
Effect of Black Corn Anthocyanin-Rich Extract (Zea mays L.) on Cecal Microbial Populations In Vivo (Gallus gallus)
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
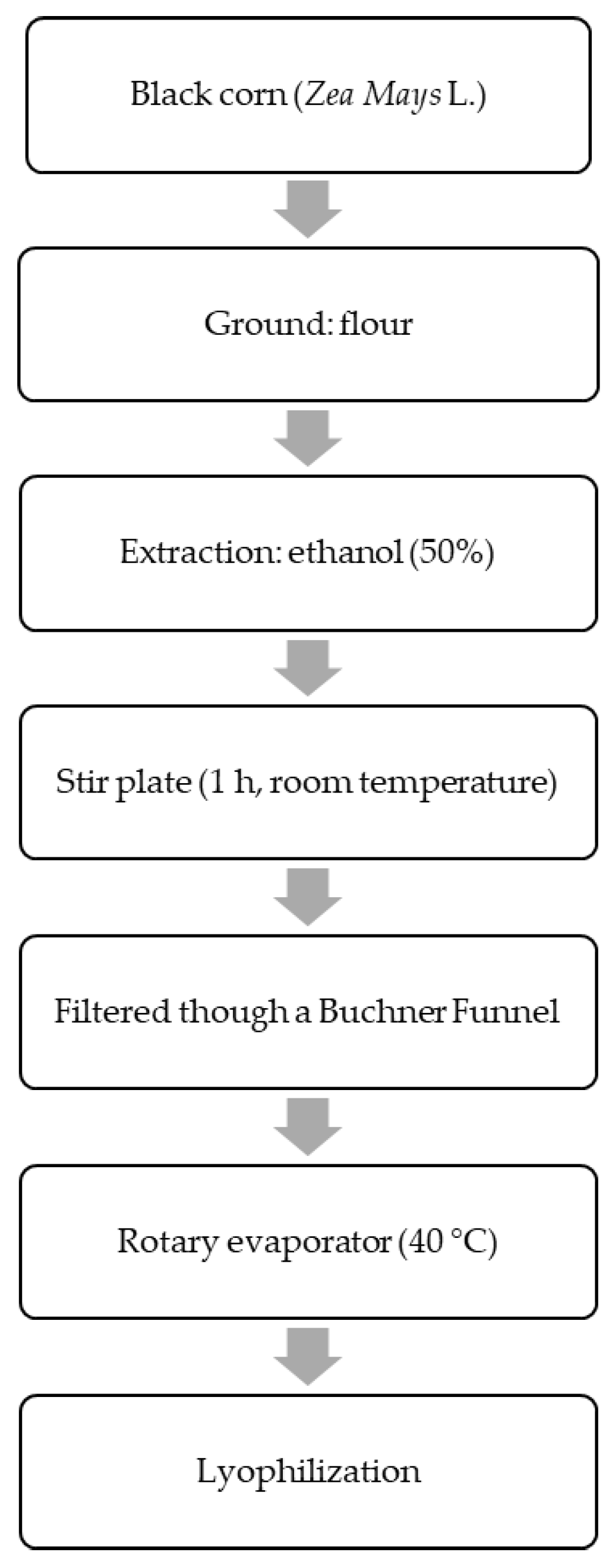
2.2. Black Corn Extract Procedure
2.3. Extract Chemical Characterization
2.3.1. Total Polyphenols and Antioxidant Capacity
2.3.2. Anthocyanin Profile Analysis
2.4. Intra-Amniotic Experiment
2.5. Total RNA Extraction from Duodenum
2.6. Gene Expression Analysis
2.7. Intestinal Content and DNA Isolation
2.8. Primers Design and PCR Amplification of Bacterial 16S rDNA
2.9. Histological Analysis
2.10. Statistical Analysis
3. Results
3.1. Black Corn Extract Characterization
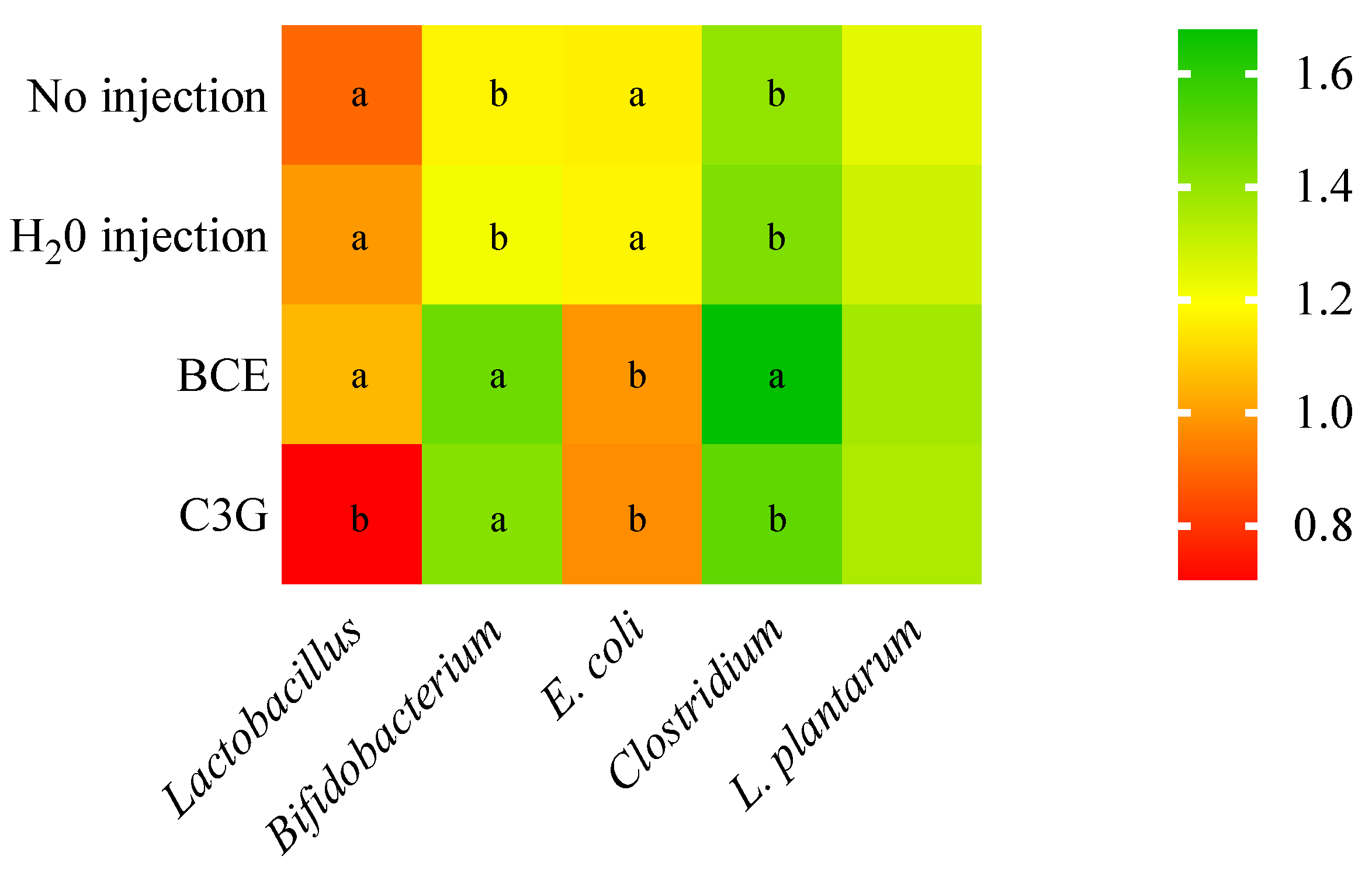
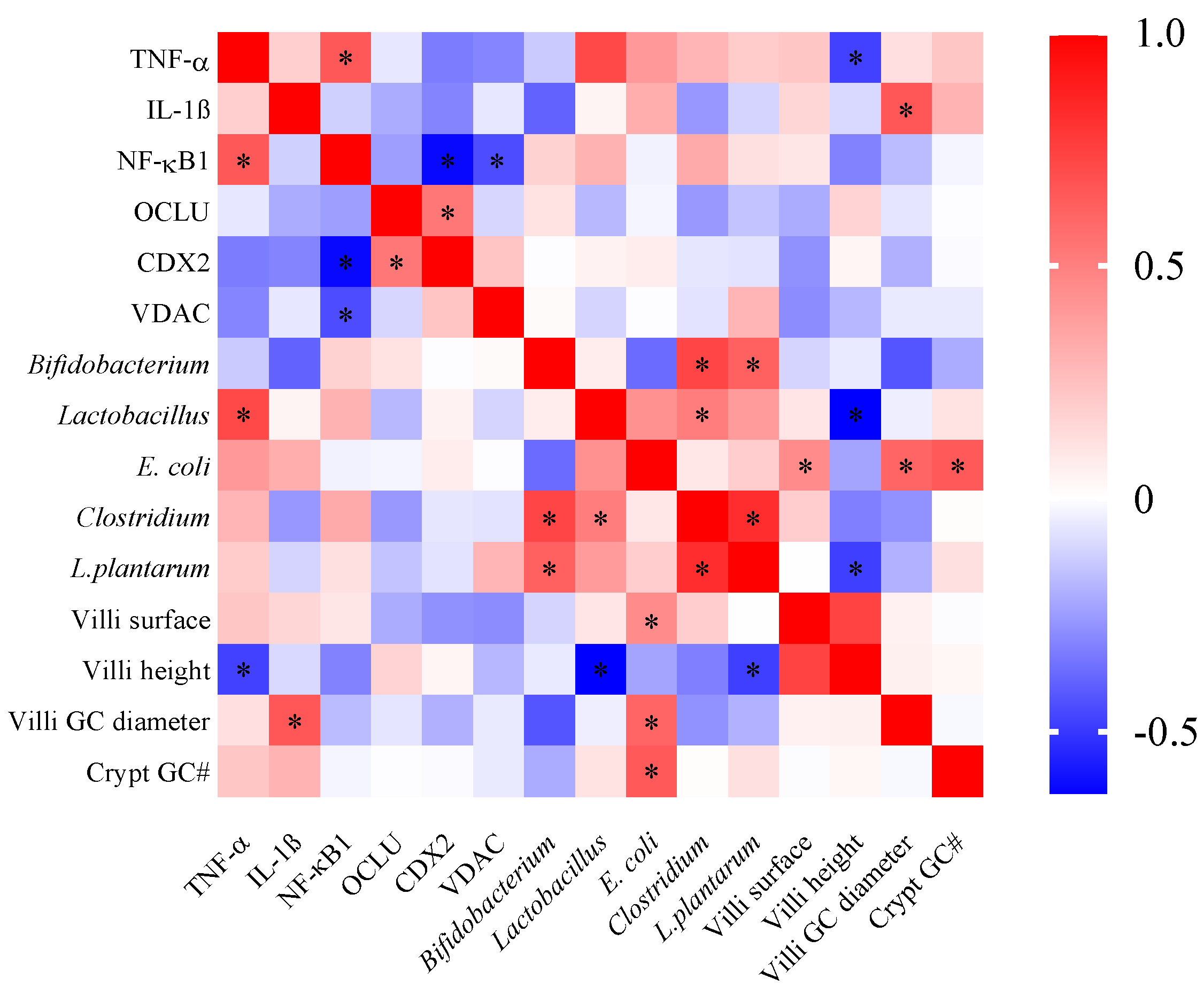
3.2. Effect of BCE on the Bacterial Population on Cecum Content
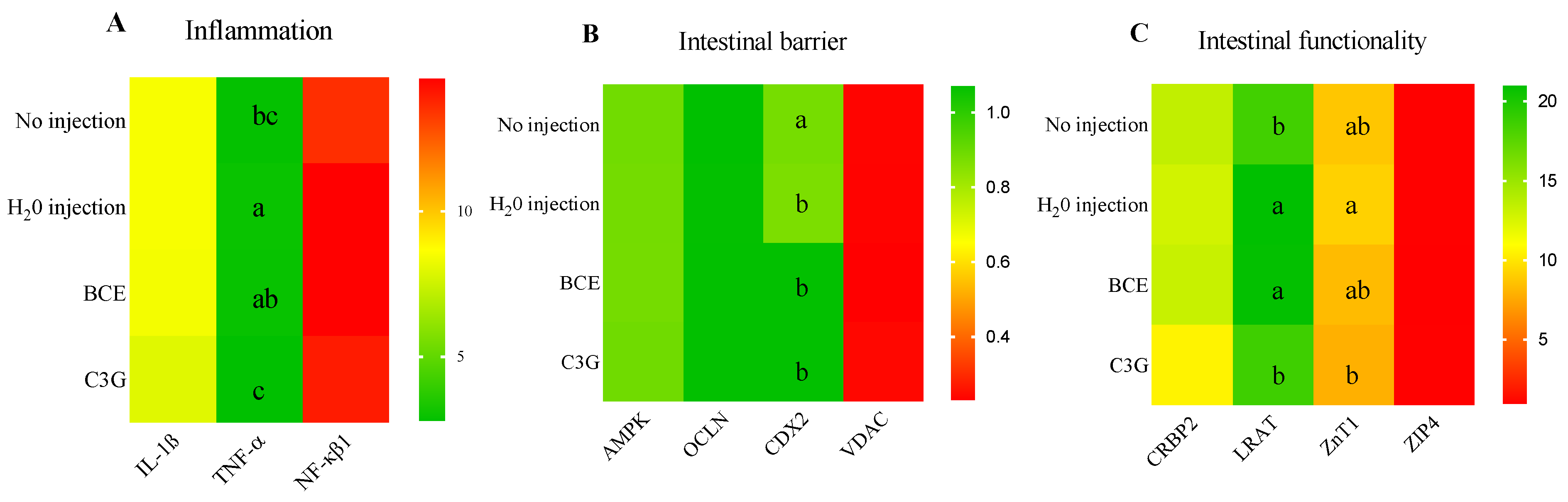
3.3. Effect of BCE on Duodenal Gene Expression
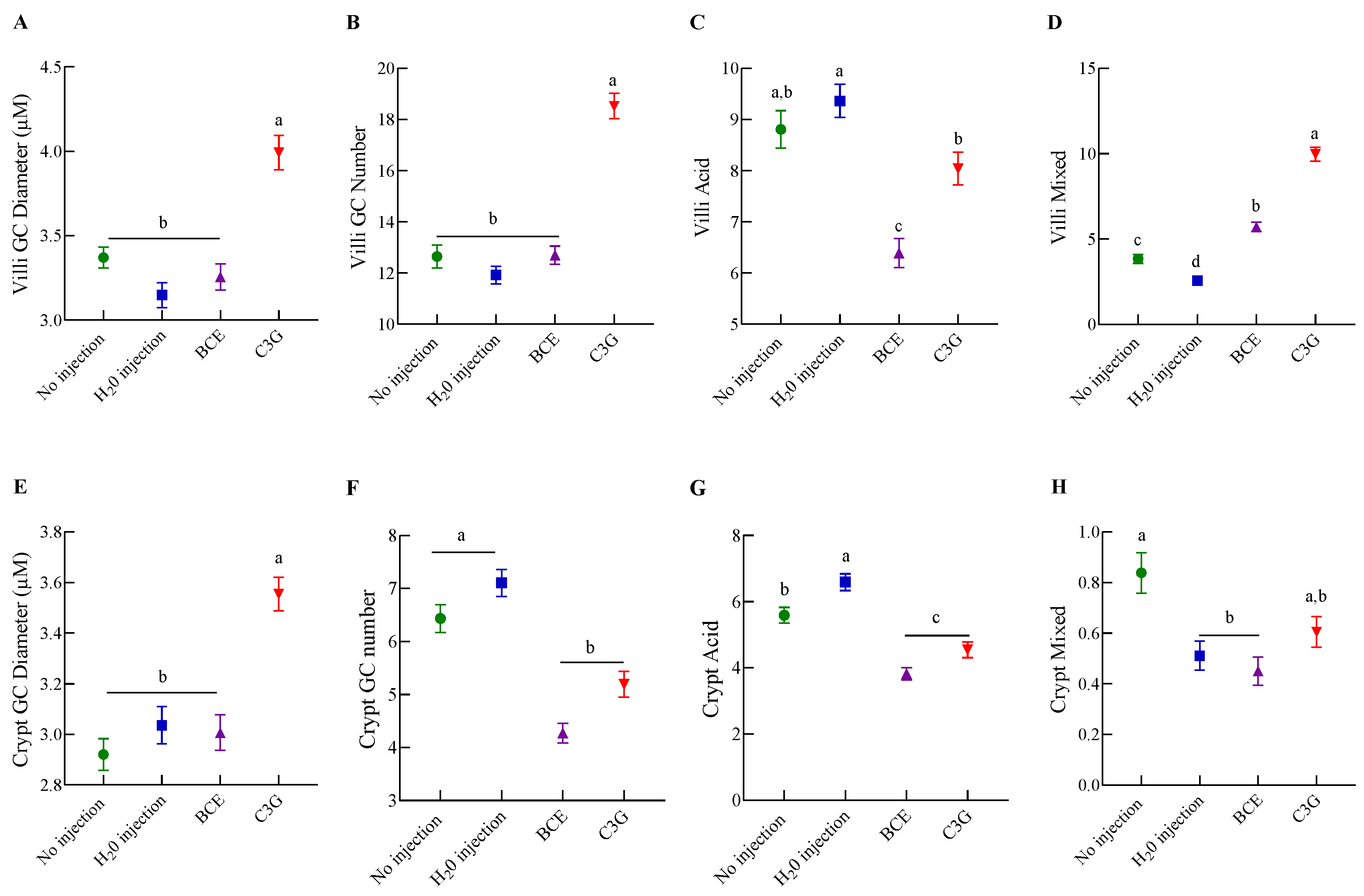
3.4. Effect of BCE on Duodenal Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salvador-Reyes, R.; Clerici, M.T.P.S. Peruvian Andean Maize: General Characteristics, Nutritional Properties, Bioactive Compounds, and Culinary Uses. Food Res. Int. 2020, 130, 108934. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Ferron, L.; Papetti, A. Colored Corn: An Up-Date on Metabolites Extraction, Health Implication, and Potential Use. Molecules 2021, 26, 199. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Pilu, R.; Tonelli, C. Anthocyanins in Corn: A Wealth of Genes for Human Health. Planta 2014, 240, 901–911. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, Y.; Jiang, L.; Zhou, W. Anthocyanins in Food. Encycl. Food Chem. 2019, 2, 10–17. [Google Scholar]
- Verediano, T.A.; Stampini Duarte Martino, H.; Dias Paes, M.C.; Tako, E. Effects of Anthocyanin on Intestinal Health: A Systematic Review. Nutrients 2021, 13, 1331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Luna-Vital, D.; de Mejia, E.G. Anthocyanins from Colored Maize Ameliorated the Inflammatory Paracrine Interplay between Macrophages and Adipocytes through Regulation of NF- ΚB and JNK-Dependent MAPK Pathways. J. Funct. Foods 2019, 54, 175–186. [Google Scholar] [CrossRef]
- Gowd, V.; Bao, T.; Wang, L.; Huang, Y.; Chen, S.; Zheng, X.; Cui, S.; Chen, W. Antioxidant and Antidiabetic Activity of Blackberry after Gastrointestinal Digestion and Human Gut Microbiota Fermentation. Food Chem. 2018, 269, 618–627. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X.; et al. Gut Microbiota Modulation and Anti-Inflammatory Properties of Anthocyanins from the Fruits of Lycium Ruthenicum Murray in Dextran Sodium Sulfate-Induced Colitis in Mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef]
- Wang, H.; Liu, D.; Ji, Y.; Liu, Y.; Xu, L.; Guo, Y. Dietary Supplementation of Black Rice Anthocyanin Extract Regulates Cholesterol Metabolism and Improves Gut Microbiota Dysbiosis in C57BL/6J Mice Fed a High-Fat and Cholesterol Diet. Mol. Nutr. Food Res. 2020, 64, e1900876. [Google Scholar] [CrossRef]
- Jurgoński, A.; Juśkiewicz, J.; Zduńczyk, Z. An Anthocyanin-Rich Extract from Kamchatka Honeysuckle Increases Enzymatic Activity within the Gut and Ameliorates Abnormal Lipid and Glucose Metabolism in Rats. Nutrition 2013, 29, 898–902. [Google Scholar] [CrossRef]
- Hribar, U.; Ulrih, P. The Metabolism of Anthocyanins. Curr. Drug Metab. 2014, 15, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.X.; Wu, S. The Effects and Mechanisms of Cyanidin-3-Glucoside and Its Phenolic Metabolites in Maintaining Intestinal Integrity. Antioxidants 2019, 8, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verediano, T.A.; Sant’ Ana, C.T.; Grancieri, M.; Parzanini Brilhante de São José, V.; Toledo, R.C.L.; Dias Paes, M.C.; Duarte Martino, H.S. Black Corn (Zea Mays L.) Whole Flour Improved the Antioxidant Capacity and Prevented Adipogenesis in Mice Fed a High-Fat Diet. Food Funct. 2022, 13, 5590–5601. [Google Scholar] [CrossRef]
- Rahman, S.; Mathew, S.; Nair, P.; Ramadan, W.S.; Vazhappilly, C.G. Health Benefits of Cyanidin-3-Glucoside as a Potent Modulator of Nrf2-Mediated Oxidative Stress. Inflammopharmacology 2021, 29, 907–923. [Google Scholar] [CrossRef]
- Domínguez-Avila, J.A.; Villa-Rodriguez, J.A.; Montiel-Herrera, M.; Pacheco-Ordaz, R.; Roopchand, D.E.; Venema, K.; González-Aguilar, G.A. Phenolic Compounds Promote Diversity of Gut Microbiota and Maintain Colonic Health. Dig. Dis. Sci. 2021, 66, 3270–3289. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The Intestinal Barrier: A Fundamental Role in Health and Disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of Anthocyanins and Consequent Effects on the Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2018, 59, 982–991. [Google Scholar] [CrossRef]
- Verediano, T.A.; Stampini Duarte Martino, H.; Kolba, N.; Fu, Y.; Cristina Dias Paes, M.; Tako, E. Black Corn (Zea Mays L.) Soluble Extract Showed Anti-Inflammatory Effects and Improved the Intestinal Barrier Integrity in Vivo (Gallus gallus). Food Res. Int. 2022, 157, 111227. [Google Scholar] [CrossRef]
- Hou, T.; Tako, E. The in Ovo Feeding Administration (Gallus gallus)—An Emerging in Vivo Approach to Assess Bioactive Compounds with Potential Nutritional Benefits. Nutrients 2018, 10, 418. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.M.E.; Shehata, A.M.; Alzahrani, S.O.; Shafi, M.E.; Mesalam, N.M.; Taha, A.E.; Swelum, A.A.; Arif, M.; Fayyaz, M.; Abd El-Hack, M.E. The Role of Polyphenols in Poultry Nutrition. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1851–1866. [Google Scholar] [CrossRef]
- Agarwal, N.; Kolba, N.; Jung, Y.; Cheng, J.; Tako, E. Saffron (Crocus Sativus L.) Flower Water Extract Disrupts the Cecal Microbiome, Brush Border Membrane Functionality, and Morphology In Vivo (Gallus gallus). Nutrients 2022, 14, 220. [Google Scholar] [CrossRef]
- Gomes, M.J.C.; Kolba, N.; Agarwal, N.; Kim, D.; Eshel, A.; Koren, O.; Tako, E. Modifications in the Intestinal Functionality, Morphology and Microbiome Following Intra-Amniotic Administration (Gallus gallus) of Grape (Vitis vinifera) Stilbenes (Resveratrol and Pterostilbene). Nutrients 2021, 13, 3247. [Google Scholar] [CrossRef]
- Gomes, M.J.C.; Martino, H.S.D.; Kolba, N.; Cheng, J.; Agarwal, N.; de Moura Rocha, M.; Tako, E. Zinc Biofortified Cowpea (Vigna Unguiculata L. Walp.) Soluble Extracts Modulate Assessed Cecal Bacterial Populations and Gut Morphology In Vivo (Gallus gallus). Front. Biosci. 2022, 27, 140. [Google Scholar] [CrossRef]
- Ranilla, L.G.; Christopher, A.; Sarkar, D.; Shetty, K.; Chirinos, R.; Campos, D. Phenolic Composition and Evaluation of the Antimicrobial Activity of Free and Bound Phenolic Fractions from a Peruvian Purple Corn (Zea mays L.) Accession. J. Food Sci. 2017, 82, 2968–2976. [Google Scholar] [CrossRef]
- Martino, H.S.D.; Kolba, N.; Tako, E. Yacon (Smallanthus sonchifolius) Flour Soluble Extract Improve Intestinal Bacterial Populations, Brush Border Membrane Functionality and Morphology in Vivo (Gallus gallus). Food Res. Int. 2020, 137, 109705. [Google Scholar] [CrossRef]
- Lao, F.; Giusti, M.M. Extraction of Purple Corn (Zea Mays L.) Cob Pigments and Phenolic Compounds Using Food-Friendly Solvents. J. Cereal Sci. 2018, 80, 87–93. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A.J. Colorimetry of Total Phenolics with Phosphomolybdic Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Grancieri, M.; Martino, H.S.D.; de Mejia, E.G. Digested Total Protein and Protein Fractions from Chia Seed (Salvia Hispanica L.) Had High Scavenging Capacity and Inhibited 5-LOX, COX-1-2, and INOS Enzymes. Food Chem. 2019, 289, 204–214. [Google Scholar] [CrossRef]
- de Araujo Santiago, M.C.P.; Galhardo Borguini, R.; da Silva de Mattos do Nascimento, L.; de Oliveira Braga, E.C.; de Carvalho Martins, V.; Senna Gouvêa, A.C.M.; Marques Peixoto, F.; Pacheco, S.; Nogueira, R.I.; de Oliveira Godoy, R.L. Jabuticaba (Myrciaria Jaboticaba (Vell.) O. Berg) Peel Powder Produced by Convective Drying Process: A Rich Anthocyanin Product. Fruits 2018, 73, 201–208. [Google Scholar] [CrossRef]
- Dias, D.M.; Kolba, N.; Hart, J.J.; Ma, M.; Sha, S.T.; Lakshmanan, N.; Nutti, M.R.; Martino, H.S.D.; Glahn, R.P.; Tako, E. Soluble Extracts from Carioca Beans (Phaseolus Vulgaris L.) Affect the Gut Microbiota and Iron Related Brush Border Membrane Protein Expression in Vivo (Gallus gallus). Food Res. Int. 2019, 123, 172–180. [Google Scholar] [CrossRef]
- Tako, E.; Glahn, R.P.; Welch, R.M.; Lei, X.; Yasuda, K.; Miller, D.D. Dietary Inulin Affects the Expression of Intestinal Enterocyte Iron Transporters, Receptors and Storage Protein and Alters the Microbiota in the Pig Intestine. Br. J. Nutr. 2008, 99, 472–480. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Kolba, N.; Sisser, P.; Turjeman, S.; Even, C.; Koren, O.; Tako, E. Intraamniotic Administration (Gallus gallus) of Genistein Alters Mineral Transport, Intestinal Morphology, and Gut Microbiota. Nutrients 2022, 14, 3473. [Google Scholar] [CrossRef]
- Bhaswant, M.; Shafie, S.R.; Mathai, M.L.; Mouatt, P.; Brown, L. Anthocyanins in chokeberry and purple maize attenuate diet-induced metabolic syndrome in rats. Nutrition 2017, 41, 24–31. [Google Scholar] [CrossRef]
- Zhang, Q.; Mejia, E.G.; Luna-Vital, D.; Tao, T.; Chandrasekaran, S.; Chatham, L.; Juvik, J.; Singh, V.; Kumar, D. Relationship of phenolic composition of selected purple maize (Zea mays L.) genotypes with their anti-inflammatory, anti-adipogenic and anti-diabetic potential. Food Chem. 2019, 289, 739–750. [Google Scholar] [CrossRef]
- Cong-Cong, X.U.; Wang, B.; Yi-Qiong, P.U.; Jian-Sheng, T.; Tong, Z. Advances in Extraction and Analysis of Phenolic Compounds from Plant Materials. Chin. J. Nat. Med. 2017, 15, 721–731. [Google Scholar]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids. Hindawi Sci. World J. 2013, 2013, 533–548. [Google Scholar] [CrossRef]
- Hanske, L.; Engst, W.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Contribution of Gut Bacteria to the Metabolism of Cyanidin 3-Glucoside in Human Microbiota-Associated Rats. Br. J. Nutr. 2013, 109, 1433–1441. [Google Scholar] [CrossRef]
- Jayamanne, V.S.; Adams, M.R. Modelling the Effects of PH, Storage Temperature and Redox Potential (Eh) on the Survival of Bifidobacteria in Fermented Milk. Int. J. Food Sci. Technol. 2009, 44, 1131–1138. [Google Scholar] [CrossRef]
- Vendrame, S.; Klimis-Zacas, D. Anti-Inflammatory Effect of Anthocyanins via Modulation of Nuclear Factor- KB and Mitogen-Activated Protein Kinase Signaling Cascades. Nutr. Rev. 2015, 73, 348–358. [Google Scholar] [CrossRef]
- Pawłowska, B.; Sobieszczańska, B.M. Intestinal Epithelial Barrier: The Target for Pathogenic Escherichia Coli. Adv. Clin. Exp. Med. 2017, 26, 1437–1445. [Google Scholar] [CrossRef] [Green Version]
- Ajiboye, T.O.; Habibu, R.S.; Saidu, K.; Haliru, F.Z.; Ajiboye, H.O.; Aliyu, N.O.; Ibitoye, O.B.; Uwazie, J.N.; Muritala, H.F.; Bello, S.A.; et al. Involvement of Oxidative Stress in Protocatechuic Acid-Mediated Bacterial Lethality. Microbiologyopen 2017, 6, e00472. [Google Scholar] [CrossRef]
- Tarabova, L.; Makova, Z.; Piesova, E.; Szaboova, R.; Faixova, Z. Intestinal Mucus Layer and Mucins (A Review). Folia Vet. 2016, 60, 21–25. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, C.; Zhang, H.; Qu, G.; Li, C.; Liu, L. Biotransformation of Polyphenols in Apple Pomace Fermented by β-Glucosidase-Producing Lactobacillus Rhamnosus L08. Foods 2021, 10, 1343. [Google Scholar] [CrossRef]
- Markowiak, P.; Ślizewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Huang, B.; Wang, Z.; Park, J.H.; Ryu, O.H.; Choi, M.K.; Lee, J.Y.; Kang, Y.H.; Lim, S.S. Anti-Diabetic Effect of Purple Corn Extract on C57BL/KsJ Db/Db Mice. Nutr. Res. Pract. 2015, 9, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Witten, J.; Samad, T.; Ribbeck, K. Selective Permeability of Mucus Barriers. Curr. Opin. Biotechnol. 2018, 52, 124–133. [Google Scholar] [CrossRef]
- Fachi, J.L.; de Souza Felipe, J.; Pral, L.P.; da Silva, B.K.; Corrêa, R.O.; de Andrade, M.C.P.; da Fonseca, D.M.; Basso, P.J.; Câmara, N.O.S.; e Souza, É.L.d.S.; et al. Butyrate Protects Mice from Clostridium Difficile-Induced Colitis through an HIF-1-Dependent Mechanism. Cell Rep. 2019, 27, 750–761.e7. [Google Scholar] [CrossRef] [Green Version]
- Gassler, N. Paneth Cells in Intestinal Physiology and Pathophysiology. World J. Gastrointest. Pathophysiol. 2017, 8, 150–160. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Forward P. (5′-3′) | Reverse P. (5′-3′) | Base Pairs Length | GI Identifier |
|---|---|---|---|---|
| Inflammatory Response | ||||
| TNFα | GACAGCCTATGCCAACAAGTA | TTACAGGAAGGGCAACTCATC | 109 | 53,854,909 |
| NF-κB1 | CACAGCTGGAGGGAAGTAAAT | TTGAGTAAGGAAGTGAGGTTGAG | 100 | 2,130,627 |
| IL-1β | CTCACAGTCCTTCGACATCTTC | TGTTGAGCCTCACTTTCTGG | 119 | 88,702,685 |
| Intestinal Functionality | ||||
| MUC2 | CCTGCTGCAAGGAAGTAGAA | GGAAGATCAGAGTGGTGCATAG | 272 | 423,101 |
| OCLN | GTCTGTGGGTTCCTCATCGT | GTTCTTCACCCACTCCTCCA | 124 | 396,026 |
| AMPK | CTCCACTTCCAGAAGGTTACTT | GCAGTAGCTATCGTTCATCCTATC | 140 | 427,185 |
| CDX2 | ACCAGGACGAAGGACAAATAC | CTTTCCTCCGGATGGTGATATAG | 103 | 374,205 |
| VDAC2 | CAGCACTCGCTTTGGAATTG | GTGTAACCCACTCCAACTAGAC | 99 | 395,498 |
| 18S rRNA | GCAAGACGAACTAAAGCGAAAG | TCGGAACTACGACGGTATCT | 100 | 7,262,899 |
| Components | Amount | Retention Time (min) |
|---|---|---|
| Cyanidin-3-glucoside (mg/100 g) | 283.91 | 6.5 |
| Pelargonidin-3-O-glucoside (mg/100 g) | 39.57 | 8.7 |
| Total phenolic compounds (mg GAE/100 g sample) | 555.00 | - |
| DPPH (%) | 70.79 | - |
| No Injection | H2O Injection | BCE | C3G | |
|---|---|---|---|---|
| Villi height (μM) | 193.12 ± 3.75 b | 171.50 ± 4.01 c | 169.69 ± 2.10 c | 202.43 ± 2.81 a |
| Villi surface (μM2) | 12,324.31 ± 344.23 b | 11,740.52 ± 336.59 b | 11,181.40 ± 224.95 b | 15,250.89 ± 390.56 a |
| Depth crypt (μM) | 24.98 ± 1.01 b | 37.30 ± 1.08 a | 24.78 ± 0.62 b | 35.35 ± 0.99 a |
| Paneth cell number | 0.95 ± 0.07 c | 1.30 ± 0.04 b | 1.27 ± 0.04 b | 2.49 ± 0.14 a |
| Paneth cell diameter | 1.69 ± 0.03 a | 1.55 ± 0.02 b | 1.50 ± 0.02 b | 1.58 ± 0.02 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrizzi Verediano, T.; Agarwal, N.; Stampini Duarte Martino, H.; Kolba, N.; Grancieri, M.; Dias Paes, M.C.; Tako, E. Effect of Black Corn Anthocyanin-Rich Extract (Zea mays L.) on Cecal Microbial Populations In Vivo (Gallus gallus). Nutrients 2022, 14, 4679. https://doi.org/10.3390/nu14214679
Agrizzi Verediano T, Agarwal N, Stampini Duarte Martino H, Kolba N, Grancieri M, Dias Paes MC, Tako E. Effect of Black Corn Anthocyanin-Rich Extract (Zea mays L.) on Cecal Microbial Populations In Vivo (Gallus gallus). Nutrients. 2022; 14(21):4679. https://doi.org/10.3390/nu14214679
Chicago/Turabian StyleAgrizzi Verediano, Thaisa, Nikita Agarwal, Hércia Stampini Duarte Martino, Nikolai Kolba, Mariana Grancieri, Maria Cristina Dias Paes, and Elad Tako. 2022. "Effect of Black Corn Anthocyanin-Rich Extract (Zea mays L.) on Cecal Microbial Populations In Vivo (Gallus gallus)" Nutrients 14, no. 21: 4679. https://doi.org/10.3390/nu14214679