1. Introduction
Aging is a heterogeneous and complex process as many transformations happen to human organisms, such as a general decline in physiological function, increasing chronic low-grade inflammatory status, and increased risks of aging-related diseases. However, as an accepted model of successful aging, centenarians avoid or delay major age-related diseases, such as diabetes mellitus, Alzheimer disease, cardiovascular disease, and cancer [
1]. Therefore, understanding the mechanism of exceptional longevity is of important referential significance for allowing populations to live longer, disease-free, and have a good quality of life. Decades of study on aging have found many genes and biological processes associated with the aging process [
2], but its exact biological mechanism is still unclear. In particular, a general molecular profile that encompasses the healthy aging process as a result of multifactorial interactions is limited, and the data for centenarians are especially lacking.
Today, metabolomic approaches have become extremely promising tools for capturing overall metabolic changes associated with the normal aging process. Jové et al. found monoacylglyceride (22:1), diacylglyceride (33:2), resolvin D6, and phosphoserine (40:5) decreasing with the aging process by analyzing the metabolic profiles of healthy humans ranging from 30 to 100 years of age [
3]. Chak et al. identified significant ageing-associated metabolites that are involved in several ageing processes, such as oxidative stress resistance, autophagy, inflammation, lipid metabolism, and apoptosis [
4]. Bunning et al. applied random forest models to explore biological processes of aging in a cross-sectional cohort of healthy individuals aged 6 months to 82 years, which highlighted established metabolites, such as amino acids, steroids, and free fatty acids [
5]. The above reports focused primarily on the normal aging process itself rather than the extreme longevity. The relevant reports on a comprehensive metabolic phenotype of centenarians are still relatively limited. Collino and Montoliu et al. performed metabolic profiling of Italian centenarians using NMR metabonomics and targeted analysis approaches and found that the centenarians possessed a unique eicosanoid metabolism network and identified phospho- and sphingolipids as markers of human longevity [
1,
6]. Nevertheless, as an important factor affecting metabolic profiles, the dietary influence has not yet been considered in both the researches. Furthermore, the exploration of integrative metabolic pathways closely related to extreme longevity in humans is still in its infancy so far, which, however, could provide insights into underlying molecular mechanisms and biological processes of healthy aging.
There is a remarkable phenomenon in Bama County, Guangxi Province, China. Based on the Population Census of China in 2020, there were 102 centenarians in the population of 236,152 in Bama County, a ratio of 43 centenarians per 1 × 10
5 persons [
7]. To date, the ratio of centenarians in this region is the highest in China, and is far above the world longevity county standard defined by the United Nations (7.5/100,000) [
8]. The rare and amazing phenomenon of extreme longevity and healthy aging possesses distinctive local features, and therefore, the centenarians from this longevous region provide a valuable paradigm of healthy and successful aging for people to follow and imitate. Accordingly, we have reported the characteristics of nutrient intakes, specific metabolites and elements in nails of elderly people from the longevous region compared with a non-longevous region in previous studies [
9,
10]. However, no work has focused on the comprehensive metabolic profiles of centenarians living in Bama longevous region as a result of their traditional and conservative lifestyle.
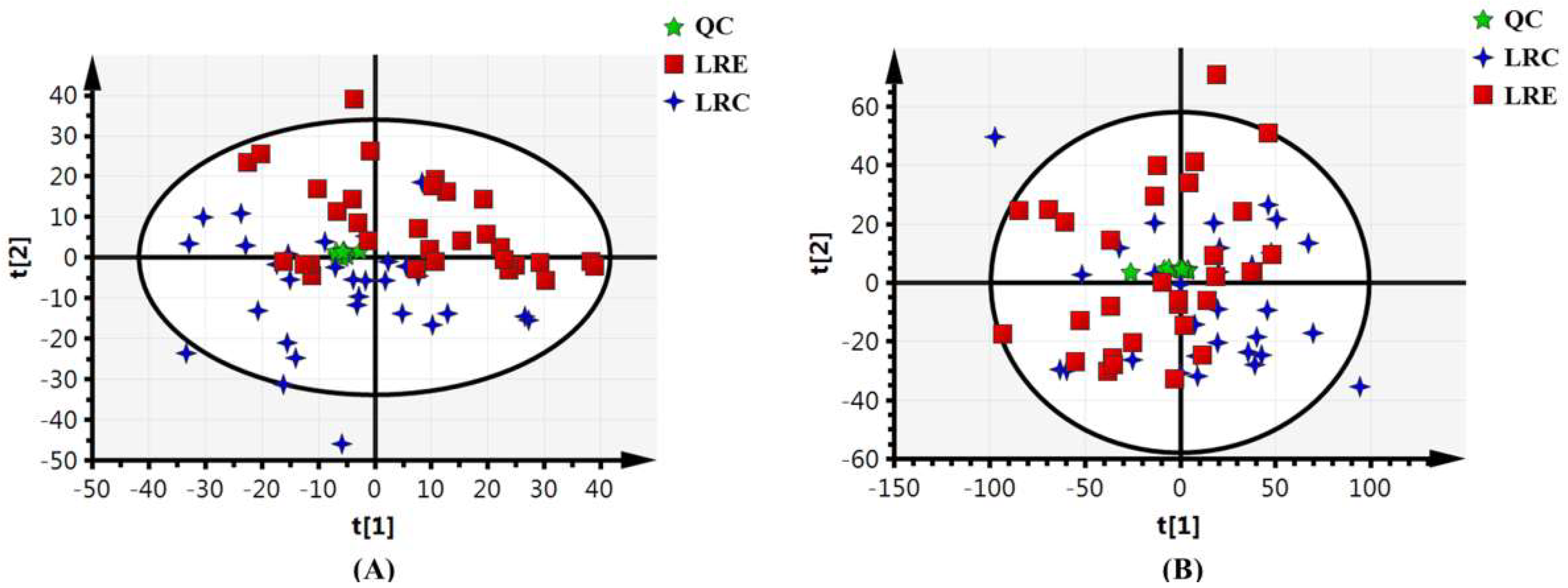
Since our centenarians well represent a model of healthy and successful aging, the primary objective of this study is to discover the metabolic pattern of healthy aging by portraying the integrative metabolic profiles and then further capturing the metabolic signatures of the extreme longevity using the non-targeted metabolomics approach combined with metabolic target analysis. To achieve the aim, the healthy centenarians in the longevous region (LRC group) and the elderly people aged 60–70 in the longevous region (LRE group) were enrolled according to strict screening criteria. The characteristic metabolites and relevant metabolic pathways were identified based on the orthogonal projections to latent structures discriminant analysis (OPLS-DA) model. Moreover, the SCFA metabolism was analyzed. This will be very important for a better understanding and pursuit of longevity from the perspective of comprehensive metabolic profiles.
4. Discussion
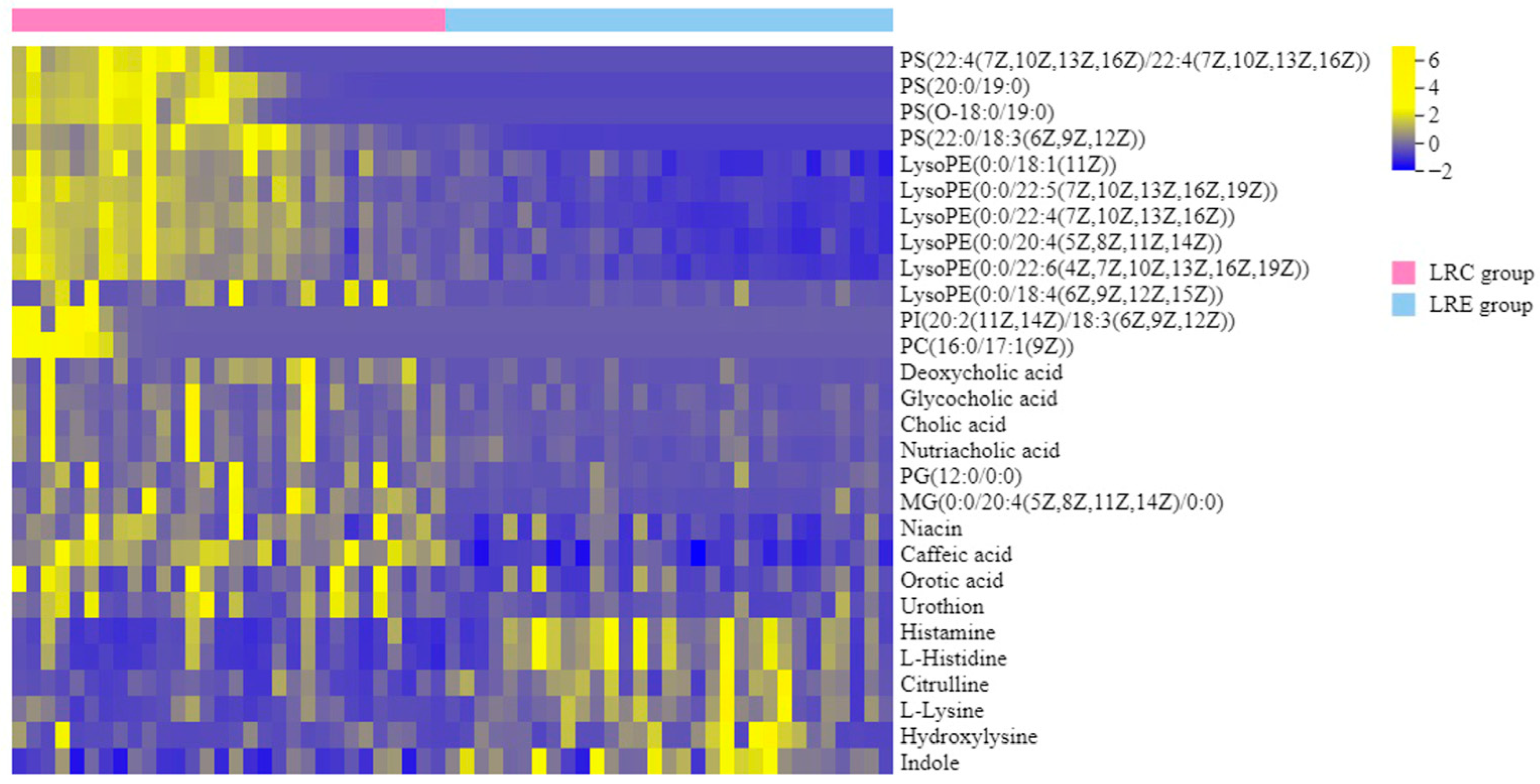
Through the systematic analysis of comprehensive metabolic profiles of the healthy centenarians by the non-targeted metabolomics approaches coupled with the metabolic target analysis, we captured the unique metabolic signatures of the exceptional longevity, which achieved initial aim of this study and expanded our previous investigation on longevity characterization [
10]. As centenarians provide an excellent paradigm of healthy aging, the exploration of metabolic patterns of healthy centenarians from the longevous region opens a window into extreme longevity. The relevant studies have elucidated some important metabolic alterations related to the aging process [
1,
3,
4,
5,
6,
14,
15]. However, most of the reports lacked the guidance of a successful model of healthy aging. Only Collino and Montoliu et al. described the metabolic phenotype of Italian centenarians [
1,
6]. Despite these findings, the underlying metabolic pathways associated with these phenotypes as well as the dietary influence on the metabolism of centenarians have remained poorly understood. Nevertheless, we discovered the 28 characteristic metabolites and 6 metabolic pathways closely related to the centenarians for the first time, reflecting distinctive remodeling of phospholipid and amino acid metabolism. Moreover, we also demonstrated diet-associated remodeling of SCFA metabolism. These specific metabolic remodeling may play a critical role in shaping healthy aging.
It has been reported that phospholipids metabolism changed with aging, whereas there were some differences in individual phospholipid molecules among different studies [
1,
6,
14,
15,
16], which could be attributed to the differences in geographic areas, genetic background, race, living environment, lifestyle, and dietary habits of the subjects. PC (14:0/18:1), PC (16:0/18:1), PC (16:0/18:2), PC (14:0/18:2), PC (16:0/18:3), PC (18:0/22:5), PE (16:0/20:4), PE (18:0/20:2), PE (18:0/20:3), PE (18:0/20:4), PI (18:0/18:1), PI (18:1/16:0), PI (20:3/18:0) were increased in Italian centenarians [
6]. However, we found that the characteristic metabolites PS (O-18:0/19:0), PS (22:4/22:4), PS (20:0/19:0) and PS (22:0/18:3) were significantly increased in the LRC group (
p < 0.05), especially PS (O-18:0/19:0), PS (22:4/22:4) and PS (20:0/19:0) with the highest FC. PSs have some potential cognitive benefits, which have been shown to increase memory performance in the elderly [
17,
18]. PSs are also widely involved in some important physiological processes, including phagocytosis by macrophage, and activation of protein kinase C [
19]. This indicates that given the cognitive benefits, higher PSs should be beneficial to health and longevity, which also reflects the specific remodeling of phospholipids metabolism of the healthy centenarians from the longevous region.
The four bile acids, including cholic acid, deoxycholic acid, glycocholic acid, and nutriacholic acid, were significantly increased in the LRC group (
p < 0.05), which were also characteristic metabolites closely related to the centenarians. Studies have demonstrated that bile acids act as metabolic regulators and nutrient sensors to regulate glucose and lipid metabolism, and immune response [
20]. As signaling molecules, in mammals, bile acids specifically bind to and activate some receptors, hence stimulating many vital longevity-promoting and healthspan-promoting processes, such as anti-inflammatory processes [
21]. Therefore, our finding suggests that the above four up-regulated bile acids may be beneficial to healthy aging. Notably, Zhang et al. found that the level of glycocholic acid was significantly decreased with age ranging from 20 to 74 years old [
22]. In turn to see the discovery of this study, it is reasonably speculated that the bile acids metabolism of the healthy centenarians may revert to a more youthful state with respect to the elderly people aged 60–70 years, which also represents the unique metabolic signatures of the healthy centenarians from the longevous region, and a path to health and longevity by means of appropriately up-regulated bile acids metabolism.
The metabolic fate of the characteristic metabolite L-histidine has three possible routes. The first pathway is its conversion to the biogenic amine histamine. As a neurotransmitter, histamine is also involved in local immune responses [
14]. The second pathway is its metabolism to urocanic acid. The third metabolic pathway that consumes L-histidine produces carnosine that is a dipeptide from β-alanine and L-histidine. Owing to its antioxidant characteristics, carnosine is considered to be a natural anti-aging substance capable of suppressing oxidative damage, glycation of proteins, and scavenging toxic age-related molecules [
23]. From this point of view, the lower L-histidine levels in our study (
p < 0.05) owing to its consumption by carnosine biosynthesis with advancing age might to some extent reflect a response to oxidative stress. A previous study showed that histidine decreased in serum with age (participants aged 32–81) [
14]. However, we found that L-histidine and histamine significantly decreased in the urine of healthy centenarians (
p < 0.05), displaying the unique and effective remodeling of histidine metabolism in centenarians to counteract oxidative stress.
We also discovered that the characteristic metabolite citrulline was significantly decreased in the LRC group (
p < 0.05), which was involved in urea cycle that was down-regulated as well. In urea cycle, arginine level was also lower in the LRC group (
p < 0.05). It has been shown that arginine was significantly decreased in ageing participants of KORA and CARLA [
4]. Moreover, arginine level was recently shown to be significantly positively correlated with dietary carbohydrate intake (
R = 0.79,
p < 0.05) [
24]. As for this study, we found that the carbohydrate intake was also significantly lower in the LRC group (
p < 0.05), with a similar trend with that in the above report. Intriguingly, aspartate was significantly increased in the LRC group (
p < 0.05). The relevant studies in cells and mammals have shown that aspartate supplementation reduces ROS production in neuroblastoma cells and reduces oxidative stress and increases antioxidant levels in the blood [
25,
26]. In fact, decreased ROS is believed to be a critical mechanism behind the extended lifespan and health span [
27]. However, we have found no available data on relationships between aspartate and human longevity so far. Based on the discovery of this study, it is reasonable to assume that aspartate metabolism plays an important and positive role in human longevity.
As a nitrogen shuttle, glutamine takes up excess ammonia and forms urea, thereby reducing toxic build-up in the brain and improving brain functions [
28]. Glutamine also plays an important role in NF-κB signal transduction pathways, contributing to the attenuation of local inflammation [
6]. Montoliu et al. found that glutamine was increased in serum of Italian centenarians by
1H-NMR [
6]. However, interestingly, we discovered a significant increase in glutamine in the urine of healthy centenarians from Bama County (
p < 0.05). Given the physiological functions of glutamine metabolism, higher glutamine should have a positive influence on the health and longevity of centenarians.
In the pathway of ascorbate and aldarate metabolism, we observed a significant increased level of myo-inositol in the LRC group (
p < 0.05). Myo-inositol promoted healthspan and prevented age-related decline in physiological functions in worm and mouse [
29,
30]. However, the relevant studies in humans have not been reported. Our finding suggests that increased myo-inositol may be conducive to longevity of centenarians.
As to tryptophan metabolism, a previous study showed that the tryptophan and indole concentrations in feces progressively decreased with age (volunteers aged 2 to 85 years) [
31]. It has also been reported that tryptophan decreased in serum with increasing age [
1]. Nevertheless, we found that tryptophan and its degradation product indole in urine of healthy centenarians were significantly down-regulated (
p < 0.05) for the first time, indicating the characteristic remodeling of tryptophan metabolism of healthy centenarians from the longevous region. Additionally, their energy (
p < 0.01) and carbohydrate (
p < 0.05) intakes were significantly lower. Other work showed that tryptophan decreased with caloric restriction in an intervention study including eight subjects [
32]. It was also reported that very low-carbohydrate ketogenic diet also decreased tryptophan levels in mice [
33]. The above intervention study and mice experiment have similar trends with our results, suggesting that the specific remodeling of tryptophan metabolism may be associated with diet of centenarians.
We also discovered a particular remodeling of SCFA metabolism of healthy centenarians associated with dietary fiber intake (
p < 0.01). It has been shown that oldest-old adults had greater potential for SCFA production [
34]. Our previous study also found that SCFAs were increased in feces of the elderly people in Bama County, compared with the elderly people from a non-longevous region [
10]. SCFAs perform various physiological functions in the gut, including anti-inflammatory, antimicrobial, and antitumorigenic effects [
35], associated with lower risks for some diseases [
36], especially butyric acid and propionic acid, promoting metabolic benefits via gut-brain neural circuits [
37]. It is therefore concluded that the relatively higher levels of SCFAs may be conducive to longevity of the centenarians, and appropriate increased dietary fibers in daily diets should be a path toward the longevity.
This study has several strengths. First, the two complementary strategies, non-targeted metabolomics and metabolite target analysis were used. Non-targeted metabolomics is a valuable approach to obtaining a comprehensive depiction of the metabolic status closely related to the phenotypic outcome of interest in an unbiased manner [
38]. Meanwhile, targeted metabolomics, which is an accurate quantitative method to analyze biochemically known and annotated metabolites, provides information that is more precise on specific metabolites and metabolic pathways [
39]. Second, the two kinds of different biological samples, including urine and feces, were analyzed to comprehensively assess the metabolic features of healthy centenarians from different perspectives. To date, most of the relevant reports primarily focus on metabolites in serum and plasma [
3,
4,
5,
6,
14,
15,
16,
40]. Collino et al. explored the metabolic changes in serum and urine of Italian centenarians [
1]. However, the investigation on the signatures of global metabolic profiles of the exceptional longevity using two kinds of biological samples—urine and feces—by means of the two complementary metabolomics approaches has not been reported yet so far. Third, the four-season consecutive 7-day WDR method was used to assess the usual habitual nutrient intakes, which reflects real-life nutrient intakes and minimizes variances in dietary intakes according to seasons and days. Among available nutrition assessment approaches, the WDR method is the most accurate and robust and is accepted as a gold standard [
41], though it is expensive, time-consuming, and generally requires considerable commitment on the part of volunteers [
42]. Fourth, characteristic metabolites and metabolic pathways closely related to centenarians from the longevous region were discovered based on the OPLS-DA model that is reliable and robust for a small sample size.
Nevertheless, some limitations of this exploratory study need to be noted. First, healthy aging also depends on many other factors, such as hereditary and environmental factors [
43], while this study is limited by its cross-sectional design, and therefore a causal relationship between metabolism and longevity cannot be concluded directly. However, the unique metabolic signatures of the centenarians from the longevous region discovered in this study provided new clues for further exploration of the relationship between metabolism and longevity. Future studies are needed to investigate the metabolic mechanisms affecting healthy aging. Second, the detailed data on the smoking history and amount were unavailable, and therefore, it is necessary to exclude the influence of smoking in future work. Third, the strict screening criteria of participants, as well as an extremely limited number of healthy centenarians, resulted in a relatively small sample size. In addition, due to the tedious process of the 28-day WDR method and the complicated protocols of the sample collection, many volunteers withdrew from the study. Nevertheless, we applied the OPLS-DA method to construct the classification model of the LRC group and LRE group and identify characteristic metabolites. As a pattern recognition approach, OPLS-DA has distinct advantages in solving the classification problem of a very small number of samples [
44], which overcomes the limitation of sample size. Further studies including additional cohorts from different longevous regions are required to validate these findings.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}