

Modulation of Human Motor Cortical Excitability and Plasticity by Opuntia Ficus Indica Fruit Consumption: Evidence from a Preliminary Study through Non-Invasive Brain Stimulation
 ,
,  , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
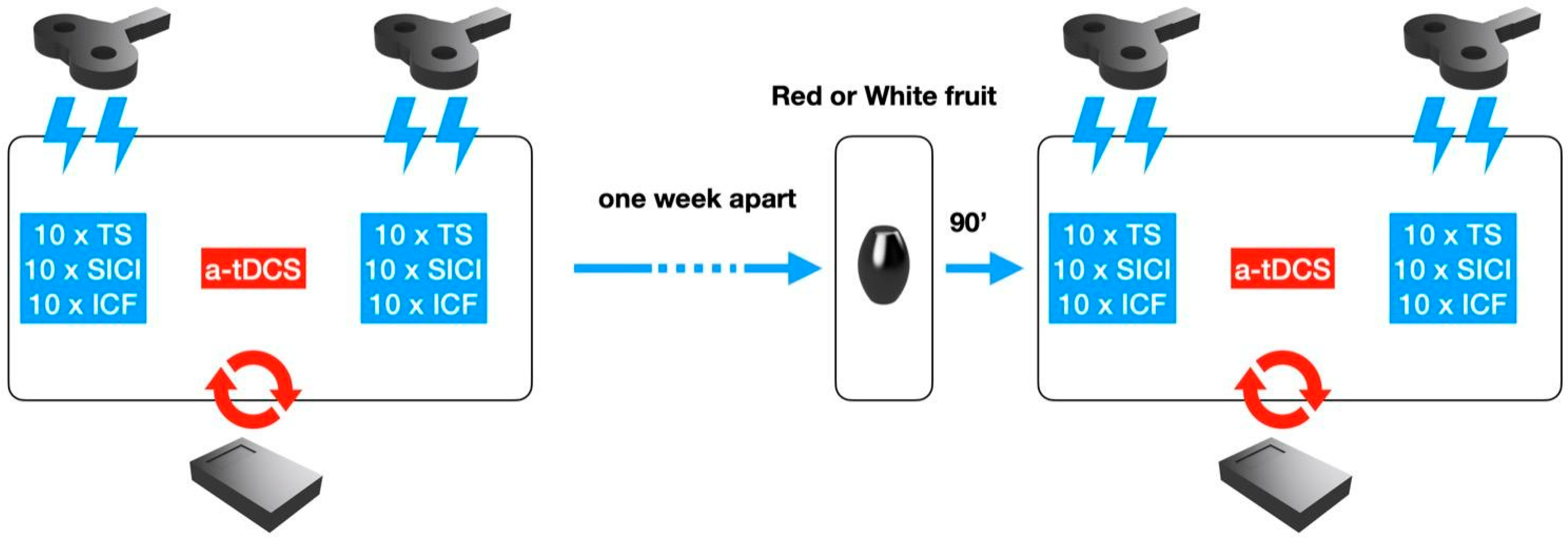
2.2. Experimental Procedure
2.2.1. ppTMS
2.2.2. a-TDCS
2.2.3. Rs-fMRI Acquisition
2.3. Statistical Analysis
3. Results
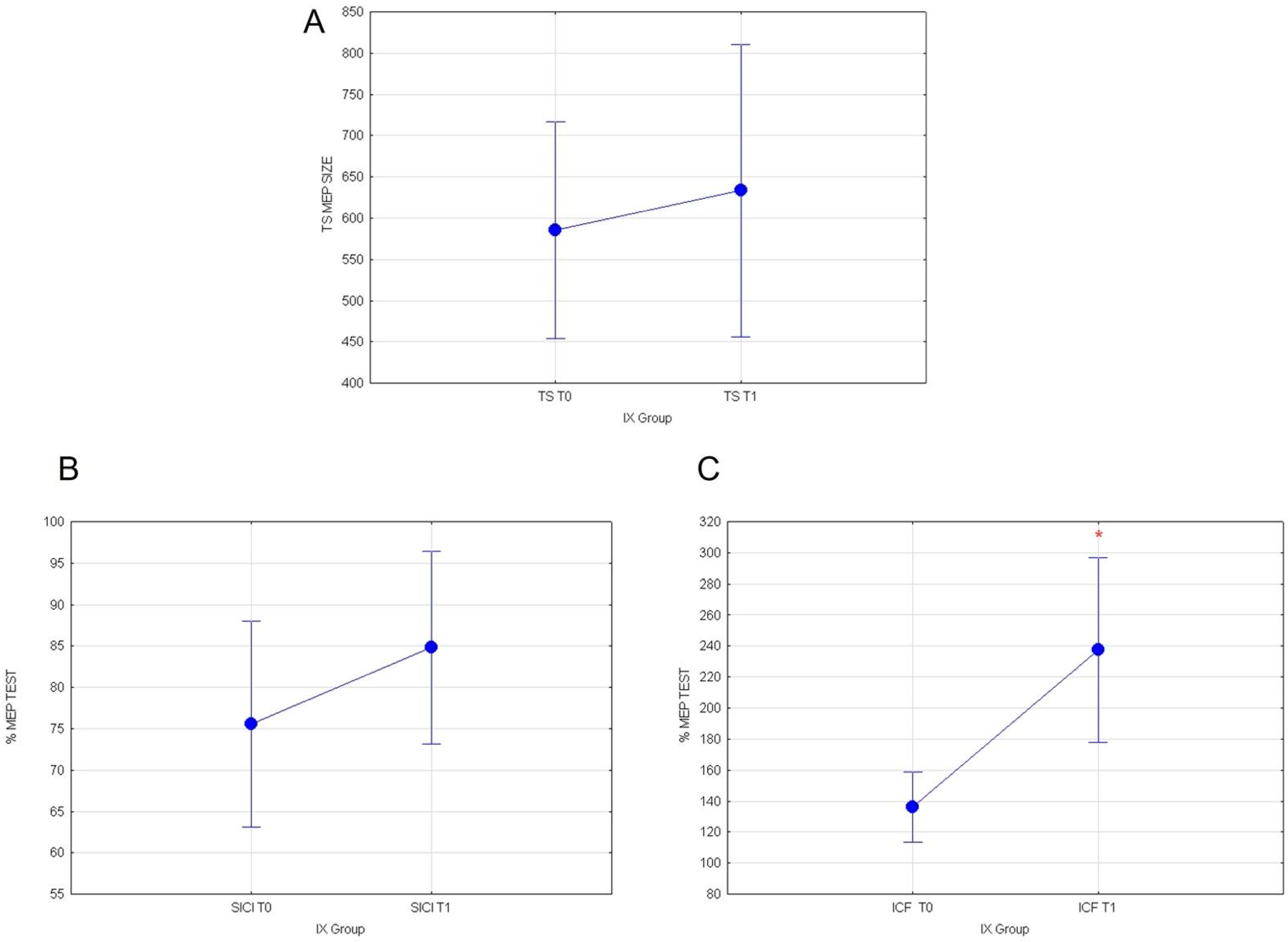
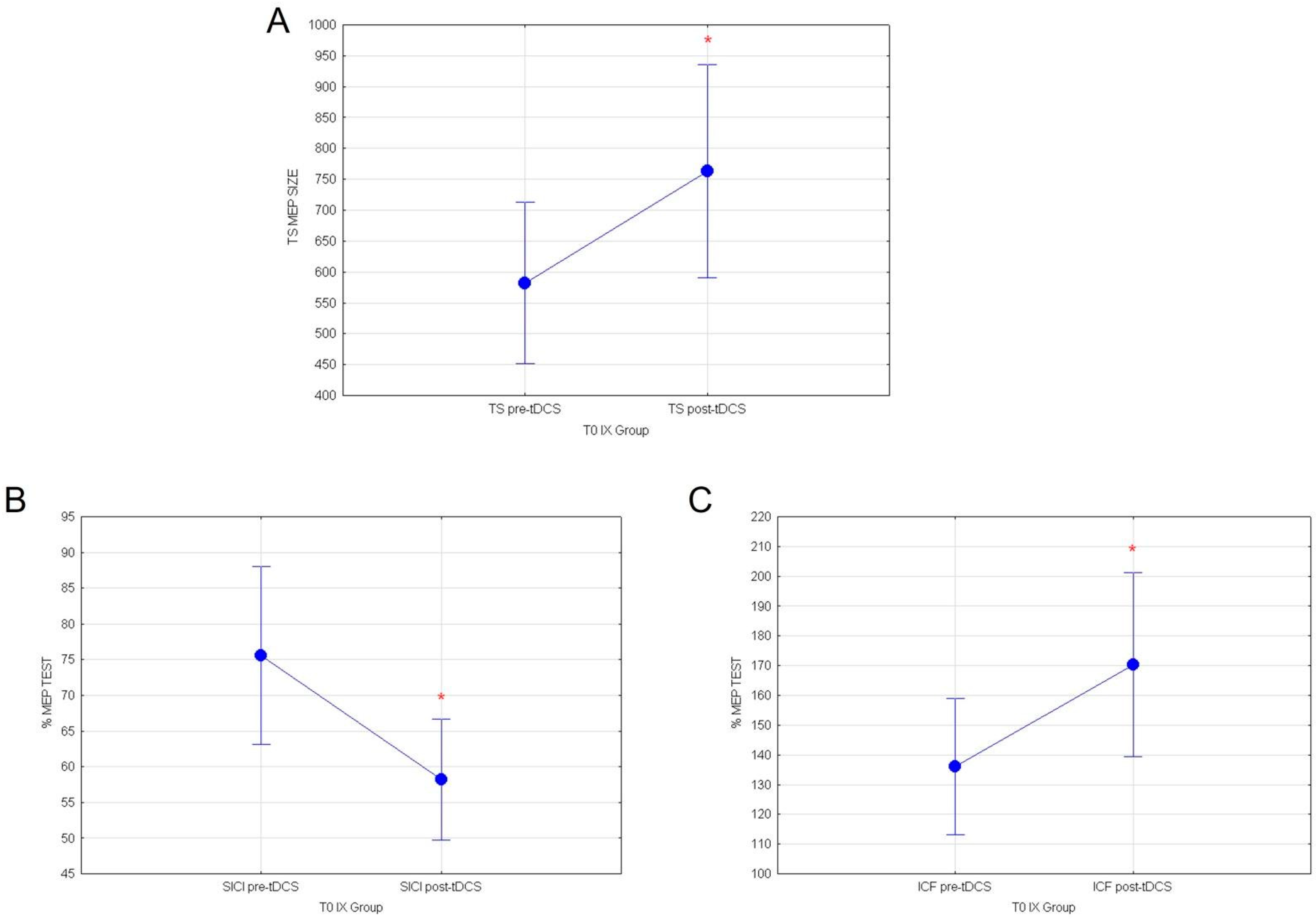
3.1. Effects of Cactus Pear Fruit on Corticospinal Excitability (TS), Intracortical Facilitation (ICF), Intracortical Inhibition (SICI), and Cortical Plasticity (a-tDCS) in IX Group
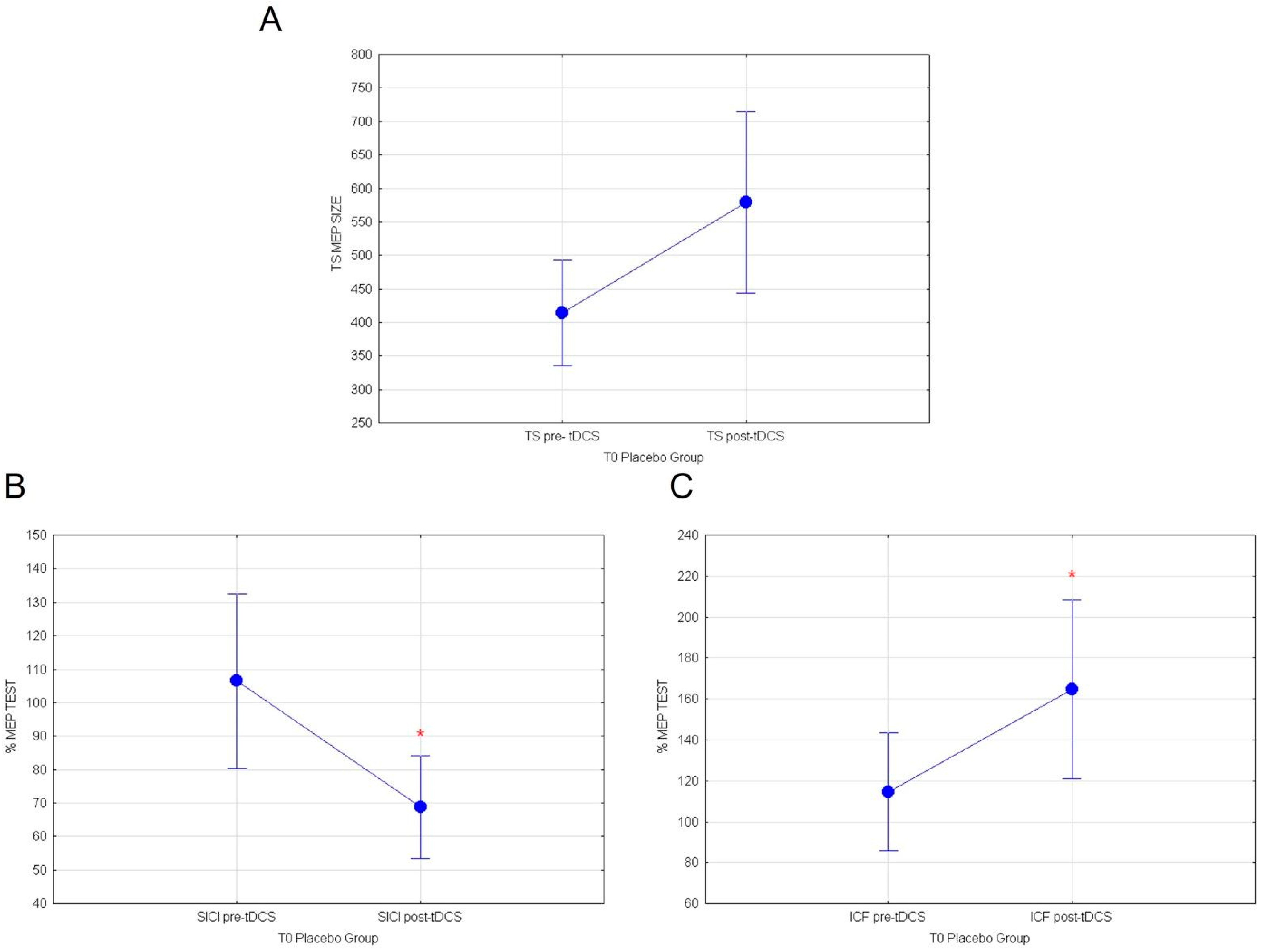
3.2. Effects of Cactus Pear Fruit on Corticospinal Excitability (TS), Intracortical Facilitation (ICF), Intracortical Inhibition (SICI), and Cortical Plasticity (a-tDCS) in the Placebo Group
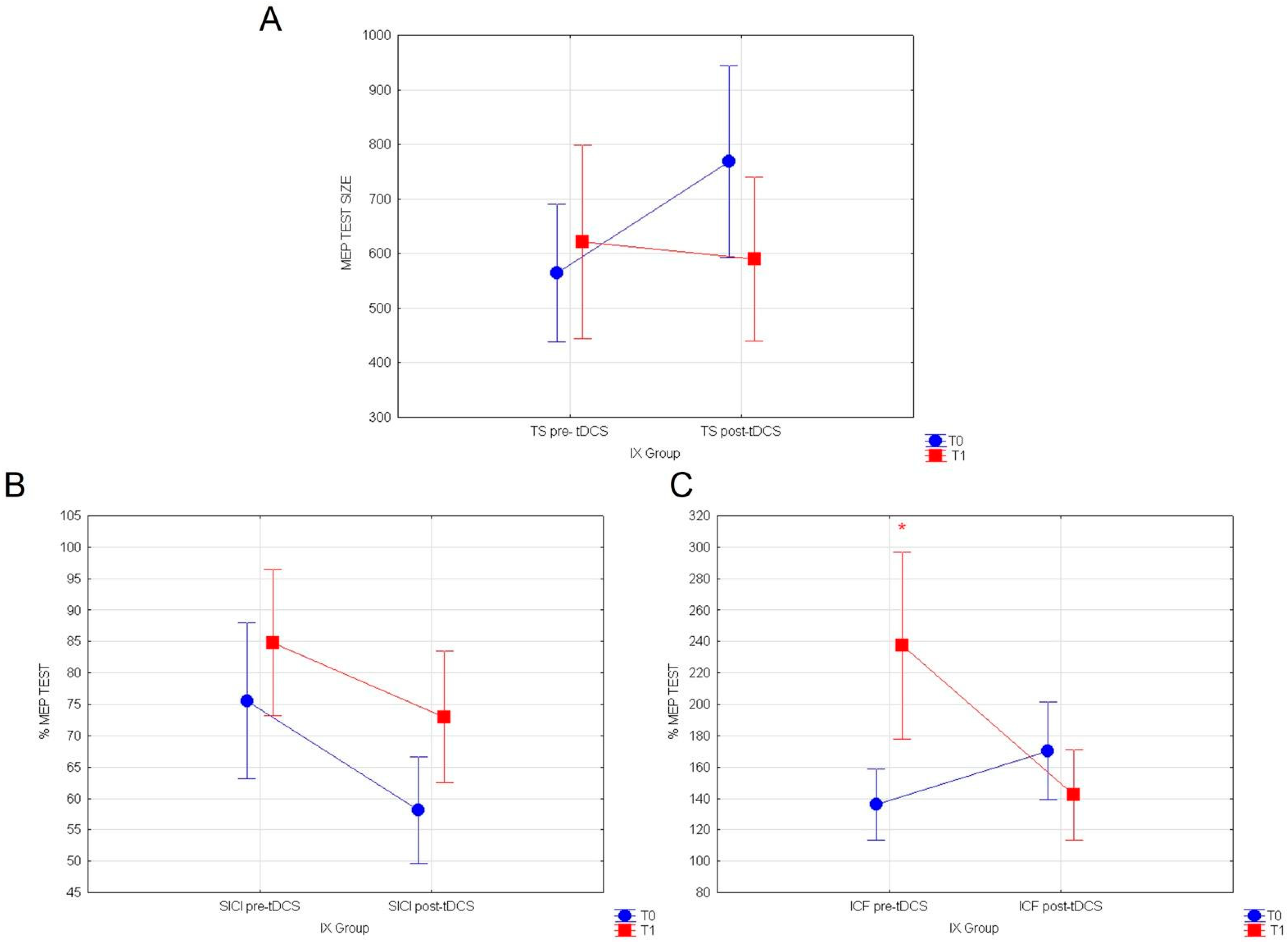
3.3. Effects of a-tDCS on Corticospinal Excitability (TS), Intracortical Facilitation (ICF), and Inhibition (SICI) before Assuming Cactus Pear Fruit in IX and Placebo Groups
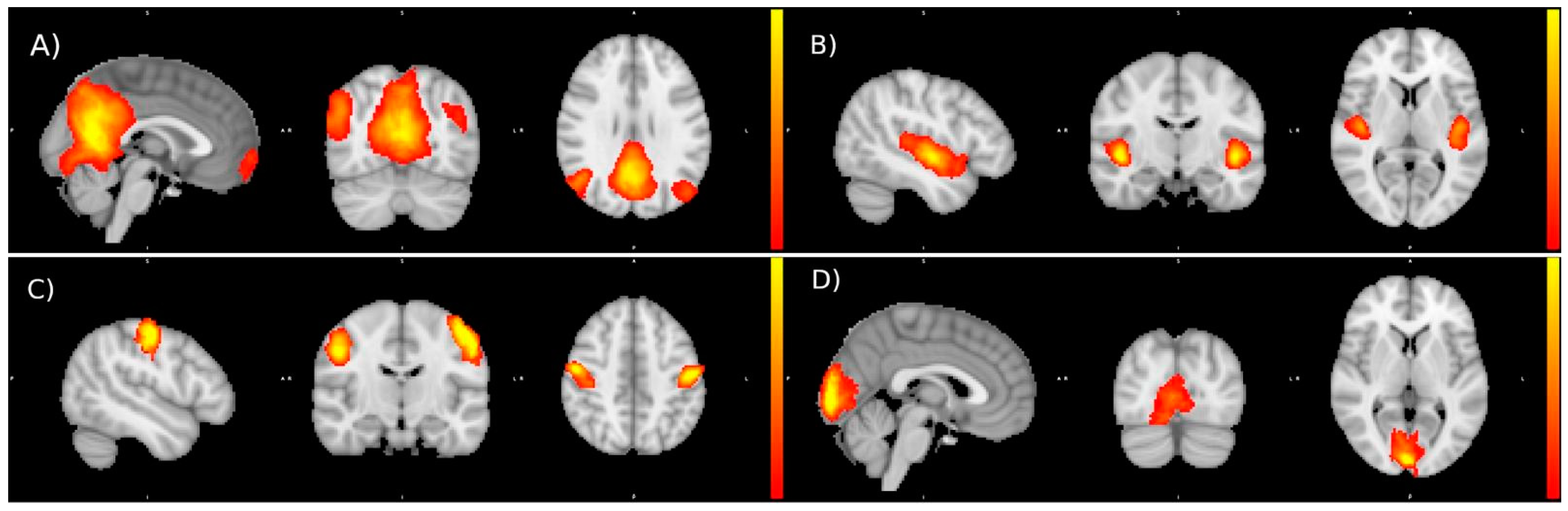
3.4. Preliminary Functional Connectivity Investigations after Cactus Pear Fruit Consumption
4. Discussion
5. Conclusions
Possible Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giglia, G.; Gambino, G.; Cuffaro, L.; Aleo, F.; Sardo, P.; Ferraro, G.; Blandino, V.; Brighina, F.; Gangitano, M.; Piccoli, T. Modulating Long Term Memory at Late-Encoding Phase: An rTMS Study. Brain Topogr. 2021, 34, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Castrillon, G.; Sollmann, N.; Kurcyus, K.; Razi, A.; Krieg, S.M.; Riedl, V. The Physiological Effects of Noninvasive Brain Stimulation Fundamentally Differ across the Human Cortex. Sci. Adv. 2020, 6, eaay2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allegra, M. Antioxidant and Anti-Inflammatory Properties of Plants Extract. Antioxidants 2019, 8, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoi, M.; Shamoto-Nagai, M.; Maruyama, W. Neuroprotection of Multifunctional Phytochemicals as Novel Therapeutic Strategy for Neurodegenerative Disorders: Antiapoptotic and Antiamyloidogenic Activities by Modulation of Cellular Signal Pathways. Future Neurol. 2019, 14, FNL9. [Google Scholar] [CrossRef] [Green Version]
- Figueira, I.; Garcia, G.; Pimpão, R.C.; Terrasso, A.P.; Costa, I.; Almeida, A.F.; Tavares, L.; Pais, T.F.; Pinto, P.; Ventura, M.R.; et al. Polyphenols Journey through Blood-Brain Barrier towards Neuronal Protection. Sci. Rep. 2017, 7, 11456. [Google Scholar] [CrossRef] [Green Version]
- Ovais, M.; Zia, N.; Ahmad, I.; Khalil, A.T.; Raza, A.; Ayaz, M.; Sadiq, A.; Ullah, F.; Shinwari, Z.K. Phyto-Therapeutic and Nanomedicinal Approaches to Cure Alzheimer’s Disease: Present Status and Future Opportunities. Front. Aging Neurosci. 2018, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.-M.; Liu, H.-Y.; Xie, L.; Liu, X.-D. Effect of Baicalin and Berberine on Transport of Nimodipine on Primary-Cultured, Rat Brain Microvascular Endothelial Cells. Acta Pharmacol. Sin. 2007, 28, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Berti, V.; Walters, M.; Sterling, J.; Quinn, C.G.; Logue, M.; Andrews, R.; Matthews, D.C.; Osorio, R.S.; Pupi, A.; Vallabhajosula, S.; et al. Mediterranean Diet and 3-Year Alzheimer Brain Biomarker Changes in Middle-Aged Adults. Neurology 2018, 90, e1789–e1798. [Google Scholar] [CrossRef]
- Spencer, J.P.E. Flavonoids: Modulators of Brain Function? Br. J. Nutr. 2008, 99 (Suppl. 1), ES60–ES77. [Google Scholar] [CrossRef] [Green Version]
- Rendeiro, C.; Guerreiro, J.D.T.; Williams, C.M.; Spencer, J.P.E. Flavonoids as Modulators of Memory and Learning: Molecular Interactions Resulting in Behavioural Effects. Proc. Nutr. Soc. 2012, 71, 246–262. [Google Scholar] [CrossRef]
- Meng, B.; Shen, L.-L.; Shi, X.-T.; Gong, Y.-S.; Fan, X.-F.; Li, J.; Cao, H. Effects of Curcumin on TTX-R Sodium Currents of Dorsal Root Ganglion Neurons in Type 2 Diabetic Rats with Diabetic Neuropathic Pain. Neurosci. Lett. 2015, 605, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Watanabe, C.M.H.; Schultz, P.G.; Rimbach, G.; Krucker, T. Age-Related Effects of Ginkgo Biloba Extract on Synaptic Plasticity and Excitability. Neurobiol. Aging 2004, 25, 955–962. [Google Scholar] [CrossRef]
- Kalmar, J.M.; Cafarelli, E. Central Fatigue and Transcranial Magnetic Stimulation: Effect of Caffeine and the Confound of Peripheral Transmission Failure. J. Neurosci. Methods 2004, 138, 15–26. [Google Scholar] [CrossRef]
- Specterman, M.; Bhuiya, A.; Kuppuswamy, A.; Strutton, P.H.; Catley, M.; Davey, N.J. The Effect of an Energy Drink Containing Glucose and Caffeine on Human Corticospinal Excitability. Physiol. Behav. 2005, 83, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Orth, M.; Amann, B.; Ratnaraj, N.; Patsalos, P.N.; Rothwell, J.C. Caffeine Has No Effect on Measures of Cortical Excitability. Clin. Neurophysiol. 2005, 116, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, V.; de Mendonça, A.; Minez, A.; Dias, A.R.; de Carvalho, M. Does Caffeine Modify Corticomotor Excitability? Neurophysiol. Clin. 2006, 36, 219–226. [Google Scholar] [CrossRef]
- de Carvalho, M.; de Carvalho, M.; Marcelino, E.; de Mendonça, A. Electrophysiological Studies in Healthy Subjects Involving Caffeine. J. Alzheimer’s Disease 2010, 20, S63–S69. [Google Scholar] [CrossRef] [Green Version]
- Nardone, R.; Buffone, E.; Covi, M.; Lochner, P.G.; Tezzon, F. Changes in Motor Cortical Excitability in Humans Following Orally Administered Theophylline. Neurosci. Lett. 2004, 355, 65–68. [Google Scholar] [CrossRef]
- Tesoriere, L.; Butera, D.; Pintaudi, A.M.; Allegra, M.; Livrea, M.A. Supplementation with Cactus Pear (Opuntia Ficus-Indica) Fruit Decreases Oxidative Stress in Healthy Humans: A Comparative Study with Vitamin C. Am. J. Clin. Nutr. 2004, 80, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Attanzio, A.; Tesoriere, L.; Vasto, S.; Pintaudi, A.M.; Livrea, M.A.; Allegra, M. Short-Term Cactus Pear [Opuntia Ficus-Indica (L.) Mill] Fruit Supplementation Ameliorates the Inflammatory Profile and Is Associated with Improved Antioxidant Status among Healthy Humans. Food Nutr. Res. 2018, 62. [Google Scholar] [CrossRef]
- Piattelli, M.; Minale, L.; Nicolaus, R.A. Pigments of centrospermae—V. Phytochemistry 1965, 4, 817–823. [Google Scholar] [CrossRef]
- Allegra, M.; Tutone, M.; Tesoriere, L.; Almerico, A.M.; Culletta, G.; Livrea, M.A.; Attanzio, A. Indicaxanthin, a Multi-Target Natural Compound from Opuntia Ficus-Indica Fruit: From Its Poly-Pharmacological Effects to Biochemical Mechanisms and Molecular Modelling Studies. Eur. J. Med. Chem. 2019, 179, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Terzo, S.; Attanzio, A.; Calvi, P.; Mulè, F.; Tesoriere, L.; Allegra, M.; Amato, A. Indicaxanthin from Opuntia Ficus-Indica Fruit Ameliorates Glucose Dysmetabolism and Counteracts Insulin Resistance in High-Fat-Diet-Fed Mice. Antioxidants 2021, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; Carletti, F.; Gambino, G.; Tutone, M.; Attanzio, A.; Tesoriere, L.; Ferraro, G.; Sardo, P.; Almerico, A.M.; Livrea, M.A. Indicaxanthin from Opuntia Ficus-Indica Crosses the Blood-Brain Barrier and Modulates Neuronal Bioelectric Activity in Rat Hippocampus at Dietary-Consistent Amounts. J. Agric. Food Chem. 2015, 63, 7353–7360. [Google Scholar] [CrossRef]
- Tutone, M.; Virzì, A.; Almerico, A.M. Reverse Screening on Indicaxanthin from Opuntia Ficus-Indica as Natural Chemoactive and Chemopreventive Agent. J. Theor. Biol. 2018, 455, 147–160. [Google Scholar] [CrossRef]
- Tesoriere, L.; Allegra, M.; Butera, D.; Livrea, M.A. Absorption, Excretion, and Distribution of Dietary Antioxidant Betalains in LDLs: Potential Health Effects of Betalains in Humans. Am. J. Clin. Nutr. 2004, 80, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Gambino, G.; Allegra, M.; Sardo, P.; Attanzio, A.; Tesoriere, L.; Livrea, M.A.; Ferraro, G.; Carletti, F. Brain Distribution and Modulation of Neuronal Excitability by Indicaxanthin from Administered at Nutritionally-Relevant Amounts. Front. Aging Neurosci. 2018, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, U.; Reis, J.; Schwenkreis, P.; Rosanova, M.; Strafella, A.; Badawy, R.; Müller-Dahlhaus, F. TMS and Drugs Revisited 2014. Clin. Neurophysiol. 2015, 126, 1847–1868. [Google Scholar] [CrossRef]
- Palmeri, R.; Parafati, L.; Arena, E.; Grassenio, E.; Restuccia, C.; Fallico, B. Antioxidant and Antimicrobial Properties of Semi-Processed Frozen Prickly Pear Juice as Affected by Cultivar and Harvest Time. Foods 2020, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Polanía, R.; Nitsche, M.A.; Ruff, C.C. Studying and Modifying Brain Function with Non-Invasive Brain Stimulation. Nat. Neurosci. 2018, 21, 174–187. [Google Scholar] [CrossRef]
- Cosentino, G.; Fierro, B.; Vigneri, S.; Talamanca, S.; Paladino, P.; Baschi, R.; Indovino, S.; Maccora, S.; Valentino, F.; Fileccia, E.; et al. Cyclical Changes of Cortical Excitability and Metaplasticity in Migraine: Evidence from a Repetitive Transcranial Magnetic Stimulation Study. Pain 2014, 155, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Giglia, G.; Brighina, F.; Indovino, S.; Fierro, B. Ipsilesional and Contralesional Regions Participate in the Improvement of Poststroke Aphasia: A Transcranial Direct Current Stimulation Study. Neurocase 2015, 21, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- American Psychiatric Association. Task Force on Nomenclature and Statistics Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Washington, DC, USA, 1980; ISBN 9780890425558. [Google Scholar]
- Rossi, S.; Antal, A.; Bestmann, S.; Bikson, M.; Brewer, C.; Brockmöller, J.; Carpenter, L.L.; Cincotta, M.; Chen, R.; Daskalakis, J.D.; et al. Safety and Recommendations for TMS Use in Healthy Subjects and Patient Populations, with Updates on Training, Ethical and Regulatory Issues: Expert Guidelines. Clin. Neurophysiol. 2021, 132, 269–306. [Google Scholar] [CrossRef]
- Giglia, G.; Pia, L.; Folegatti, A.; Puma, A.; Fierro, B.; Cosentino, G.; Berti, A.; Brighina, F. Far Space Remapping by Tool Use: A rTMS Study Over the Right Posterior Parietal Cortex. Brain Stimul. 2015, 8, 795–800. [Google Scholar] [CrossRef]
- Costa, V.; Giglia, G.; Talamanca, S.; Finocchiaro, C.; Cosentino, G.; Fierro, B.; Brighina, F. Transcranial Direct Current Stimulation over the Right Inferior Parietal Cortex Reduces Transposition Errors in a Syllabic Reordering Task. Symmetry 2021, 13, 2077. [Google Scholar] [CrossRef]
- Piccoli, T.; Maniaci, G.; Collura, G.; Gagliardo, C.; Brancato, A.; La Tona, G.; Gangitano, M.; La Cascia, C.; Picone, F.; Marrale, M.; et al. Increased Functional Connectivity in Gambling Disorder Correlates with Behavioural and Emotional Dysregulation: Evidence of a Role for the Cerebellum. Behav. Brain Res. 2020, 390, 112668. [Google Scholar] [CrossRef]
- Biabani, M.; Aminitehrani, M.; Zoghi, M.; Farrell, M.; Egan, G.; Jaberzadeh, S. The Effects of Transcranial Direct Current Stimulation on Short-Interval Intracortical Inhibition and Intracortical Facilitation: A Systematic Review and Meta-Analysis. Rev. Neurosci. 2018, 29, 99–114. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Sustained Excitability Elevations Induced by Transcranial DC Motor Cortex Stimulation in Humans. Neurology 2001, 57, 1899–1901. [Google Scholar] [CrossRef]
- Trepel, C.; Racine, R.J. Long-Term Potentiation in the Neocortex of the Adult, Freely Moving Rat. Cereb. Cortex 1998, 8, 719–729. [Google Scholar] [CrossRef]
- Froc, D.J.; Chapman, C.A.; Trepel, C.; Racine, R.J. Long-Term Depression and Depotentiation in the Sensorimotor Cortex of the Freely Moving Rat. J. Neurosci. 2000, 20, 438–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, M.A.; Antal, A.; Liebetanz, D.; Lang, N.; Tergau, F.; Paulus, W. Erzeugung Und Modulation von Neuroplastizität Durch Transkranielle Gleichstromstimulation Beim Menschen. Klin. Neurophysiol. 2004, 35, 55–60. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Liebetanz, D.; Schlitterlau, A.; Henschke, U.; Fricke, K.; Frommann, K.; Lang, N.; Henning, S.; Paulus, W.; Tergau, F. GABAergic Modulation of DC Stimulation-Induced Motor Cortex Excitability Shifts in Humans. Eur. J. Neurosci. 2004, 19, 2720–2726. [Google Scholar] [CrossRef] [PubMed]
- Brighina, F.; Cosentino, G.; Vigneri, S.; Talamanca, S.; Palermo, A.; Giglia, G.; Fierro, B. Abnormal Facilitatory Mechanisms in Motor Cortex of Migraine with Aura. Eur. J. Pain 2011, 15, 928–935. [Google Scholar] [CrossRef]
- Baxter, P.S.; Hardingham, G.E. Adaptive Regulation of the Brain’s Antioxidant Defences by Neurons and Astrocytes. Free Radic. Biol. Med. 2016, 100, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorio, M.; Emadi Andani, M.; Marotta, A.; Classen, J.; Tinazzi, M. Placebo-Induced Changes in Excitatory and Inhibitory Corticospinal Circuits during Motor Performance. J. Neurosci. 2014, 34, 3993–4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüscher, C.; Malenka, R.C. NMDA Receptor-Dependent Long-Term Potentiation and Long-Term Depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 2012, 4, a005710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mineo, L.; Concerto, C.; Patel, D.; Mayorga, T.; Paula, M.; Chusid, E.; Aguglia, E.; Battaglia, F. Valeriana Officinalis Root Extract Modulates Cortical Excitatory Circuits in Humans. Neuropsychobiology 2017, 75, 46–51. [Google Scholar] [CrossRef]
- Concerto, C.; Boo, H.; Hu, C.; Sandilya, P.; Krish, A.; Chusid, E.; Coira, D.; Aguglia, E.; Battaglia, F. Hypericum Perforatum Extract Modulates Cortical Plasticity in Humans. Psychopharmacology 2018, 235, 145–153. [Google Scholar] [CrossRef]
- Schulte-Löbbert, S.; Holoubek, G.; Müller, W.E.; Schubert-Zsilavecz, M.; Wurglics, M. Comparison of the Synaptosomal Uptake Inhibition of Serotonin by St John’s Wort Products. J. Pharm. Pharmacol. 2010, 56, 813–818. [Google Scholar] [CrossRef]
- Gambino, G.; Rizzo, V.; Giglia, G.; Ferraro, G.; Sardo, P. Microtubule Dynamics and Neuronal Excitability: Advances on Cytoskeletal Components Implicated in Epileptic Phenomena. Cell. Mol. Neurobiol. 2022, 42, 533–543. [Google Scholar] [CrossRef]
- Gambino, G.; Rizzo, V.; Giglia, G.; Ferraro, G.; Sardo, P. Cannabinoids, TRPV and Nitric Oxide: The Three Ring Circus of Neuronal Excitability. Brain Struct. Funct. 2020, 225, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jaudon, F.; Thalhammer, A.; Cingolani, L.A. Integrin Adhesion in Brain Assembly: From Molecular Structure to Neuropsychiatric Disorders. Eur. J. Neurosci. 2021, 53, 3831–3850. [Google Scholar] [CrossRef] [PubMed]
- Kavalali, E.T.; Monteggia, L.M. Targeting Homeostatic Synaptic Plasticity for Treatment of Mood Disorders. Neuron 2020, 106, 715–726. [Google Scholar] [CrossRef]
- Kennedy, D.O. Phytochemicals for Improving Aspects of Cognitive Function and Psychological State Potentially Relevant to Sports Performance. Sports Med. 2019, 49, 39–58. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-tdcs vs. Post-tdcs | |
|---|---|
| IX Group (t0) | |
| TS | p = 0.0487 |
| SICI | p = 0.023 |
| ICF | p = 0.047 |
| Placebo Group (t0) | |
| TS | p = 0.053 |
| SICI | p = 0.012 |
| ICF | p = 0.0259 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambino, G.; Brighina, F.; Allegra, M.; Marrale, M.; Collura, G.; Gagliardo, C.; Attanzio, A.; Tesoriere, L.; Di Majo, D.; Ferraro, G.; et al. Modulation of Human Motor Cortical Excitability and Plasticity by Opuntia Ficus Indica Fruit Consumption: Evidence from a Preliminary Study through Non-Invasive Brain Stimulation. Nutrients 2022, 14, 4915. https://doi.org/10.3390/nu14224915
Gambino G, Brighina F, Allegra M, Marrale M, Collura G, Gagliardo C, Attanzio A, Tesoriere L, Di Majo D, Ferraro G, et al. Modulation of Human Motor Cortical Excitability and Plasticity by Opuntia Ficus Indica Fruit Consumption: Evidence from a Preliminary Study through Non-Invasive Brain Stimulation. Nutrients. 2022; 14(22):4915. https://doi.org/10.3390/nu14224915
Chicago/Turabian StyleGambino, Giuditta, Filippo Brighina, Mario Allegra, Maurizio Marrale, Giorgio Collura, Cesare Gagliardo, Alessandro Attanzio, Luisa Tesoriere, Danila Di Majo, Giuseppe Ferraro, and et al. 2022. "Modulation of Human Motor Cortical Excitability and Plasticity by Opuntia Ficus Indica Fruit Consumption: Evidence from a Preliminary Study through Non-Invasive Brain Stimulation" Nutrients 14, no. 22: 4915. https://doi.org/10.3390/nu14224915