
Indolepropionic Acid, a Gut Bacteria-Produced Tryptophan Metabolite and the Risk of Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease
 , and
, and
Abstract
:1. Introduction
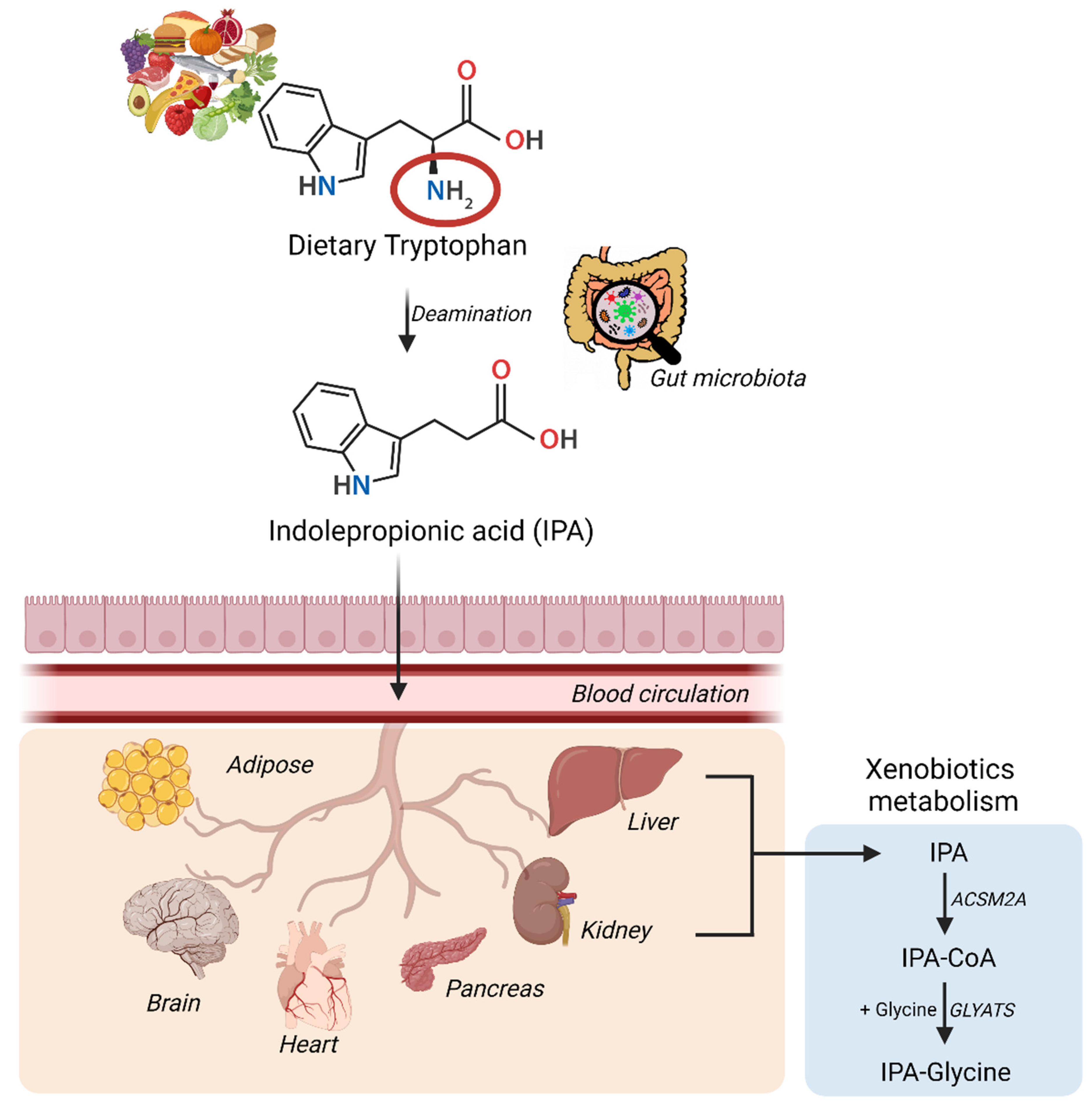
2. Diet, Gut Microbiota, and Indolepropionic Acid
2.1. The Role of Gut Microbiota in the Formation of IPA
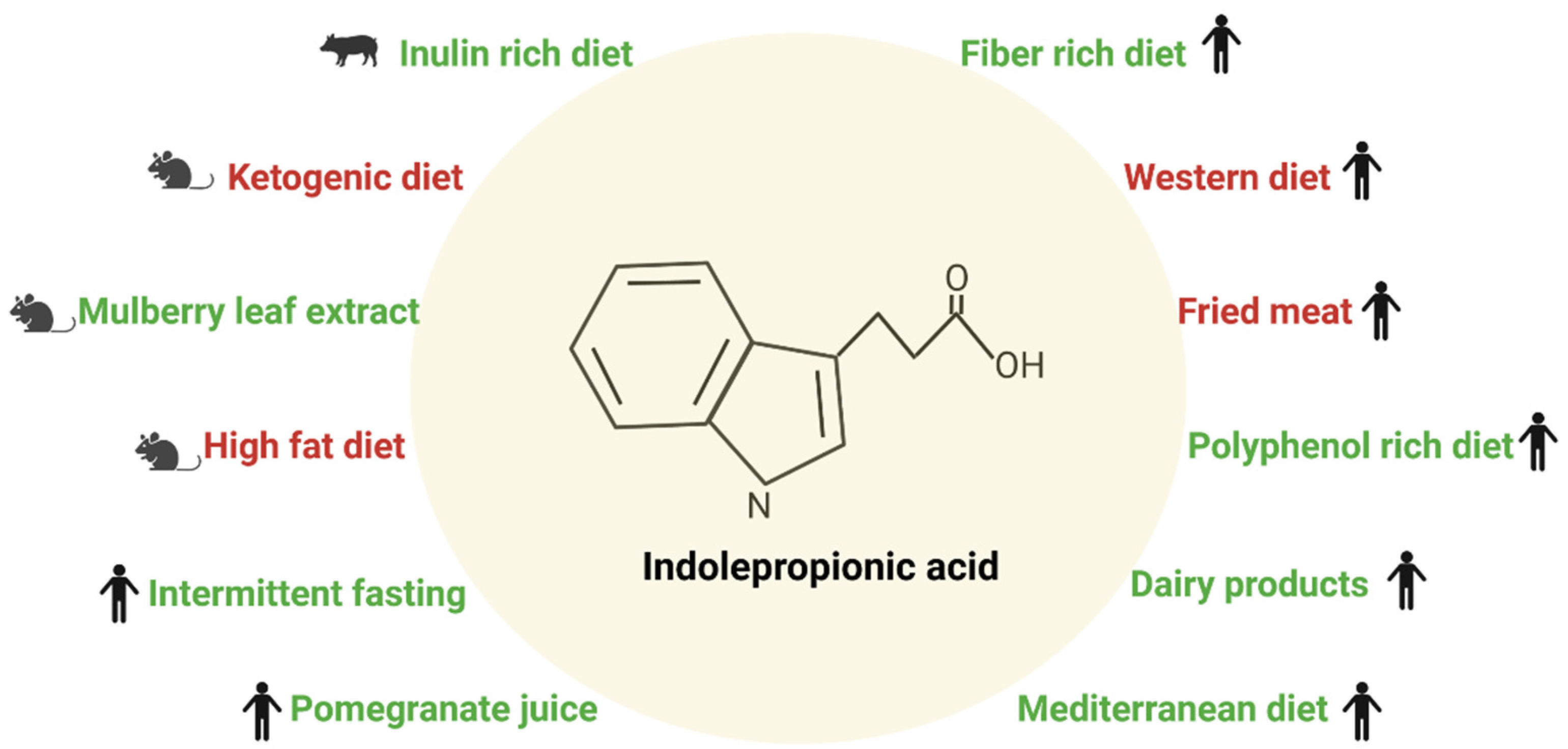
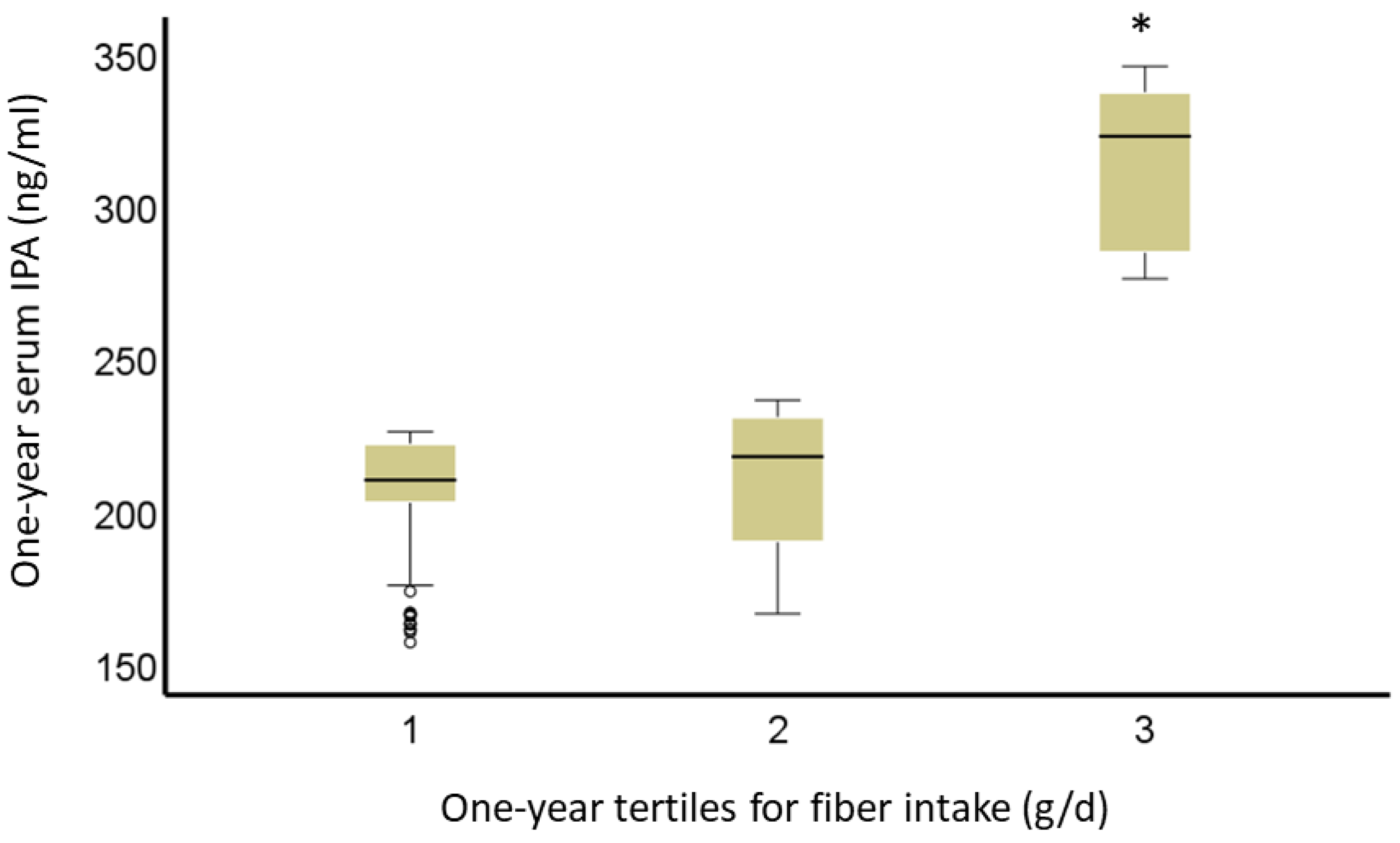
2.2. Diet and Indolepropionic Acid
3. Indolepropionic Acid and Type 2 Diabetes
3.1. Effect of Indolepropionic Acid on Glucose Metabolism and Insulin Secretion
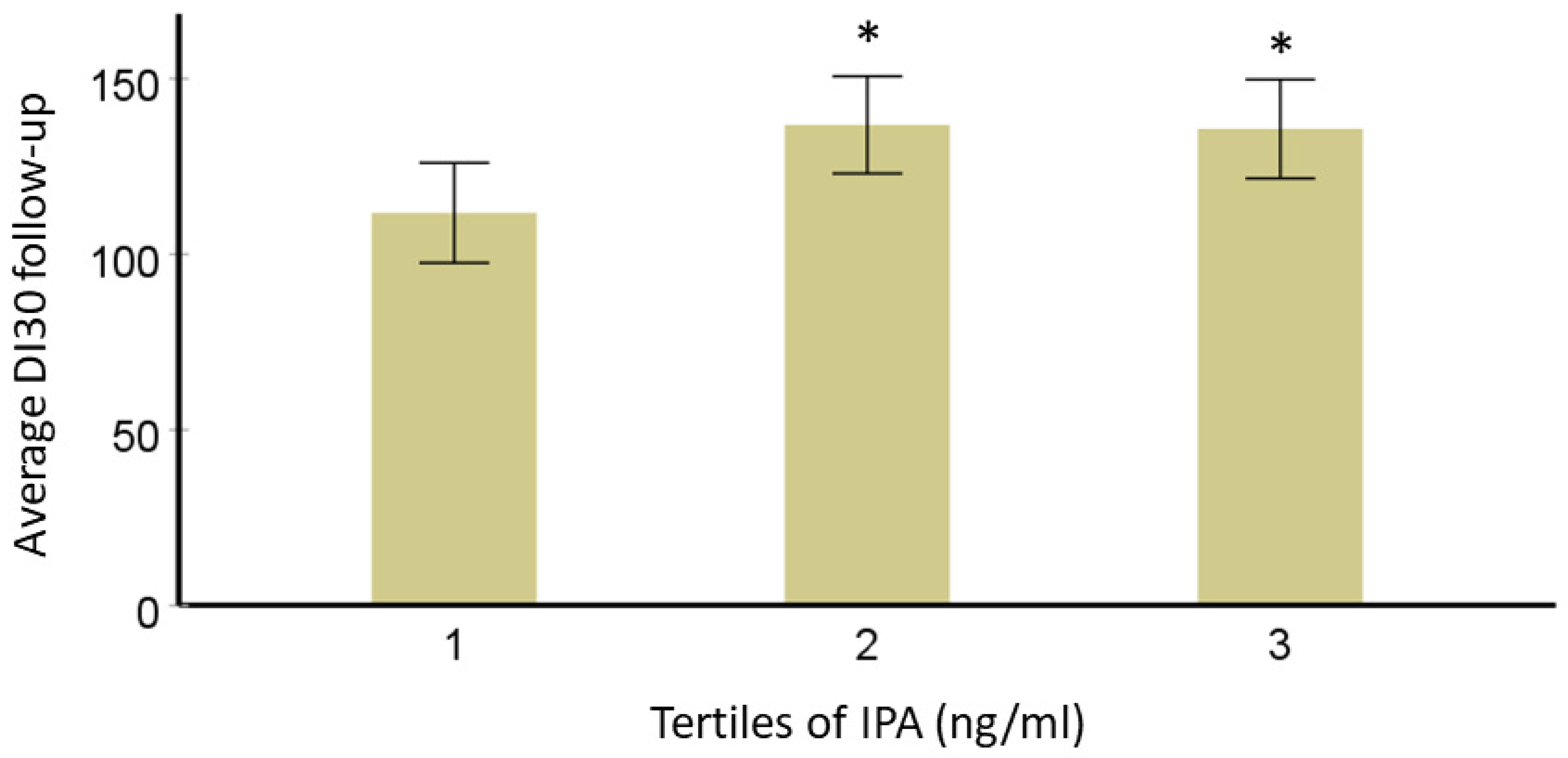
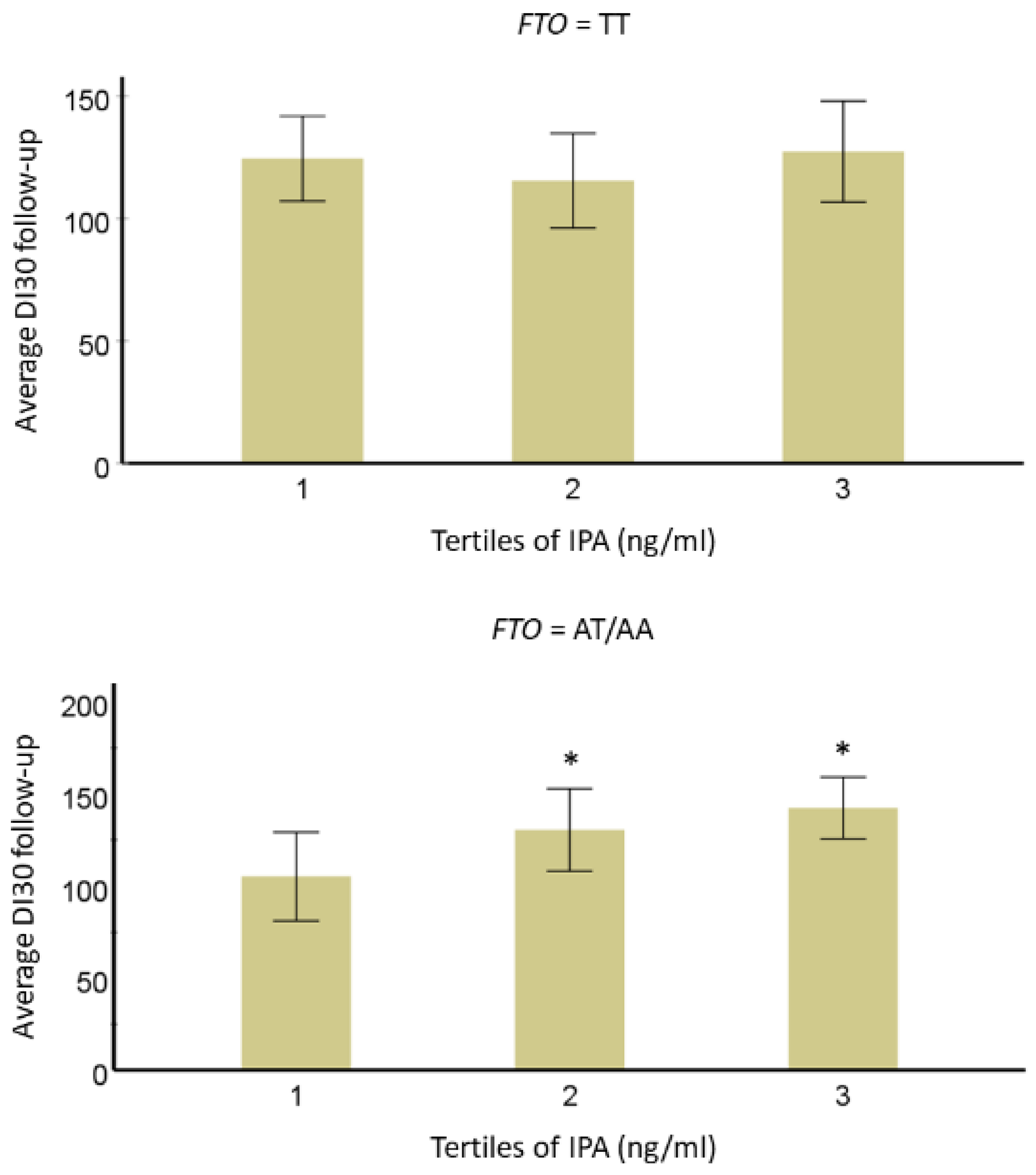
3.2. Interaction of IPA with T2D-Linked Genetic Variants
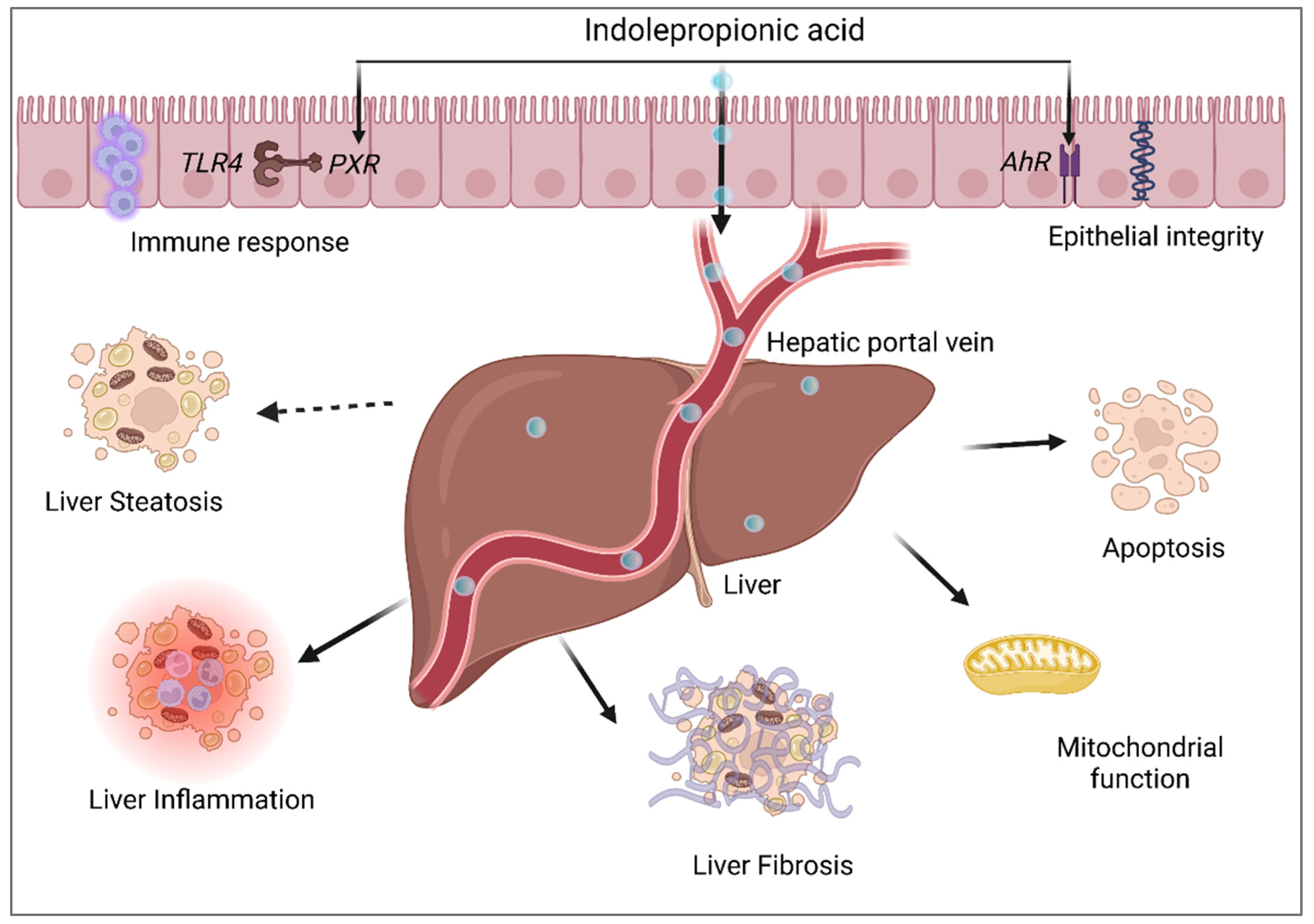
4. Indolepropionic Acid and Non-Alcoholic Fatty Liver Disease
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, F.W. The Fate of Indolepropionic Acid in the Animal Organism. Biochem. J. 1923, 17, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Bendheim, P.E.; Poeggeler, B.; Neria, E.; Ziv, V.; Pappolla, M.A.; Chain, D.G. Development of Indole-3-Propionic Acid (OXIGONTM) for Alzheimer’s Disease. J. Mol. Neurosci. 2002, 19, 213–217. [Google Scholar] [CrossRef]
- Konopelski, P.; Mogilnicka, I. Biological Effects of Indole-3-Propionic Acid, a Gut Microbiota-Derived Metabolite, and Its Precursor Tryptophan in Mammals’ Health and Disease. Int. J. Mol. Sci. 2022, 23, 1222. [Google Scholar] [CrossRef]
- Menni, C.; Hernandez, M.M.; Vital, M.; Mohney, R.P.; Spector, T.D.; Valdes, A.M. Circulating Levels of the Anti-Oxidant Indoleproprionic Acid Are Associated with Higher Gut Microbiome Diversity. Null 2019, 10, 688–695. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Goodarzi, M.O. Metabolites Linking the Gut Microbiome with Risk for Type 2 Diabetes. Curr. Nutr. Rep. 2020, 9, 83–93. [Google Scholar] [CrossRef]
- Massey, W.; Brown, J.M. The Gut Microbial Endocrine Organ in Type 2 Diabetes. Endocrinology 2021, 162, bqaa235. [Google Scholar] [CrossRef]
- De Mello, V.D.; Paananen, J.; Lindström, J.; Lankinen, M.A.; Shi, L.; Kuusisto, J.; Pihlajamäki, J.; Auriola, S.; Lehtonen, M.; Rolandsson, O.; et al. Indolepropionic Acid and Novel Lipid Metabolites Are Associated with a Lower Risk of Type 2 Diabetes in the Finnish Diabetes Prevention Study. Sci. Rep. 2017, 7, 46337. [Google Scholar] [CrossRef] [Green Version]
- Tuomainen, M.; Lindström, J.; Lehtonen, M.; Auriola, S.; Pihlajamäki, J.; Peltonen, M.; Tuomilehto, J.; Uusitupa, M.; de Mello, D.V.; Hanhineva, K. Associations of Serum Indolepropionic Acid, a Gut Microbiota Metabolite, with Type 2 Diabetes and Low-Grade Inflammation in High-Risk Individuals. Nutr. Diabetes 2018, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Jiang, M.; Zhao, J.; Song, Y.; Du, W.; Shi, J. The Mechanism Underlying the Influence of Indole-3-Propionic Acid: A Relevance to Metabolic Disorders. Front. Endocrinol. 2022, 13, 841703. [Google Scholar] [CrossRef]
- Xia, M.-F.; Bian, H.; Gao, X. NAFLD and Diabetes: Two Sides of the Same Coin? Rationale for Gene-Based Personalized NAFLD Treatment. Front. Pharmacol. 2019, 10, 877. [Google Scholar] [CrossRef] [PubMed]
- Burkitt, D.P. Related Disease--Related Cause? Lancet 1969, 294, 1229–1231. [Google Scholar] [CrossRef]
- Cummings, J.H.; Engineer, A. Denis Burkitt and the Origins of the Dietary Fibre Hypothesis. Nutr. Res. Rev. 2018, 31, 1–15. [Google Scholar] [CrossRef] [Green Version]
- O’Keefe, S.J. The Association between Dietary Fibre Deficiency and High-Income Lifestyle-Associated Diseases: Burkitt’s Hypothesis Revisited. Lancet Gastroenterol. Hepatol. 2019, 4, 984–996. [Google Scholar] [CrossRef]
- Lindström, J.; Peltonen, M.; Eriksson, J.G.; Louheranta, A.; Fogelholm, M.; Uusitupa, M.; Tuomilehto, J. High-Fibre, Low-Fat Diet Predicts Long-Term Weight Loss and Decreased Type 2 Diabetes Risk: The Finnish Diabetes Prevention Study. Diabetologia 2006, 49, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Abildgaard, A.; Elfving, B.; Hokland, M.; Wegener, G.; Lund, S. The Microbial Metabolite Indole-3-Propionic Acid Improves Glucose Metabolism in Rats, but Does Not Affect Behaviour. Null 2018, 124, 306–312. [Google Scholar] [CrossRef]
- Dodd, D.; Spitzer, M.H.; Van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A Gut Bacterial Pathway Metabolizes Aromatic Amino Acids into Nine Circulating Metabolites. Nature 2017, 551, 648–652. [Google Scholar] [CrossRef] [Green Version]
- Negatu, D.A.; Gengenbacher, M.; Dartois, V.; Dick, T. Indole Propionic Acid, an Unusual Antibiotic Produced by the Gut Microbiota, With Anti-Inflammatory and Antioxidant Properties. Front. Microbiol. 2020, 11, 575586. [Google Scholar] [CrossRef]
- Serger, E.; Luengo-Gutierrez, L.; Chadwick, J.S.; Kong, G.; Zhou, L.; Crawford, G.; Danzi, M.C.; Myridakis, A.; Brandis, A.; Bello, A.T.; et al. The Gut Metabolite Indole-3 Propionate Promotes Nerve Regeneration and Repair. Nature 2022, 607, 585–592. [Google Scholar] [CrossRef]
- Sehgal, R.; Ilha, M.; Vaittinen, M.; Kaminska, D.; Männistö, V.; Kärjä, V.; Tuomainen, M.; Hanhineva, K.; Romeo, S.; Pajukanta, P.; et al. Indole-3-Propionic Acid, a Gut-Derived Tryptophan Metabolite, Associates with Hepatic Fibrosis. Nutrients 2021, 13, 3509. [Google Scholar] [CrossRef]
- Uusitupa, M.; Louheranta, A.; Lindström, J.; Valle, T.; Sundvall, J.; Eriksson, J.; Tuomilehto, J. The Finnish Diabetes Prevention Study. Br. J. Nutr. 2000, 83 (Suppl. S1), S137–S142. [Google Scholar] [CrossRef]
- Palego, L.; Betti, L.; Rossi, A.; Giannaccini, G. Tryptophan Biochemistry: Structural, Nutritional, Metabolic, and Medical Aspects in Humans. J. Amino Acids 2016, 2016, 8952520. [Google Scholar] [CrossRef] [Green Version]
- Hendrikx, T.; Schnabl, B. Antimicrobial Proteins: Intestinal Guards to Protect against Liver Disease. J. Gastroenterol. 2019, 54, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Roager, H.M.; Licht, T.R. Microbial Tryptophan Catabolites in Health and Disease. Nat. Commun. 2018, 9, 3210–3294. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Qi, R.; Wang, J.; Liu, Z.; Wu, Z. Indole-3-Propionic Acid, a Functional Metabolite of Clostridium Sporogenes, Promotes Muscle Tissue Development and Reduces Muscle Cell Inflammation. Int. J. Mol. Sci. 2021, 22, 12435. [Google Scholar] [CrossRef]
- Nishizawa, T.; Aldrich, C.C.; Sherman, D.H. Molecular Analysis of the Rebeccamycin L-Amino Acid Oxidase from Lechevalieria Aerocolonigenes ATCC 39243. J. Bacteriol. 2005, 187, 2084–2092. [Google Scholar] [CrossRef] [Green Version]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe 2018, 23, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Melhem, N.J.; Taleb, S. Tryptophan: From Diet to Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 9904. [Google Scholar] [CrossRef]
- Arnoriaga-Rodríguez, M.; Mayneris-Perxachs, J.; Burokas, A.; Pérez-Brocal, V.; Moya, A.; Portero-Otin, M.; Ricart, W.; Maldonado, R.; Fernández-Real, J.-M. Gut Bacterial ClpB-like Gene Function Is Associated with Decreased Body Weight and a Characteristic Microbiota Profile. Microbiome 2020, 8, 59. [Google Scholar] [CrossRef]
- Michonneau, D.; Latis, E.; Curis, E.; Dubouchet, L.; Ramamoorthy, S.; Ingram, B.; de Latour, R.P.; Robin, M.; de Fontbrune, F.S.; Chevret, S.; et al. Metabolomics Analysis of Human Acute Graft-versus-Host Disease Reveals Changes in Host and Microbiota-Derived Metabolites. Nat. Commun. 2019, 10, 5695. [Google Scholar] [CrossRef] [Green Version]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity via Interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics Analysis Reveals Large Effects of Gut Microflora on Mammalian Blood Metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic Bacterial Metabolites Regulate Gastrointestinal Barrier Function via the Xenobiotic Sensor PXR and Toll-like Receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef] [Green Version]
- Jennis, M.; Cavanaugh, C.R.; Leo, G.C.; Mabus, J.R.; Lenhard, J.; Hornby, P.J. Microbiota-Derived Tryptophan Indoles Increase after Gastric Bypass Surgery and Reduce Intestinal Permeability in Vitro and in Vivo. Neurogastroenterol. Motil. 2018, 30, e13178. [Google Scholar] [CrossRef]
- Van der Lelie, D.; Oka, A.; Taghavi, S.; Umeno, J.; Fan, T.-J.; Merrell, K.E.; Watson, S.D.; Ouellette, L.; Liu, B.; Awoniyi, M.; et al. Rationally Designed Bacterial Consortia to Treat Chronic Immune-Mediated Colitis and Restore Intestinal Homeostasis. Nat. Commun. 2021, 12, 3105. [Google Scholar] [CrossRef]
- Alexeev, E.E.; Lanis, J.M.; Kao, D.J.; Campbell, E.L.; Kelly, C.J.; Battista, K.D.; Gerich, M.E.; Jenkins, B.R.; Walk, S.T.; Kominsky, D.J.; et al. Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 2018, 188, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Dong, F.; Hao, F.; Murray, I.A.; Smith, P.B.; Koo, I.; Tindall, A.M.; Kris-Etherton, P.M.; Gowda, K.; Amin, S.G.; Patterson, A.D.; et al. Intestinal Microbiota-Derived Tryptophan Metabolites Are Predictive of Ah Receptor Activity. Gut Microbes 2020, 12, 1788899. [Google Scholar] [CrossRef]
- Cao, B.; Zhao, R.-Y.; Li, H.-H.; Xu, X.-M.; Cui, H.; Deng, H.; Chen, L.; Wei, B. Oral Administration of Asparagine and 3-Indolepropionic Acid Prolongs Survival Time of Rats with Traumatic Colon Injury. Mil. Med. Res. 2022, 9, 37. [Google Scholar] [CrossRef]
- Lee-Sarwar, K.; Lasky-Su, J.; Kelly, R.S.; Litonjua, A.A.; Weiss, S.T. Metabolome-Microbiome Crosstalk and Human Disease. Metabolites 2020, 10, 181. [Google Scholar] [CrossRef]
- Liu, Z.; Dai, X.; Zhang, H.; Shi, R.; Hui, Y.; Jin, X.; Zhang, W.; Wang, L.; Wang, Q.; Wang, D.; et al. Gut Microbiota Mediates Intermittent-Fasting Alleviation of Diabetes-Induced Cognitive Impairment. Nat. Commun. 2020, 11, 855. [Google Scholar] [CrossRef] [Green Version]
- Levi, I.; Gurevich, M.; Perlman, G.; Magalashvili, D.; Menascu, S.; Bar, N.; Godneva, A.; Zahavi, L.; Chermon, D.; Kosower, N.; et al. Potential Role of Indolelactate and Butyrate in Multiple Sclerosis Revealed by Integrated Microbiome-Metabolome Analysis. Cell Rep. Med. 2021, 2, 100246. [Google Scholar] [CrossRef]
- Grifka-Walk, H.M.; Jenkins, B.R.; Kominsky, D.J. Amino Acid Trp: The Far Out Impacts of Host and Commensal Tryptophan Metabolism. Front. Immunol. 2021, 12, 653208. [Google Scholar] [CrossRef] [PubMed]
- Koshiba, S.; Motoike, I.N.; Saigusa, D.; Inoue, J.; Aoki, Y.; Tadaka, S.; Shirota, M.; Katsuoka, F.; Tamiya, G.; Minegishi, N.; et al. Identification of Critical Genetic Variants Associated with Metabolic Phenotypes of the Japanese Population. Commun. Biol. 2020, 3, 662. [Google Scholar] [CrossRef] [PubMed]
- Gesper, M.; Nonnast, A.B.H.; Kumowski, N.; Stoehr, R.; Schuett, K.; Marx, N.; Kappel, B.A. Gut-Derived Metabolite Indole-3-Propionic Acid Modulates Mitochondrial Function in Cardiomyocytes and Alters Cardiac Function. Front. Med. 2021, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Li, J.; Yu, B.; Moon, J.-Y.; Chai, J.C.; Merino, J.; Hu, J.; Ruiz-Canela, M.; Rebholz, C.; Wang, Z.; et al. Host and Gut Microbial Tryptophan Metabolism and Type 2 Diabetes: An Integrative Analysis of Host Genetics, Diet, Gut Microbiome and Circulating Metabolites in Cohort Studies. Gut 2022, 71, 1095–1105. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of Gut Microbiota in Type 2 Diabetes Pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [Green Version]
- Salgaço, M.K.; Oliveira, L.G.S.; Costa, G.N.; Bianchi, F.; Sivieri, K. Relationship between Gut Microbiota, Probiotics, and Type 2 Diabetes Mellitus. Appl. Microbiol. Biotechnol. 2019, 103, 9229–9238. [Google Scholar] [CrossRef]
- Zhu, C.; Sawrey-Kubicek, L.; Beals, E.; Rhodes, C.H.; Houts, H.E.; Sacchi, R.; Zivkovic, A.M. Human Gut Microbiome Composition and Tryptophan Metabolites Were Changed Differently by Fast Food and Mediterranean Diet in 4 Days: A Pilot Study. Nutr. Res. 2020, 77, 62–72. [Google Scholar] [CrossRef]
- Li, Y.; Xu, W.; Zhang, F.; Zhong, S.; Sun, Y.; Huo, J.; Zhu, J.; Wu, C. The Gut Microbiota-Produced Indole-3-Propionic Acid Confers the Antihyperlipidemic Effect of Mulberry-Derived 1-Deoxynojirimycin. mSystems 2020, 5, e00313-20. [Google Scholar] [CrossRef]
- Kundi, Z.M.; Lee, J.C.-Y.; Pihlajamäki, J.; Chan, C.B.; Leung, K.S.; So, S.S.Y.; Nordlund, E.; Kolehmainen, M.; El-Nezami, H. Dietary Fiber from Oat and Rye Brans Ameliorate Western Diet–Induced Body Weight Gain and Hepatic Inflammation by the Modulation of Short-Chain Fatty Acids, Bile Acids, and Tryptophan Metabolism. Mol. Nutr. Food Res. 2021, 65, 1900580. [Google Scholar] [CrossRef]
- Ferrere, G.; Tidjani Alou, M.; Liu, P.; Goubet, A.-G.; Fidelle, M.; Kepp, O.; Durand, S.; Iebba, V.; Fluckiger, A.; Daillère, R.; et al. Ketogenic Diet and Ketone Bodies Enhance the Anticancer Effects of PD-1 Blockade. JCI Insight 2021, 6, e145207. [Google Scholar] [CrossRef]
- Gao, J.; Guo, X.; Wei, W.; Li, R.; Hu, K.; Liu, X.; Jiang, W.; Liu, S.; Wang, W.; Sun, H.; et al. The Association of Fried Meat Consumption With the Gut Microbiota and Fecal Metabolites and Its Impact on Glucose Homoeostasis, Intestinal Endotoxin Levels, and Systemic Inflammation: A Randomized Controlled-Feeding Trial. Diabetes Care 2021, 44, 1970–1979. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, L.; Xia, B.; Tang, S.; Liu, L.; Xie, J.; Zhang, H. Bioregional Alterations in Gut Microbiome Contribute to the Plasma Metabolomic Changes in Pigs Fed with Inulin. Microorganisms 2020, 8, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimentel, G.; Burton, K.J.; von Ah, U.; Bütikofer, U.; Pralong, F.P.; Vionnet, N.; Portmann, R.; Vergères, G. Metabolic Footprinting of Fermented Milk Consumption in Serum of Healthy Men. J. Nutr. 2018, 148, 851–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellis, L.; van Erk, M.J.; van Ommen, B.; Bakker, G.C.M.; Hendriks, H.F.J.; Cnubben, N.H.P.; Kleemann, R.; van Someren, E.P.; Bobeldijk, I.; Rubingh, C.M.; et al. Plasma Metabolomics and Proteomics Profiling after a Postprandial Challenge Reveal Subtle Diet Effects on Human Metabolic Status. Metabolomics 2012, 8, 347–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, S.; Yang, J.; Lee, R.; Huang, J.; Small, G.; Guo, Y.; Heber, D.; Li, Z. Effect of Pomegranate Juice Consumption on Plasma Indole Propionic Acid, a Bacterial-Derived Tryptophan Metabolite (P20-003-19). Curr. Dev. Nutr. 2019, 3 (Suppl. S1), nzz040-P20. [Google Scholar] [CrossRef] [Green Version]
- Peron, G.; Meroño, T.; Gargari, G.; Hidalgo-Liberona, N.; Miñarro, A.; Lozano, E.V.; Castellano-Escuder, P.; González-Domínguez, R.; del Bo’, C.; Bernardi, S.; et al. A Polyphenol-Rich Diet Increases the Gut Microbiota Metabolite Indole 3-Propionic Acid in Older Adults with Preserved Kidney Function. Mol. Nutr. Food Res. 2022, 66, 2100349. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Y.; Sun, S.; Xie, Y.; Pan, C.; Li, M.; Li, C.; Liu, Y.; Xu, Z.; Liu, W.; et al. Indolepropionic Acid Reduces Obesity-Induced Metabolic Dysfunction through Colonic Barrier Restoration Mediated via Tuft Cell-Derived IL-25. FEBS J. 2022, 289, 5985–6004. [Google Scholar] [CrossRef]
- Dong, T.S.; Mayer, E.A.; Osadchiy, V.; Chang, C.; Katzka, W.; Lagishetty, V.; Gonzalez, K.; Kalani, A.; Stains, J.; Jacobs, J.P.; et al. A Distinct Brain-Gut-Microbiome Profile Exists for Females with Obesity and Food Addiction. Obesity 2020, 28, 1477–1486. [Google Scholar] [CrossRef]
- Gou, W.; Ling, C.-W.; He, Y.; Jiang, Z.; Fu, Y.; Fengzhe, X.; Miao, Z.-L.; Ting-yu, S.; Jie-Sheng, L.; Zhu, H.; et al. Interpretable Machine Learning Algorithm Reveals Novel Gut Microbiome Features in Predicting Type 2 Diabetes. Curr. Dev. Nutr. 2020, 4, 1559. [Google Scholar] [CrossRef]
- Gudmundsdottir, V.; Pedersen, H.K.; Mazzoni, G.; Allin, K.H.; Artati, A.; Beulens, J.W.; Banasik, K.; Brorsson, C.; Cederberg, H.; Chabanova, E.; et al. Whole Blood Co-Expression Modules Associate with Metabolic Traits and Type 2 Diabetes: An IMI-DIRECT Study. Genome Med. 2020, 12, 109. [Google Scholar] [CrossRef]
- Morze, J.; Wittenbecher, C.; Schwingshackl, L.; Danielewicz, A.; Rynkiewicz, A.; Hu, F.B.; Guasch-Ferré, M. Metabolomics and Type 2 Diabetes Risk: An Updated Systematic Review and Meta-Analysis of Prospective Cohort Studies. Diabetes Care 2022, 45, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Lustgarten, M.S.; Price, L.L.; Chale, A.; Phillips, E.M.; Fielding, R.A. Branched Chain Amino Acids Are Associated with Muscle Mass in Functionally Limited Older Adults. J. Gerontol. Biol. Sci. Med. Sci. 2014, 69, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seridi, L.; Leo, G.C.; Dohm, G.L.; Pories, W.J.; Lenhard, J. Time Course Metabolome of Roux-En-Y Gastric Bypass Confirms Correlation between Leptin, Body Weight and the Microbiome. PLoS ONE 2018, 13, e0198156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangipurapu, J.; Fernandes Silva, L.; Kuulasmaa, T.; Smith, U.; Laakso, M. Microbiota-Related Metabolites and the Risk of Type 2 Diabetes. Diabetes Care 2020, 43, 1319–1325. [Google Scholar] [CrossRef]
- Kemppainen, S.M.; Fernandes Silva, L.; Lankinen, M.A.; Schwab, U.; Laakso, M. Metabolite Signature of Physical Activity and the Risk of Type 2 Diabetes in 7271 Men. Metabolites 2022, 12, 69. [Google Scholar] [CrossRef]
- Tan, Y.M.; Gao, Y.; Teo, G.; Koh, H.W.L.; Tai, E.S.; Khoo, C.M.; Choi, K.P.; Zhou, L.; Choi, H. Plasma Metabolome and Lipidome Associations with Type 2 Diabetes and Diabetic Nephropathy. Metabolites 2021, 11, 228. [Google Scholar] [CrossRef]
- Gundu, C.; Arruri, V.K.; Sherkhane, B.; Khatri, D.K.; Singh, S.B. Indole-3-Propionic Acid Attenuates High Glucose Induced ER Stress Response and Augments Mitochondrial Function by Modulating PERK-IRE1-ATF4-CHOP Signalling in Experimental Diabetic Neuropathy. Arch. Physiol. Biochem. 2022, 1–14. [Google Scholar] [CrossRef]
- Mercer, K.E.; Yeruva, L.; Pack, L.; Graham, J.L.; Stanhope, K.L.; Chintapalli, S.V.; Wankhade, U.D.; Shankar, K.; Havel, P.J.; Adams, S.H.; et al. Xenometabolite Signatures in the UC Davis Type 2 Diabetes Mellitus Rat Model Revealed Using a Metabolomics Platform Enriched with Microbe-Derived Metabolites. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G157–G169. [Google Scholar] [CrossRef]
- Kuhn, B.; Hilpert, H.; Benz, J.; Binggeli, A.; Grether, U.; Humm, R.; Märki, H.P.; Meyer, M.; Mohr, P. Structure-Based Design of Indole Propionic Acids as Novel PPARα/γ Co-Agonists. Bioorg. Med. Chem. Lett. 2006, 16, 4016–4020. [Google Scholar] [CrossRef]
- Konopelski, P.; Konop, M.; Gawrys-Kopczynska, M.; Podsadni, P.; Szczepanska, A.; Ufnal, M. Indole-3-Propionic Acid, a Tryptophan-Derived Bacterial Metabolite, Reduces Weight Gain in Rats. Nutrients 2019, 11, 591. [Google Scholar] [CrossRef]
- Shin, S.-Y.; Fauman, E.B.; Petersen, A.-K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.-P.; et al. An Atlas of Genetic Influences on Human Blood Metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Kuusisto, J.; Vänttinen, M.; Kuulasmaa, T.; Lindström, J.; Tuomilehto, J.; Uusitupa, M.; Laakso, M. Variants of Transcription Factor 7-like 2 (TCF7L2) Gene Predict Conversion to Type 2 Diabetes in the Finnish Diabetes Prevention Study and Are Associated with Impaired Glucose Regulation and Impaired Insulin Secretion. Diabetologia 2007, 50, 1192–1200. [Google Scholar] [CrossRef] [Green Version]
- Lappalainen, T.J.; Tolppanen, A.-M.; Kolehmainen, M.; Schwab, U.; Lindström, J.; Tuomilehto, J.; Pulkkinen, L.; Eriksson, J.G.; Laakso, M.; Gylling, H.; et al. The Common Variant in the FTO Gene Did Not Modify the Effect of Lifestyle Changes on Body Weight: The Finnish Diabetes Prevention Study. Obesity 2009, 17, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Lindi, V.I.; Uusitupa, M.I.J.; Lindström, J.; Louheranta, A.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; et al. Association of the Pro12Ala Polymorphism in the PPAR-Γ2 Gene With 3-Year Incidence of Type 2 Diabetes and Body Weight Change in the Finnish Diabetes Prevention Study. Diabetes 2002, 51, 2581–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindström, J.; Peltonen, M.; Eriksson, J.G.; Ilanne-Parikka, P.; Aunola, S.; Keinänen-Kiukaanniemi, S.; Uusitupa, M.; Tuomilehto, J. Improved lifestyle and decreased diabetes risk over 13 years: Long-term follow-up of the randomised Finnish Diabetes Prevention Study (DPS). Diabetologia 2013, 56, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stančáková, A.; Javorský, M.; Kuulasmaa, T.; Haffner, S.M.; Kuusisto, J.; Laakso, M. Changes in Insulin Sensitivity and Insulin Release in Relation to Glycemia and Glucose Tolerance in 6,414 Finnish Men. Diabetes 2009, 58, 1212–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, A.P.; Voight, B.F.; Teslovich, T.M.; Ferreira, T.; Segrè, A.V.; Steinthorsdottir, V.; Strawbridge, R.J.; Khan, H.; Grallert, H.; Mahajan, A.; et al. Large-Scale Association Analysis Provides Insights into the Genetic Architecture and Pathophysiology of Type 2 Diabetes. Nat. Genet. 2012, 44, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Wen, S.W.; Kaminga, A.C.; Liu, A. Gut Metabolites and Inflammation Factors in Non-Alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 8848. [Google Scholar] [CrossRef]
- Deprince, A.; Haas, J.T.; Staels, B. Dysregulated Lipid Metabolism Links NAFLD to Cardiovascular Disease. Mol. Metab. 2020, 42, 101092. [Google Scholar] [CrossRef]
- Hendrikx, T.; Schnabl, B. Indoles: Metabolites Produced by Intestinal Bacteria Capable of Controlling Liver Disease Manifestation. J. Intern. Med. 2019, 286, 32–40. [Google Scholar] [CrossRef]
- Frasinariu, O.; Serban, R.; Trandafir, L.M.; Miron, I.; Starcea, M.; Vasiliu, I.; Alisi, A.; Temneanu, O.R. The Role of Phytosterols in Nonalcoholic Fatty Liver Disease. Nutrients 2022, 14, 2187. [Google Scholar] [CrossRef] [PubMed]
- Simonen, M.; Männistö, V.; Leppänen, J.; Kaminska, D.; Kärjä, V.; Venesmaa, S.; Käkelä, P.; Kuusisto, J.; Gylling, H.; Laakso, M.; et al. Desmosterol in Human Nonalcoholic Steatohepatitis. Hepatology 2013, 58, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Farook, V.S.; Reddivari, L.; Chittoor, G.; Puppala, S.; Arya, R.; Fowler, S.P.; Hunt, K.J.; Curran, J.E.; Comuzzie, A.G.; Lehman, D.M.; et al. Metabolites as Novel Biomarkers for Childhood Obesity-Related Traits in Mexican-American Children. Pediatr. Obes. 2015, 10, 320–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.; Chen, X.; Yu, C.; Deng, Y.; Zhang, Y.; Chen, S.; Chen, X.; Chen, K.; Yang, Y.; Ling, W. Gut Microbially Produced Indole-3-Propionic Acid Inhibits Atherosclerosis by Promoting Reverse Cholesterol Transport and Its Deficiency Is Causally Related to Atherosclerotic Cardiovascular Disease. Circ. Res. 2022, 131, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Cason, C.A.; Dolan, K.T.; Sharma, G.; Tao, M.; Kulkarni, R.; Helenowski, I.B.; Doane, B.M.; Avram, M.J.; McDermott, M.M.; Chang, E.B.; et al. Plasma Microbiome-Modulated Indole- and Phenyl-Derived Metabolites Associate with Advanced Atherosclerosis and Postoperative Outcomes. J. Vasc. Surg. 2018, 68, 1552–1562. [Google Scholar] [CrossRef]
- Lefler, S.R.; St Peter, S.D.; Rodriguez-Davalos, M.I.; Perumean, J.C.; Spence, N.A.; Moss, A.A.; Mulligan, D.C. Amelioration of Hepatic Ischemia/Reperfusion Injury Using Indolepropionic Acid. Transplant. Proc. 2002, 34, 3065–3066. [Google Scholar] [CrossRef]
- Zhao, Z.-H.; Xin, F.-Z.; Xue, Y.; Hu, Z.; Han, Y.; Ma, F.; Zhou, D.; Liu, X.-L.; Cui, A.; Liu, Z.; et al. Indole-3-Propionic Acid Inhibits Gut Dysbiosis and Endotoxin Leakage to Attenuate Steatohepatitis in Rats. Exp. Mol. Med. 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Cabrera, J.; Burkhardt, S.; Osuna, C.; Lewiński, A. Indole-3-Propionic Acid, a Melatonin-Related Molecule, Protects Hepatic Microsomal Membranes from Iron-Induced Oxidative Damage: Relevance to Cancer Reduction. J. Cell. Biochem. 2001, 81, 507–513. [Google Scholar] [CrossRef]
- De Oca, A.P.M.; Julián, M.T.; Ramos, A.; Puig-Domingo, M.; Alonso, N. Microbiota, Fiber, and NAFLD: Is There Any Connection? Nutrients 2020, 12, 3100. [Google Scholar] [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Jankó, L.; Csonka, T.; Lente, G.; Sebő, É.; Tóth, J.; Tóth, D.; Árkosy, P.; et al. Indolepropionic Acid, a Metabolite of the Microbiome, Has Cytostatic Properties in Breast Cancer by Activating AHR and PXR Receptors and Inducing Oxidative Stress. Cancers 2020, 12, 2411. [Google Scholar] [CrossRef]
- Liu, D.; Wang, J.; Zeng, H.; Zhou, F.; Wen, B.; Zhang, X.; Luo, Y.; Wu, W.; Huang, J.; Liu, Z. The Metabolic Regulation of Fuzhuan Brick Tea in High-Fat Diet-Induced Obese Mice and the Potential Contribution of Gut Microbiota. Food Funct. 2022, 13, 356–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Coker, O.O.; Chu, E.S.H.; Fu, K.; Lau, H.C.H.; Wang, Y.-X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary Cholesterol Drives Fatty Liver-Associated Liver Cancer by Modulating Gut Microbiota and Metabolites. Gut 2021, 70, 761. [Google Scholar] [CrossRef] [PubMed]
- Mazagova, M.; Wang, L.; Anfora, A.T.; Wissmueller, M.; Lesley, S.A.; Miyamoto, Y.; Eckmann, L.; Dhungana, S.; Pathmasiri, W.; Sumner, S.; et al. Commensal Microbiota Is Hepatoprotective and Prevents Liver Fibrosis in Mice. FASEB J. 2015, 29, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owumi, S.E.; Najophe, E.S.; Otunla, M.T. 3-Indolepropionic Acid Prevented Chlorpyrifos-Induced Hepatorenal Toxicities in Rats by Improving Anti-Inflammatory, Antioxidant, and pro-Apoptotic Responses and Abating DNA Damage. Environ. Sci. Pollut. Res. Int. 2022, 29, 74377–74393. [Google Scholar] [CrossRef] [PubMed]
- Owumi, S.E.; Otunla, M.T.; Arunsi, U.O.; Najophe, E.S. 3-Indolepropionic Acid Upturned Male Reproductive Function by Reducing Oxido-Inflammatory Responses and Apoptosis along the Hypothalamic-Pituitary-Gonadal Axis of Adult Rats Exposed to Chlorpyrifos. Toxicology 2021, 463, 152996. [Google Scholar] [CrossRef]
- Ren, Z.; Chen, X.; Hong, L.; Zhao, X.; Cui, G.; Li, A.; Liu, Y.; Zhou, L.; Sun, R.; Shen, S.; et al. Nanoparticle Conjugation of Ginsenoside Rg3 Inhibits Hepatocellular Carcinoma Development and Metastasis. Small 2020, 16, e1905233. [Google Scholar] [CrossRef] [Green Version]
- Yisireyili, M.; Takeshita, K.; Saito, S.; Murohara, T.; Niwa, T. Indole-3-Propionic Acid Suppresses Indoxyl Sulfate-Induced Expression of Fibrotic and Inflammatory Genes in Proximal Tubular Cells. Nagoya J. Med. Sci. 2017, 79, 477–486. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Experimental Model | Result | Molecular Mechanism | Reference |
|---|---|---|---|---|
| T2D-induced cognitive impairment | db/db mice; i.p. IPA (10 mg/Kg/day)-2 weeks | Improved insulin sensitivity and mitochondrial biogenesis | Energy metabolism via mitochondrial function | [39] |
| Obesity | High-fat diet mice p.o. IPA (100 mg/Kg) from week 7 to 14 after a high-fat diet and NIH3T3 cells (IPA 100 µM) | Reduced body weight, increased energy expenditure, glucose clearance, and insulin sensitivity, and reduced lipid accumulation in serum and liver | Reduced TNFα, TLR4 expression in adipocytes, Increases IL-25 production in tuft cells via FFAR3 signaling | [57] |
| Obesity | Sprague-Dawley i.p. IPA (30 mg/kg) for 1 week | Reduced body weight gain | Not proposed | [70] |
| Hyperlipidemia | ICR mice p.o. IPA (100 mg/kg) for 60 days | Reduced body weight gain and serum lipids | Reduced expression of SREBP1c, FAS, SREBP2, and HMGR | [48] |
| T2D and obesity | DIO mice IPA (20 mg/kg) for 4 days and T84 cells (IPA—5 µmol/L) | Increased intestinal permeability and reduced cytokine-induced changes | Reduced GLUT5 in both and ALDOB gene expression in mice | [33] |
| Insulin resistance | Sprague-Dawley rats IPA in diet (27.3 mg/Kg/day) for 6 weeks | Reduced glucose, insulin, and HOMA-IR levels | Glucose uptake | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehgal, R.; de Mello, V.D.; Männistö, V.; Lindström, J.; Tuomilehto, J.; Pihlajamäki, J.; Uusitupa, M. Indolepropionic Acid, a Gut Bacteria-Produced Tryptophan Metabolite and the Risk of Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease. Nutrients 2022, 14, 4695. https://doi.org/10.3390/nu14214695
Sehgal R, de Mello VD, Männistö V, Lindström J, Tuomilehto J, Pihlajamäki J, Uusitupa M. Indolepropionic Acid, a Gut Bacteria-Produced Tryptophan Metabolite and the Risk of Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease. Nutrients. 2022; 14(21):4695. https://doi.org/10.3390/nu14214695
Chicago/Turabian StyleSehgal, Ratika, Vanessa D. de Mello, Ville Männistö, Jaana Lindström, Jaakko Tuomilehto, Jussi Pihlajamäki, and Matti Uusitupa. 2022. "Indolepropionic Acid, a Gut Bacteria-Produced Tryptophan Metabolite and the Risk of Type 2 Diabetes and Non-Alcoholic Fatty Liver Disease" Nutrients 14, no. 21: 4695. https://doi.org/10.3390/nu14214695