
Combination Treatment with Hydroxytyrosol and Vitamin E Improves NAFLD-Related Fibrosis
 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Treatments, and Cell Viability
2.2. Cell Proliferation Assay
2.3. Real-Time Monitoring of Cell Proliferation
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. Immunofluorescence (IF)
2.6. Migration Assay
2.7. Collagen Lattice Contraction Assay
2.8. Intracellular Reactive Oxygen Species (ROS)
2.9. Western Blot (WB)
2.10. Pilot Animal Study
2.11. Study Patients
2.12. Enzyme-Linked ImmunoSorbent Assay (ELISA)
2.13. Statistical Analysis
3. Results
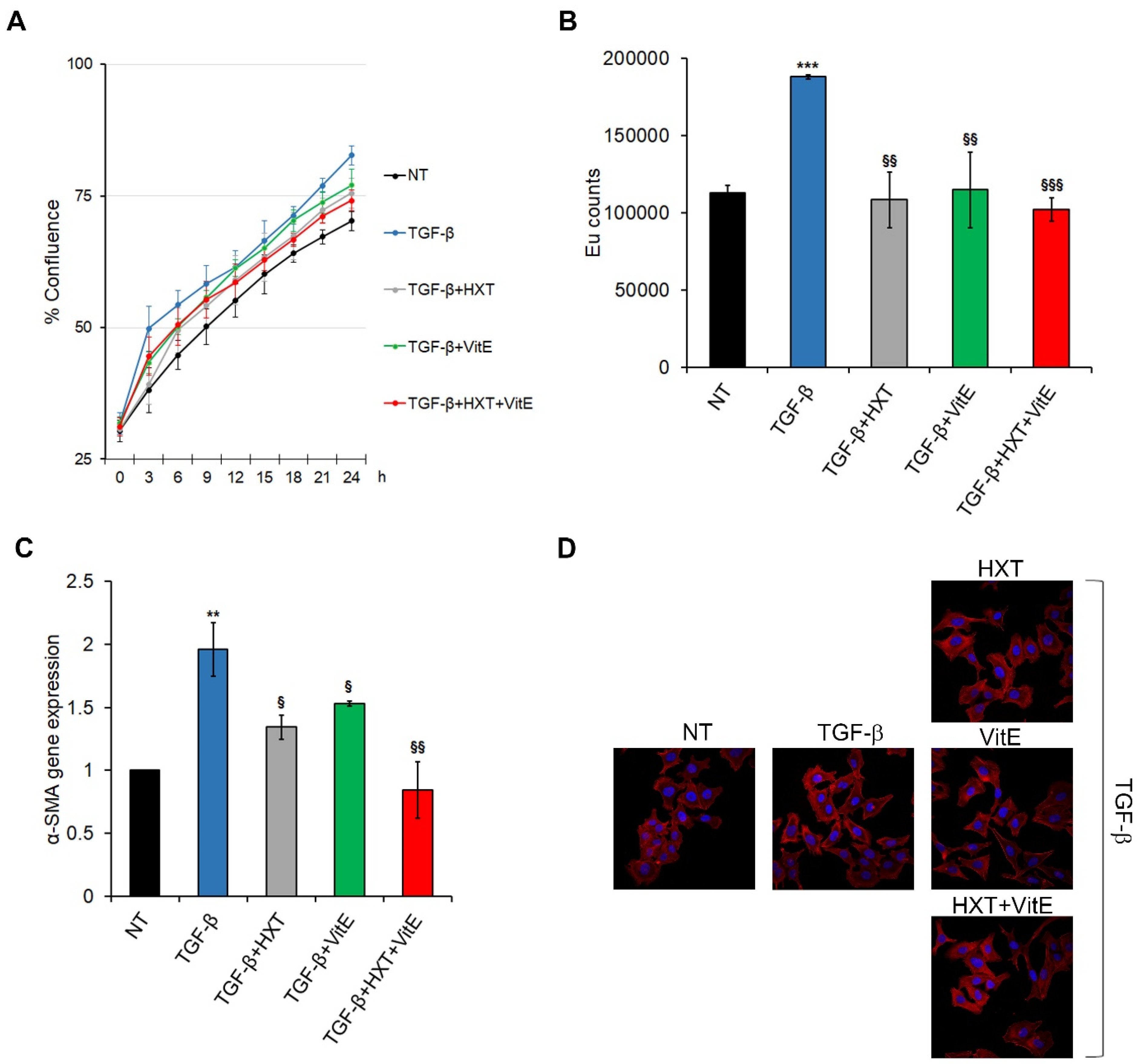
3.1. The Combination Treatment with HXT and VitE Reduces TGF-β-Induced HSC Activation
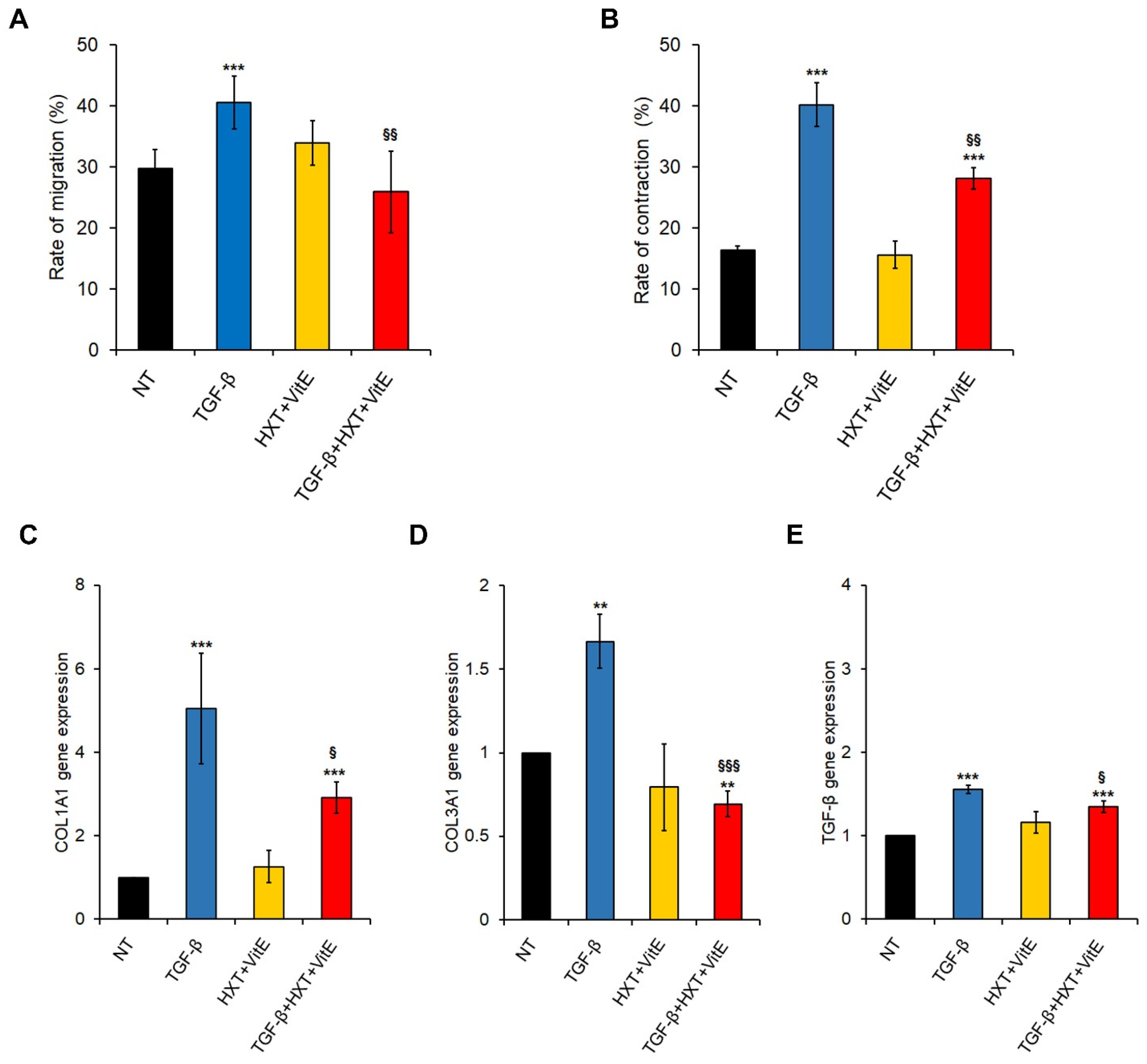
3.2. The Combination Treatment with HXT and VitE Inhibits TGF-β-Induced HSC Migration, Contractility and Pro-Fibrogenic Phenotype
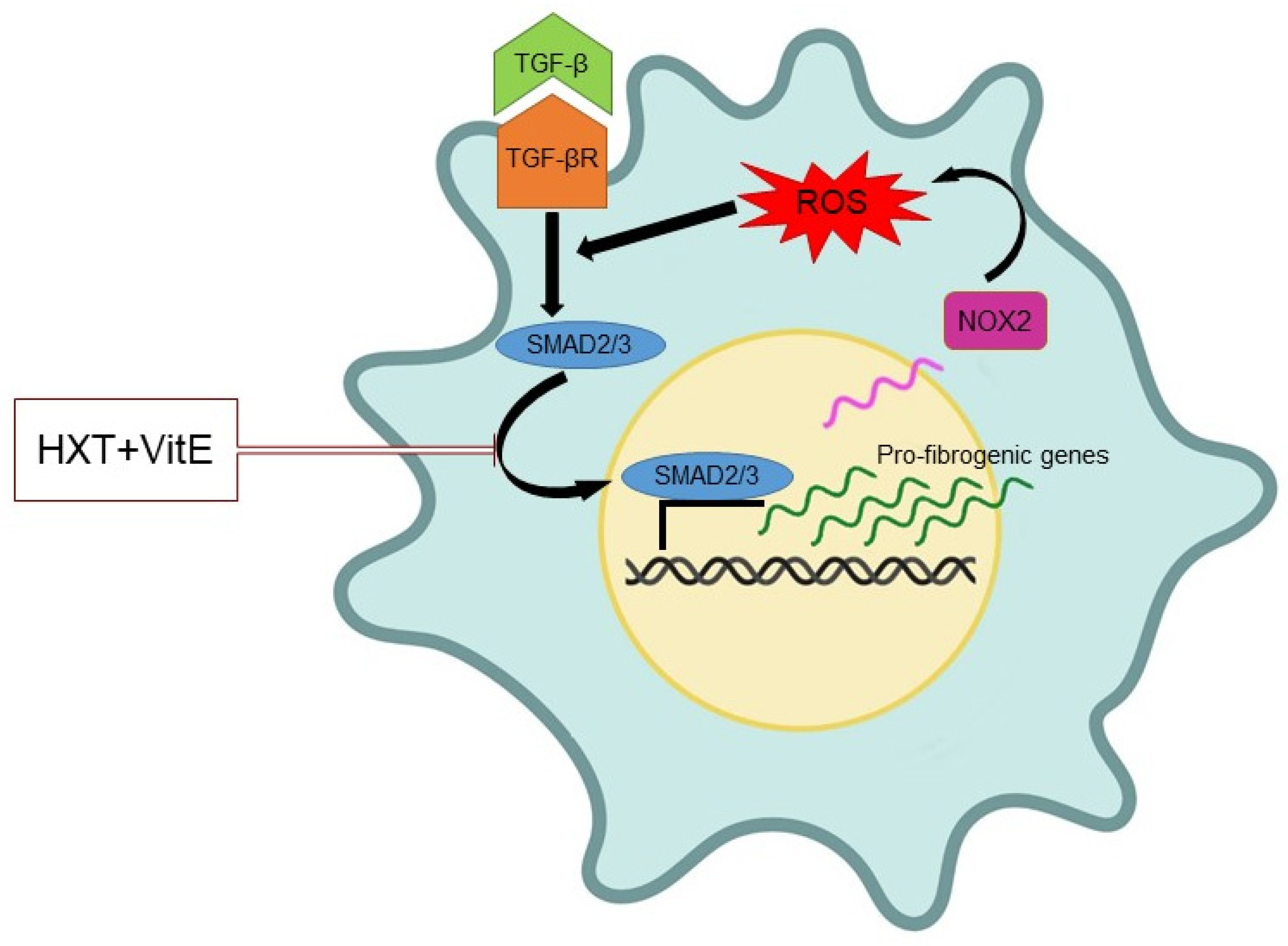
3.3. The Combination Treatment with HXT and VitE Inhibits TGF-β-Induced HSC Oxidative Stress by Attenuating the TGF-β/SMAD Signaling Pathway
3.4. HXT + VitE Reduces CCl4 plus WD-Induced Liver Fibrosis in Mice
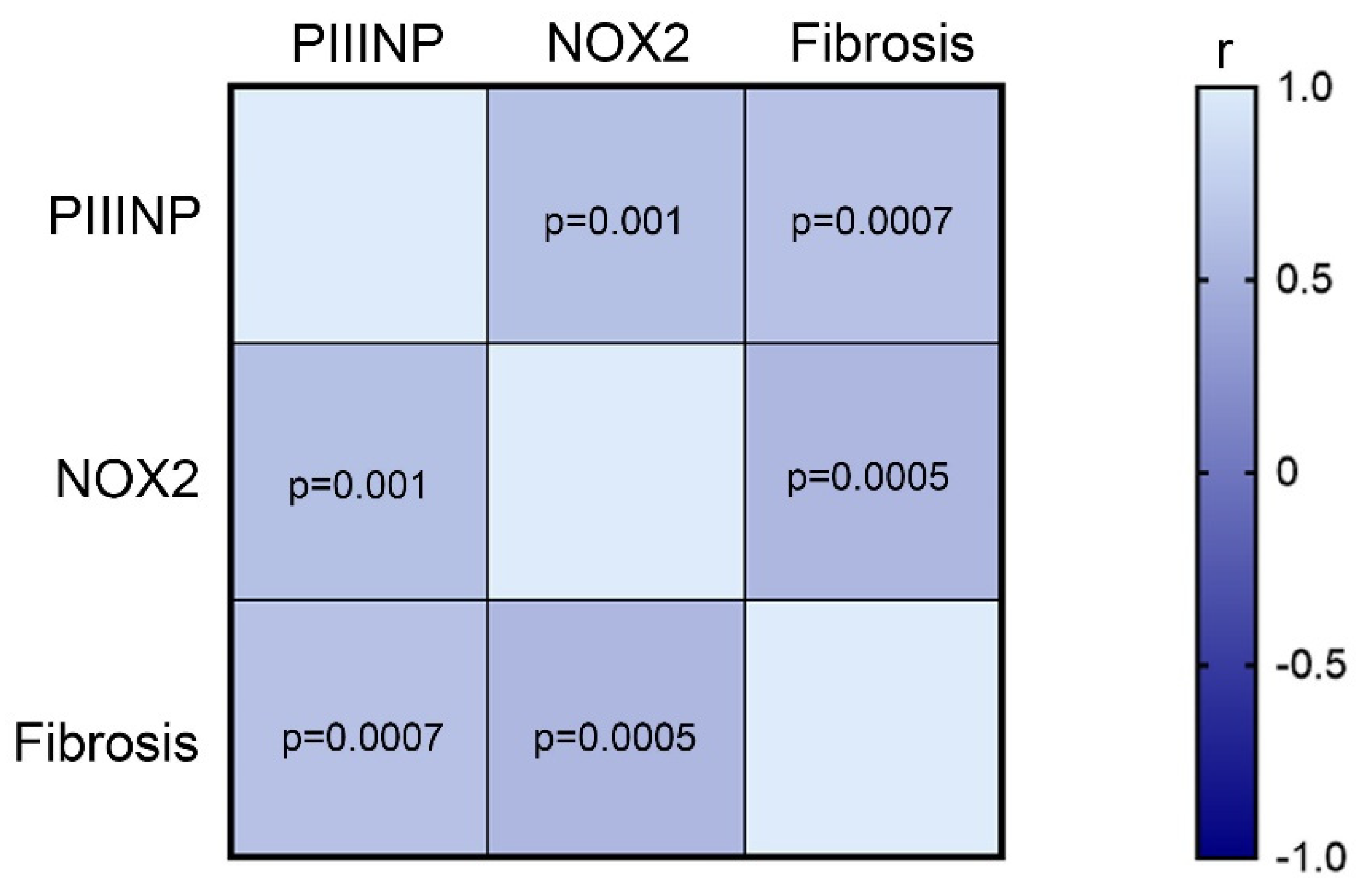
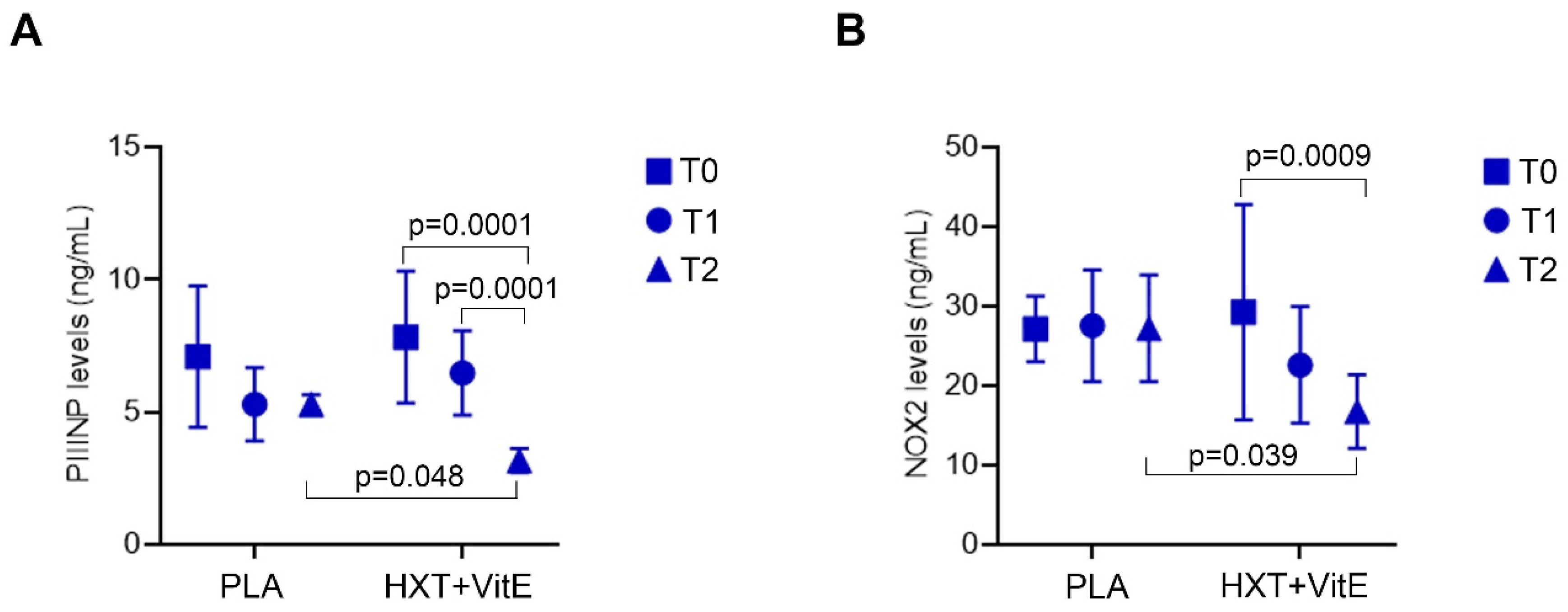
3.5. HXT + VitE Treatment Reduces PIIINP and NOX2 Levels in Children with NAFLD
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paik, J.M.; Golabi, P.; Younossi, Y.; Mishra, A.; Younossi, Z.M. Changes in the Global Burden of Chronic Liver Diseases From 2012 to 2017: The Growing Impact of NAFLD. Hepatology 2020, 72, 1605–1616. [Google Scholar] [CrossRef] [PubMed]
- Nobili, V.; Alisi, A.; Valenti, L.; Miele, L.; Feldstein, A.E.; Alkhouri, N. NAFLD in children: New genes, new diagnostic mo-dalities and new drugs. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Goldner, D.; Lavine, J.E. Nonalcoholic Fatty Liver Disease in Children: Unique Considerations and Challenges. Gastroenterology 2020, 158, 1967–1983.e1. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M.; Kleiner, D.E.; Carpenter, D.H.; Rinella, M.; Harrison, S.A.; Loomba, R.; Younossi, Z.; Neuschwander-Tetri, B.A.; Sanyal, A.J.; American Association for the Study of Liver Diseases NASH Task Force. NAFLD: Reporting Histologic Findings in Clinical Practice. Hepatology 2021, 73, 2028–2038. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.L.; Howe, L.; Jones, H.; Higgins, J.; Lawlor, D.A.; Fraser, A. The Prevalence of Non-Alcoholic Fatty Liver Disease in Children and Adolescents: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0140908. [Google Scholar] [CrossRef]
- Yu, E.L.; Schwimmer, J.B. Epidemiology of Pediatric Nonalcoholic Fatty Liver Disease. Clin. Liver Dis. 2021, 17, 196–199. [Google Scholar] [CrossRef]
- Cananzi, M.; Vajro, P.; Rela, M.; Dhawan, A. NAFLD and Liver Transplantation in Children—Working Group Report from the ILTS Single Topic Conference on NAFLD. Transplantation 2019, 103, 68–70. [Google Scholar] [CrossRef]
- Doycheva, I.; Issa, D.; Watt, K.D.; Lopez, R.; Rifai, G.; Alkhouri, N. Nonalcoholic Steatohepatitis is the Most Rapidly Increasing Indication for Liver Transplantation in Young Adults in the United States. J. Clin. Gastroenterol. 2018, 52, 339–346. [Google Scholar] [CrossRef]
- Thompson, M.D. Developmental Programming of NAFLD by Parental Obesity. Hepatol. Commun. 2020, 4, 1392–1403. [Google Scholar] [CrossRef]
- Abeysekera, K.W.; Orr, J.G.; Madley-Dowd, P.; Fernandes, G.S.; Zuccolo, L.; Gordon, F.H.; Lawlor, D.A.; Heron, J.; Hickman, M. Association of Maternal Pre-Pregnancy BMI And Breastfeeding with NAFLD in Young Adults: A Parental Negative Control Study. Lancet Reg. Health Eur. 2021, 10, 100206. [Google Scholar] [CrossRef]
- Goyal, N.P.; Rosenthal, S.B.; Nasamran, C.; Behling, C.A.; Angeles, J.E.; Fishbein, M.H.; Harlow, K.E.; Jain, A.K.; Mollewston, J.P.; Newton, K.P.; et al. Nonalcoholic Fatty Liver Disease Risk and Histologic Severity Are Associated with Genetic Polymorphisms in Children. Hepatology 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Mandala, A.; Janssen, R.C.; Palle, S.; Short, K.R.; Friedman, J.E. Pediatric Non-Alcoholic Fatty Liver Disease: Nutritional Origins and Potential Molecular Mechanisms. Nutrients 2020, 12, 3166. [Google Scholar] [CrossRef] [PubMed]
- Panera, N.; Barbaro, B.; Della Corte, C.; Mosca, A.; Nobili, V.; Alisi, A. A Review of the Pathogenic and Therapeutic Role of Nutrition in Pediatric Nonalcoholic Fatty Liver Disease. Nutr. Res. 2018, 58, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Katsagoni, C.N.; Papachristou, E.; Sidossis, A.; Sidossis, L. Effects of Dietary and Lifestyle Interventions on Liver, Clinical and Metabolic Parameters in Children and Adolescents with Non-Alcoholic Fatty Liver Disease: A Systematic Review. Nutrients 2020, 12, 2864. [Google Scholar] [CrossRef]
- Bayram, H.M.; Majoo, F.M.; Ozturkcan, A. Polyphenols in the Prevention and Treatment of Non-Alcoholic Fatty Liver Disease: An Update of Preclinical and Clinical Studies. Clin. Nutr. ESPEN 2021, 44, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Soto-Alarcon, S.A.; Valenzuela, R.; Valenzuela, A.; Videla, L.A. Liver Protective Effects of Extra Virgin Olive Oil: Interaction between Its Chemical Composition and the Cell-signaling Pathways Involved in Protection. Endocr. Metab. Immune Disord. Drug Targets 2017, 18, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, R.; Videla, L.A. Impact of the Co-Administration of N-3 Fatty Acids and Olive Oil Components in Preclinical Nonalcoholic Fatty Liver Disease Models: A Mechanistic View. Nutrients 2020, 12, 499. [Google Scholar] [CrossRef]
- Nobili, V.; Alisi, A.; Mosca, A.; Crudele, A.; Zaffina, S.; Denaro, M.; Smeriglio, A.; Trombetta, D. The Antioxidant Effects of Hydroxytyrosol and Vitamin E on Pediatric Nonalcoholic Fatty Liver Disease, in a Clinical Trial: A New Treatment? Antioxid. Redox Signal. 2019, 31, 127–133. [Google Scholar] [CrossRef]
- Mosca, A.; Crudele, A.; Smeriglio, A.; Braghini, M.R.; Panera, N.; Comparcola, D.; Alterio, A.; Sartorelli, M.R.; Tozzi, G.; Raponi, M.; et al. Antioxidant Activity of Hydroxytyrosol and Vitamin E Reduces Systemic Inflammation in Children with Paediatric NAFLD. Dig. Liver Dis. 2020, 53, 1154–1158. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Panera, N.; Mina, M.; Gnani, D.; De Stefanis, C.; Crudele, A.; Rychlicki, C.; Petrini, S.; Bruscalupi, G.; Agostinelli, L.; et al. LPS-Induced TNF-α Factor Mediates Pro-Inflammatory and Pro-Fibrogenic Pattern in Non-Alcoholic Fatty Liver Disease. Oncotarget 2015, 6, 41434–41452. [Google Scholar] [CrossRef] [Green Version]
- Condorelli, A.; Logli, E.; Cianfarani, F.; Teson, M.; Diociaiuti, A.; El Hachem, M.; Zambruno, G.; Castiglia, D.; Odorisio, T. MicroRNA-145-5p Regulates Fibrotic Features of Recessive Dystrophic Epidermolysis Bullosa Skin Fibroblasts. Br. J. Dermatol. 2019, 181, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of Hepatic Stellate Cell Activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Novo, E.; Parola, M. Redox Mechanisms in Hepatic Chronic Wound Healing and Fibrogenesis. Fibrogenesis Tissue Repair 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.-M.; Desai, L.P. Reciprocal Regulation Of TGF-Β And Reactive Oxygen Species: A Perverse Cycle for Fibrosis. Redox Biol. 2015, 6, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Dewidar, B.; Meyer, C.; Dooley, S.; Meindl-Beinker, A.N. TGF-β in Hepatic Stellate Cell Activation and Liver Fibrogenesis—Updated 2019. Cells 2019, 8, 1419. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Lee, Y.A.; Fujiwara, N.; Ybanez, M.; Allen, B.; Martins, S.; Fiel, M.I.; Goossens, N.; Chou, H.-I.; Hoshida, Y.; et al. A Simple Diet- And Chemical-Induced Murine NASH Model with Rapid Progression of Steatohepatitis, Fibrosis and Liver Cancer. J. Hepatol. 2018, 69, 385–395. [Google Scholar] [CrossRef]
- Heyens, L.J.M.; Busschots, D.; Koek, G.H.; Robaeys, G.; Francque, S. Liver Fibrosis in Non-alcoholic Fatty Liver Disease: From Liver Biopsy to Non-invasive Biomarkers in Diagnosis and Treatment. Front. Med. 2021, 8, 615978. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Sánchez, A.; Martínez-Ortega, A.J.; Remón-Ruiz, P.J.; Piñar-Gutiérrez, A.; Pereira-Cunill, J.L.; García-Luna, P.P. Therapeutic Properties and Use of Extra Virgin Olive Oil in Clinical Nutrition: A Narrative Review and Literature Update. Nutrients 2022, 14, 1440. [Google Scholar] [CrossRef]
- Kim, S.W.; Hur, W.; Li, T.Z.; Lee, Y.K.; Choi, J.E.; Hong, S.W.; Lyoo, K.-S.; You, C.R.; Jung, E.S.; Jung, C.K.; et al. Oleuropein Prevents the Progression of Steatohepatitis to Hepatic Fibrosis Induced by a High-Fat Diet in Mice. Exp. Mol. Med. 2014, 46, e92. [Google Scholar] [CrossRef]
- Gabbia, D.; Carpi, S.; Sarcognato, S.; Cannella, L.; Colognesi, M.; Scaffidi, M.; Polini, B.; Digiacomo, M.; Salsano, J.E.; Manera, C.; et al. The Extra Virgin Olive Oil Polyphenol Oleocanthal Exerts Antifibrotic Effects in the Liver. Front. Nutr. 2021, 8, 715183. [Google Scholar] [CrossRef]
- Harrison, S.A.; Torgerson, S.; Hayashi, P.; Ward, J.; Schenker, S. Vitamin E and Vitamin C Treatment Improves Fibrosis in Pa-Tients with Nonalcoholic Steatohepatitis. Am. J. Gastroenterol. 2003, 98, 2485–2490. [Google Scholar] [CrossRef]
- Chan, Y.T.; Wang, N.; Tan, H.Y.; Li, S.; Feng, Y. Targeting Hepatic Stellate Cells for the Treatment of Liver Fibrosis by Natural Products: Is It the Dawning of a New Era? Front. Pharmacol. 2020, 11, 548. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-Smooth Muscle Actin Expression Upregulates Fibroblast Contractile Activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Tovar, E.; Muriel, P. Molecular Mechanisms that Link Oxidative Stress, Inflammation, and Fibrosis in the Liver. Antioxidants 2020, 9, 1279. [Google Scholar] [CrossRef] [PubMed]
- Mak, A.L.; Lee, J.; van Dijk, A.M.; Vali, Y.; Aithal, G.P.; Schattenberg, J.M.; Anstee, Q.M.; Brosnan, M.J.; Zafarmand, M.H.; Ramsoekh, D.; et al. Systematic Review with Meta-Analysis: Di-agnostic Accuracy of Pro-C3 for Hepatic Fibrosis in Patients with Non-Alcoholic Fatty Liver Disease. Biomedicines 2021, 9, 1920. [Google Scholar] [CrossRef] [PubMed]
- Mosca, A.; Comparcola, D.; Romito, I.; Mantovani, A.; Nobili, V.; Byrne, C.D.; Alisi, A.; Targher, G. Plasma N-Terminal Pro-Peptide of Type III Procollagen Accurately Predicts Liver Fibrosis Severity in Children with Non-Alcoholic Fatty Liver Disease. Liver Int. 2019, 39, 2317–2329. [Google Scholar] [CrossRef]
- Loffredo, L.; Zicari, A.M.; Perri, L.; Carnevale, R.; Nocella, C.; Angelico, F.; Del Ben, M.; Mosca, A.; Zaffina, S.; Panera, N.; et al. Does Nox2 Overactivate in Children with Nonalcoholic Fatty Liver Disease? Antioxid Redox Signal 2019, 30, 1325–1330. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PLA Arm (n = 9) | HXT + VitE Arm (n = 16) | PLA vs. HXT + VitE (T1) | PLA vs. HXT + VitE (T2) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 | T1 | T2 | p | T0 | T1 | T2 | p | p | p | |
| Age (years) | 11.5 (2.6) | 12.1 (2.6) | 13.8 (2.6) | 0.45 | 12.3 (2.4) | 12.7 (2.4) | 14.3 (2.3) | 0.32 | 0.84 | 0.12 |
| BMI (Kg/m2) | 27 (6.7) | 26.9 (6.9) | 27.8 (3.6) | 0.78 | 26.9 (4.2) | 27.4 (4.9) | 26.2 (3) | 0.87 | 0.77 | 0.68 |
| WC (cm) | 92.3 (18.4) | 98.2 (19.6) | 94.2 (10.5) | 0.56 | 94.3 (10.8) | 98.1 (9.7) | 95.8 (7.4) | 0.94 | 0.92 | 0.84 |
| Total Cholesterol (mg/dL) | 172.5 (26.9) | 172.5 (37.8) | 161 (32.5) | 0.12 | 161.5 (30.8) | 149 (27.6) | 157.4 (39.7) | 0.51 | 0.10 | 0.42 |
| HDL-C (mg/dL) | 45.2 (5.9) | 42.2 (5.7) | 41.2 (6.4) | 0.83 | 47.3 (6.5) | 45.6 (7.2) | 49 (9.8) | 0.75 | 0.67 | 0.11 |
| LDL-C (mg/dL) | 117.5 (29.6) | 117 (38.2) | 106.3 (33.8) | 0.07 | 101.4 (30.8) | 95 (21) | 95.1 (35.8) | 0.27 | 0.04 | 0.034 |
| Triglycerides (mg/dL) | 112.3 (41.8) | 167.3 (42.4) | 117 (55) | 0.08 | 115.7 (56.5) | 87.7 (33.6) | 90.2 (42.6) | 0.03 | 0.04 | 0.03 |
| Uric Acid (mg/dL) | 5.5 (1.5) | 5.9 (1.6) | 6.5 (1.7) | 0.11 | 6.1 (1.2) | 5.9 (1.1) | 6.4 (1.5) | 0.40 | 0.96 | 0.95 |
| Fasting Glucose (mg/dL) | 87.2 (7.7) | 84.7 (3.2) | 89.7 (5.4) | 0.66 | 86.5 (5.8) | 86.7 (6.3) | 91 (6) | 0.71 | 0.67 | 0.57 |
| Fasting Insulin (mU/L) | 18.2 (2.8) | 14.8 (3.2) | 27.5 (15.4) | 0.05 | 24.3 (8.8) | 19.2 (10) | 25.4 (10.7) | 0.08 | 0.18 | 0.41 |
| HOMA- IR | 3.9 (0.7) | 3.1 (0.7) | 6.1 (3.5) | 0.03 | 5.2 (2) | 3.8 (2.3) | 5.6 (2.2) | 0.09 | 0.84 | 0.69 |
| AST (UI/L) | 29.4 (6.7) | 28.8 (7.1) | 28 (16.2) | 0.74 | 44.4 (45.4) | 36.7 (23.8) | 24.7 (10.9) | 0.04 | 0.11 | 0.22 |
| ALT (UI/L) | 40.3 (23.8) | 39.1 (19.1) | 35.2 (20.6) | 0.24 | 59.9 (56.4) | 45.2 (32.9) | 34.6 (24.5) | 0.05 | 0.57 | 0.89 |
| GGT (UI/L) | 20 (15.3) | 19.4 (11.7) | 26.4 (17) | 0.21 | 27.6 (28) | 23.1 (24.7) | 21.6 (11.2) | 0.04 | 0.61 | 0.46 |
| STEATOSIS | ||||||||||
| MILD | 0 | 1 (11.1) | 0 | -- | 0 | 3 (18.7) | 3 (18.7) | -- | 0.57 | -- |
| MODERATE | 3 (33.3) | 3 (33.3) | 5 (55.5) | 0.06 | 9 (56.2) | 11 (68.8) | 12 (75) | 0.07 | 0.02 | 0.12 |
| SEVERE | 6 (66.7) | 5 (55.6) | 4 (44.5) | 0.06 | 7 (43.8) | 2 (12.5) | 1 (6.3) | 0.001 | 0.03 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panera, N.; Braghini, M.R.; Crudele, A.; Smeriglio, A.; Bianchi, M.; Condorelli, A.G.; Nobili, R.; Conti, L.A.; De Stefanis, C.; Lioci, G.; et al. Combination Treatment with Hydroxytyrosol and Vitamin E Improves NAFLD-Related Fibrosis. Nutrients 2022, 14, 3791. https://doi.org/10.3390/nu14183791
Panera N, Braghini MR, Crudele A, Smeriglio A, Bianchi M, Condorelli AG, Nobili R, Conti LA, De Stefanis C, Lioci G, et al. Combination Treatment with Hydroxytyrosol and Vitamin E Improves NAFLD-Related Fibrosis. Nutrients. 2022; 14(18):3791. https://doi.org/10.3390/nu14183791
Chicago/Turabian StylePanera, Nadia, Maria Rita Braghini, Annalisa Crudele, Antonella Smeriglio, Marzia Bianchi, Angelo Giuseppe Condorelli, Rebecca Nobili, Libenzio Adrian Conti, Cristiano De Stefanis, Gessica Lioci, and et al. 2022. "Combination Treatment with Hydroxytyrosol and Vitamin E Improves NAFLD-Related Fibrosis" Nutrients 14, no. 18: 3791. https://doi.org/10.3390/nu14183791