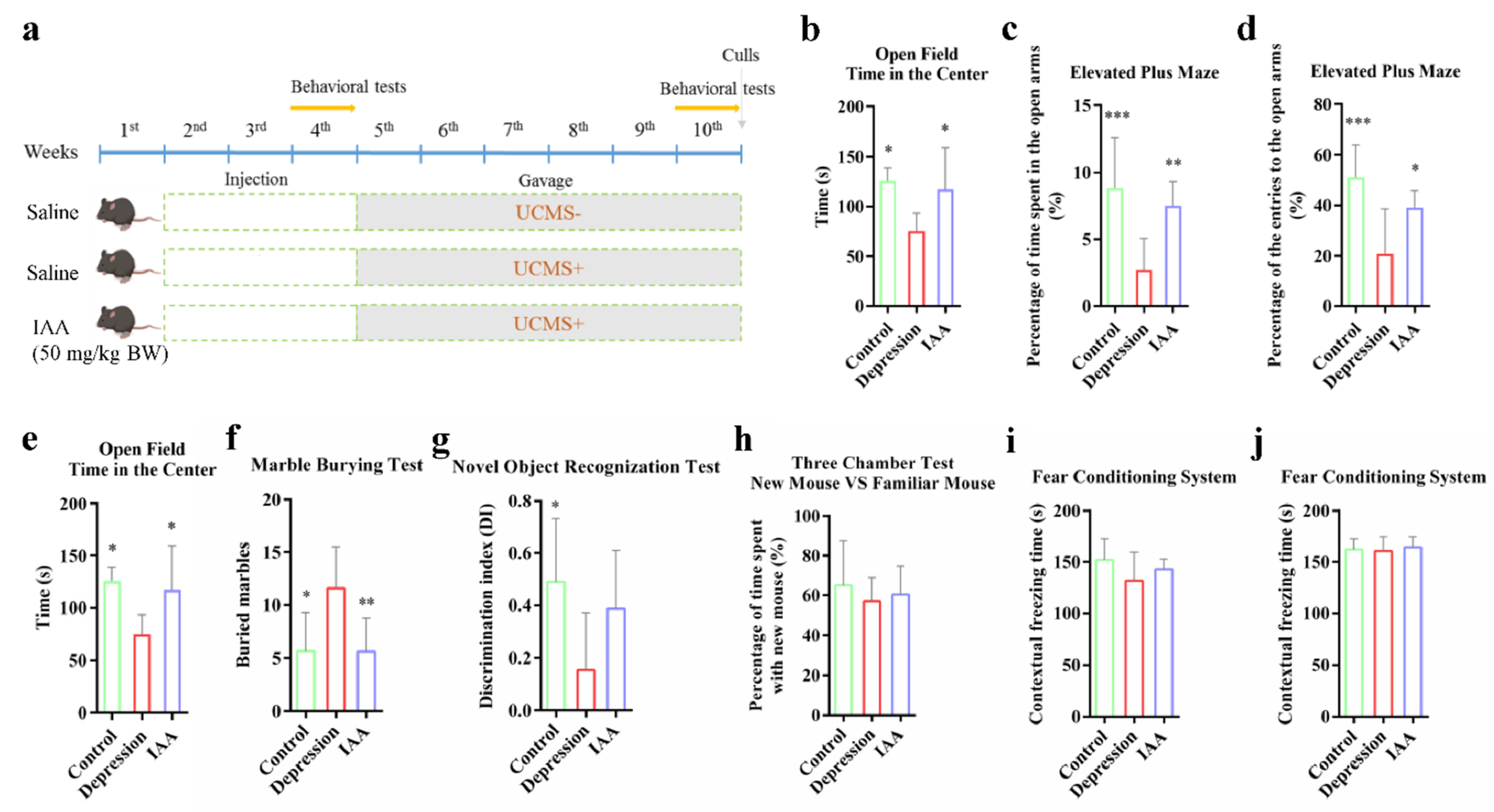
2.1. Animal Design
2.2. Behavioural Assays
2.3. UPLC-MS Analysis of Trp-Indole Derivatives and Neurotransmitters
2.4. SCFAs Analysis
2.5. Quantitative Real-Time PCR
2.6. Gut Microbiota Profiling
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Statistical Analysis
3. Results
3.1. IAA Alters the Anxiety- and Depression-Like Behaviours Induced by UCMS
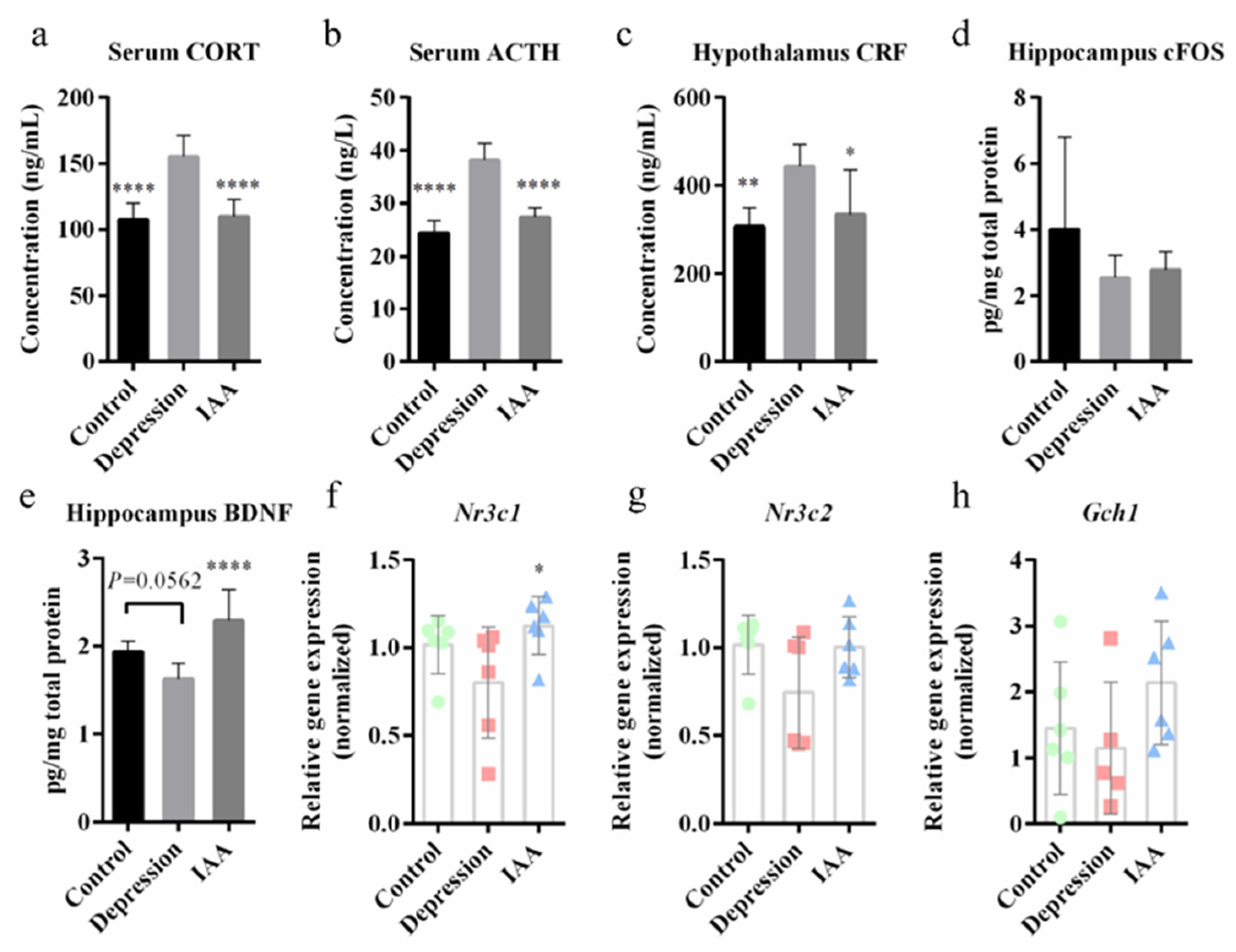
3.2. IAA Treatment Affects the HPA Axis and c-Fos/BDNF Expression
3.3. Administration of IAA Regulates Dopamine and the Serotonergic System
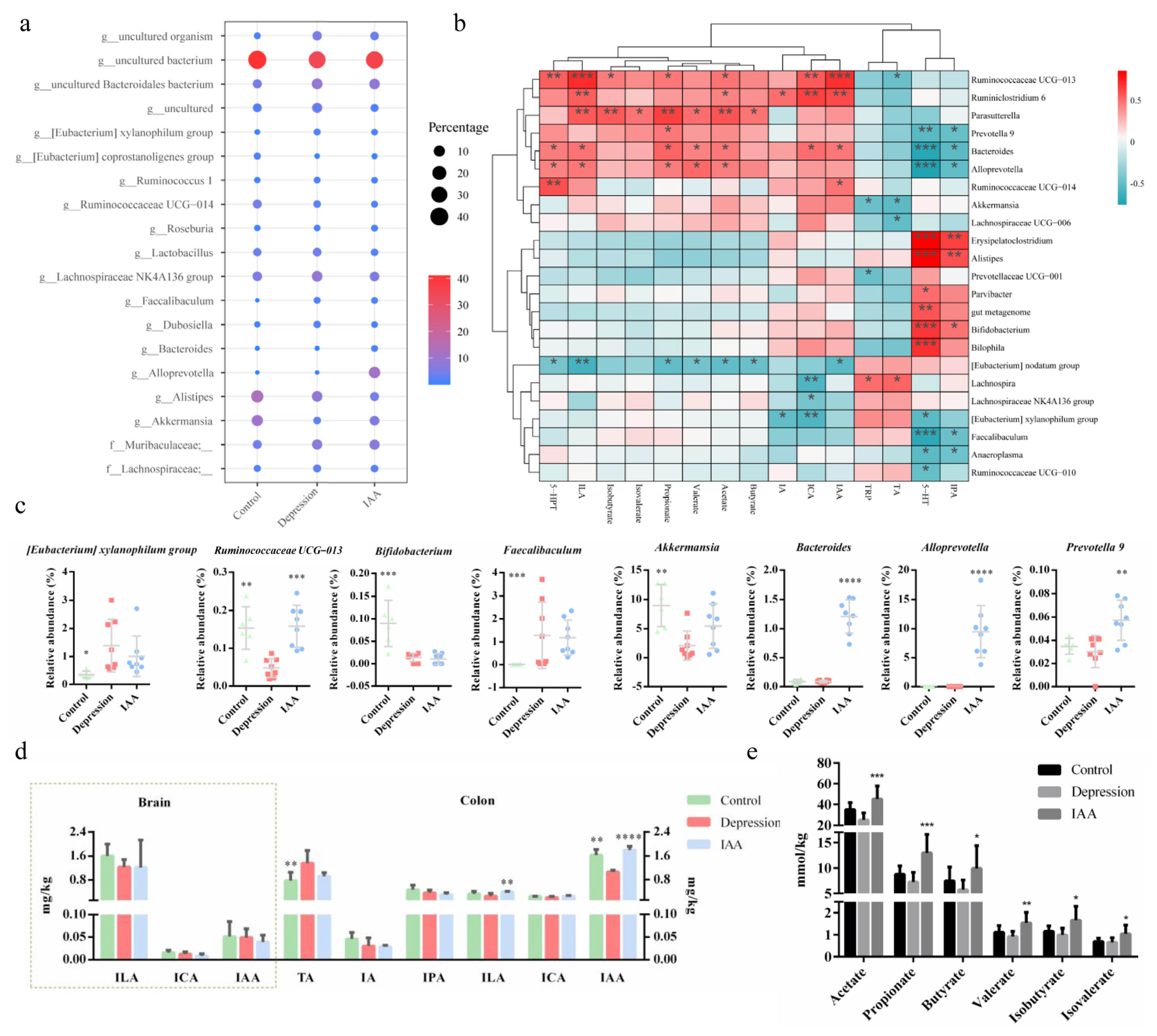
3.4. The Gut Microbiota and SCFA Concentrations Are Influenced by IAA
3.5. IAA Affects the Tryptophan–Indoles Metabolism Pathways in the Circulation and the Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Joodaki, M.; Radahmadi, M.; Alaei, H. Comparing the Therapeutic Effects of Crocin, Escitalopram and Co-Administration of Escitalopram and Crocin on Learning and Memory in Rats with Stress-Induced Depression. Malays. J. Med Sci. 2021, 28, 50. [Google Scholar] [CrossRef]
- Cruz-Pereira, J.S.; Rea, K.; Nolan, Y.M.; O’Leary, O.F.; Dinan, T.G.; Cryan, J.F. Depression’s unholy trinity: Dysregulated stress, immunity, and the microbiome. Annu. Rev. Psychol. 2020, 71, 49–78. [Google Scholar] [CrossRef]
- Manji, H.K.; Drevets, W.C.; Charney, D.S. The cellular neurobiology of depression. Nat. Med. 2001, 7, 541–547. [Google Scholar] [CrossRef]
- Frolkis, A.D.; A Vallerand, I.; Shaheen, A.-A.; Lowerison, M.W.; Swain, M.G.; Barnabe, C.; Patten, S.B.; Kaplan, G.G. Depression increases the risk of inflammatory bowel disease, which may be mitigated by the use of antidepressants in the treatment of depression. Gut 2019, 68, 1606–1612. [Google Scholar] [CrossRef]
- Chevalier, G.; Siopi, E.; Guenin-Macé, L.; Pascal, M.; Laval, T.; Rifflet, A.; Boneca, I.G.; Demangel, C.; Colsch, B.; Pruvost, A.; et al. Effect of gut microbiota on depressive-like behaviors in mice is mediated by the endocannabinoid system. Nat. Commun. 2020, 11, 6363. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hua, D.; Wang, Q.; Yang, L.; Wang, X.; Luo, A.; Yang, C. The role of bacteria and its derived metabolites in chronic pain and depression: Recent findings and research progress. Int. J. Neuropsychopharmacol. 2020, 23, 26–41. [Google Scholar] [CrossRef]
- Settanni, C.R.; Ianiro, G.; Bibbò, S.; Cammarota, G.; Gasbarrini, A. Gut microbiota alteration and modulation in psychiatric disorders: Current evidence on fecal microbiota transplantation. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 109, 110258. [Google Scholar] [CrossRef]
- Paiva, I.H.R.; Duarte-Silva, E.; Peixoto, C.A. The role of prebiotics in cognition, anxiety, and depression. Eur. Neuropsychopharmacol. 2020, 34, 1–18. [Google Scholar] [CrossRef]
- Tian, P.; Zou, R.; Wang, L.; Chen, Y.; Qian, X.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; Wang, G.; et al. Multi-Probiotics ameliorate Major depressive disorder and accompanying gastrointestinal syndromes via serotonergic system regulation. J. Adv. Res. 2022. [Google Scholar] [CrossRef]
- Li, C.-C.; Jiang, N.; Gan, L.; Zhao, M.-J.; Chang, Q.; Liu, X.-M.; Pan, R.-L. Peripheral and cerebral abnormalities of the tryptophan metabolism in the depression-like rats induced by chronic unpredicted mild stress. Neurochem. Int. 2020, 138, 104771. [Google Scholar] [CrossRef]
- Tian, P.; Chen, Y.; Zhu, H.; Wang, L.; Qian, X.; Zou, R.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; et al. Bifidobacterium breve CCFM1025 attenuates major depression disorder via regulating gut microbiome and tryptophan metabolism: A randomized clinical trial. Brain Behav. Immun. 2022, 100, 233–241. [Google Scholar] [CrossRef]
- Wang, D.; Wu, J.; Zhu, P.; Xie, H.; Lu, L.; Bai, W.; Pan, W.; Shi, R.; Ye, J.; Xia, B.; et al. Tryptophan-rich diet ameliorates chronic unpredictable mild stress induced depression-and anxiety-like behavior in mice: The potential involvement of gut-brain axis. Food Res. Int. 2022, 157, 111289. [Google Scholar] [CrossRef]
- Jaglin, M.; Rhimi, M.; Philippe, C.; Pons, N.; Bruneau, A.; Goustard, B.; Daugé, V.; Maguin, E.; Naudon, L.; Rabot, S. Indole, a signaling molecule produced by the gut microbiota, negatively impacts emotional behaviors in rats. Front. Neurosci. 2018, 12, 216. [Google Scholar] [CrossRef]
- Mir, H.-D.; Milman, A.; Monnoye, M.; Douard, V.; Philippe, C.; Aubert, A.; Castanon, N.; Vancassel, S.; Guérineau, N.C.; Naudon, L.; et al. The gut microbiota metabolite indole increases emotional responses and adrenal medulla activity in chronically stressed male mice. Psychoneuroendocrinology 2020, 119, 104750. [Google Scholar] [CrossRef]
- Eliwa, H.; Brizard, B.; Le Guisquet, A.-M.; Hen, R.; Belzung, C.; Surget, A. Adult neurogenesis augmentation attenuates anhedonia and HPA axis dysregulation in a mouse model of chronic stress and depression. Psychoneuroendocrinology 2021, 124, 105097. [Google Scholar] [CrossRef]
- Wei, G.Z.; Martin, K.A.; Xing, P.Y.; Agrawal, R.; Whiley, L.; Wood, T.K.; Hejndorf, S.; Ng, Y.Z.; Low, J.Z.Y.; Rossant, J.; et al. Tryptophan-metabolizing gut microbes regulate adult neurogenesis via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2021, 118, e2021091118. [Google Scholar] [CrossRef]
- Serger, E.; Luengo-Gutierrez, L.; Chadwick, J.S.; Kong, G.; Zhou, L.; Crawford, G.; Danzi, M.C.; Myridakis, A.; Brandis, A.; Bello, A.T.; et al. The gut metabolite indole-3 propionate promotes nerve regeneration and repair. Nature 2022, 607, 585–592. [Google Scholar] [CrossRef]
- Anderson, G.M.; Gerner, R.H.; Cohen, D.J.; Fairbanks, L. Central tryptamine turnover in depression, schizophrenia, and anorexia: Measurement of indoleacetic acid in cerebrospinal fluid. Biol. Psychiatry 1984, 19, 1427–1435. [Google Scholar]
- Pu, J.; Liu, Y.; Gui, S.; Tian, L.; Yu, Y.; Wang, D.; Zhong, X.; Chen, W.; Chen, X.; Chen, Y.; et al. Effects of pharmacological treatment on metabolomic alterations in animal models of depression. Transl. Psychiatry 2022, 12, 175. [Google Scholar] [CrossRef]
- Zheng, S.; Yu, M.; Lu, X.; Huo, T.; Ge, L.; Yang, J.; Wu, C.; Li, F. Urinary metabonomic study on biochemical changes in chronic unpredictable mild stress model of depression. Clin. Chim. Acta 2010, 411, 204–209. [Google Scholar] [CrossRef]
- Tian, P.; O’Riordan, K.J.; Lee, Y.-K.; Wang, G.; Zhao, J.; Zhang, H.; Cryan, J.F.; Chen, W. Towards a psychobiotic therapy for depression: Bifidobacterium breve CCFM1025 reverses chronic stress-induced depressive symptoms and gut microbial abnormalities in mice. Neurobiol. Stress 2020, 12, 100216. [Google Scholar] [CrossRef]
- Lefèvre, A.; Mavel, S.; Nadal-Desbarats, L.; Galineau, L.; Attucci, S.; Dufour, D.; Sokol, H.; Emond, P. Validation of a global quantitative analysis methodology of tryptophan metabolites in mice using LC-MS. Talanta 2019, 195, 593–598. [Google Scholar] [CrossRef]
- Zhu, G.; Ma, F.; Wang, G.; Wang, Y.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacteria attenuate the development of metabolic disorders, with inter- and intra-species differences. Food Funct. 2018, 9, 3509–3522. [Google Scholar] [CrossRef]
- Badawy, A. Novel nutritional treatment for manic and psychotic disorders: A review of tryptophan and tyrosine depletion studies and the potential of protein-based formulations using glycomacropeptide. Psychopharmacology 2013, 228, 347–358. [Google Scholar] [CrossRef]
- Fukuwatari, T. Possibility of amino acid treatment to prevent the psychiatric disorders via modulation of the production of tryptophan metabolite kynurenic acid. Nutrients 2020, 12, 1403. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawrońska-Grzywacz, M.; Herbet, M. HPA Axis in the pathomechanism of depression and schizophrenia: New therapeutic strategies based on its participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef]
- Thomson, F.; Craighead, M. Innovative approaches for the treatment of depression: Targeting the HPA axis. Neurochem. Res. 2008, 33, 691–707. [Google Scholar] [CrossRef]
- Castrén, E.; Monteggia, L.M. Brain-derived neurotrophic factor signaling in depression and antidepressant action. Biol. Psychiatry 2021, 90, 128–136. [Google Scholar] [CrossRef]
- Hing, B.; Sathyaputri, L.; Potash, J.B. A comprehensive review of genetic and epigenetic mechanisms that regulate BDNF expression and function with relevance to major depressive disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2018, 177, 143–167. [Google Scholar] [CrossRef]
- Popova, N.K.; Ilchibaeva, T.V.; Naumenko, V.S. Neurotrophic factors (BDNF and GDNF) and the serotonergic system of the brain. Biochemistry 2017, 82, 308–317. [Google Scholar] [CrossRef]
- Weilburg, J.B. An overview of SSRI and SNRI therapies for depression. Manag. Care 2004, 13, 25–33. [Google Scholar]
- Ye, L.; Bae, M.; Cassilly, C.D.; Jabba, S.V.; Thorpe, D.W.; Martin, A.M.; Lu, H.-Y.; Wang, J.; Thompson, J.D.; Lickwar, C.R.; et al. Enteroendocrine cells sense bacterial tryptophan catabolites to activate enteric and vagal neuronal pathways. Cell Host Microbe 2021, 29, 179–196.e179. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef] [Green Version]
- Saturio, S.; Nogacka, A.M.; Alvarado-Jasso, G.M.; Salazar, N.; Reyes-Gavilán, C.G.D.L.; Gueimonde, M.; Arboleya, S. Role of bifidobacteria on infant health. Microorganisms 2021, 9, 2415. [Google Scholar] [CrossRef]
- Tian, P.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium with the role of 5-hydroxytryptophan synthesis regulation alleviates the symptom of depression and related microbiota dysbiosis. J. Nutr. Biochem. 2019, 66, 43–51. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhao, J.; Li, C.; Zhang, M.; Wei, L.; Zhang, X.; Kurskaya, O.; Bi, H.; Gao, T. Effect of combined chronic predictable and unpredictable stress on depression-like symptoms in mice. Ann. Transl. Med. 2020, 8, 942. [Google Scholar] [CrossRef]
- Xu, J.; Tang, M.; Wu, X.; Kong, X.; Liu, Y.; Xu, X.-X. Lactobacillus rhamnosus zz-1 exerts preventive effects on chronic unpredictable mild stress-induced depression in mice via regulating the intestinal microenvironment. Food Funct. 2022, 13, 4331–4343. [Google Scholar] [CrossRef]
- Lu, S.; Mikkelsen, D.; Yao, H.; Williams, B.A.; Flanagan, B.M.; Gidley, M.J. Wheat cell walls and constituent polysaccharides induce similar microbiota profiles upon in vitro fermentation despite different short chain fatty acid end-product levels. Food Funct. 2021, 12, 1135–1146. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Tian, P.; Wang, Z.; Pan, R.; Shang, K.; Wang, G.; Zhao, J.; Chen, W. Indole Acetic Acid Exerts Anti-Depressive Effects on an Animal Model of Chronic Mild Stress. Nutrients 2022, 14, 5019. https://doi.org/10.3390/nu14235019
Chen Y, Tian P, Wang Z, Pan R, Shang K, Wang G, Zhao J, Chen W. Indole Acetic Acid Exerts Anti-Depressive Effects on an Animal Model of Chronic Mild Stress. Nutrients. 2022; 14(23):5019. https://doi.org/10.3390/nu14235019
Chicago/Turabian StyleChen, Ying, Peijun Tian, Zheng Wang, Ruili Pan, Kexin Shang, Gang Wang, Jianxin Zhao, and Wei Chen. 2022. "Indole Acetic Acid Exerts Anti-Depressive Effects on an Animal Model of Chronic Mild Stress" Nutrients 14, no. 23: 5019. https://doi.org/10.3390/nu14235019